- Будова ядра

Содержание
- 2. План: Будова нуклеїнових кислот. Будова клітинного ядра. Будова, типи і властивості метафазних хромосом.
- 3. Будова нуклеїнових кислот У залежності від природи вуглеводу, що входить до складу нуклеотидів, нуклеїнові кислоти розподіляють
- 4. Порівняння структури дволанцюгової ДНК та одноланцюгової РНК та азотистих основ
- 5. Азотисті основи тимін цитозин урацил аденін гуанін
- 6. Гетероциклічні основи, що входять до складу нуклеїнових кислот, є похідними пурину та піримідину. До основ групи
- 7. Основами групи піримідину , урацил (U), тимін (Т) і цитозин (С): урацил (U) 2,4- діоксипіримідин Тимін
- 8. До складу ДНК входять: аденін, гуанін, цитозин і тимін. До складу РНК аденін, гуанін, цитозин і
- 9. У нуклеїнових кислотах органічні основи сполучені N-глікозидним зв'язком з залишком D-рибози або 2-дезокси-D-рибози. Глікозидний зв'язок здійснюється
- 10. аденін дезоксирибоза дезоксирибонуклеозид N-глікозидний звязок
- 11. Назви нуклеозидів утворюють аналогічно назвам глікозидів. Так, нуклеозид, який складається з рибози та урацилу, називають β-урацилрибофуранозидом,
- 12. У назвах дезоксирибонуклеозидів додатково вводиться префікс дезокси- (дезоксі-), наприклад: дезоксіаденозин, дезоксигуанозин, дезоксицитидин. Винятком є назва нуклеозиду,
- 13. У нуклеїнових кислотах гідроксильна група біля С5' або С3' пентозного залишку нуклеозиду етерифікована ортофосфорною кислотою. Складний
- 14. Номенклатура нуклеотидів
- 15. Нуклеїнові кислоти являють собою продукти полімеризації моно-нуклеотидів. Нуклеотиди сполучаються в довгі ланцюги за допомогою фосфодіефірних зв'язків,
- 16. Рибонуклеїнові (РНК) та дезоксирибонуклеїнові (ДНК) кислоти Нуклеїнові кислоти являють собою високомолекулярні гетерополімери, які складаються з залишків
- 17. Певна послідовність нуклеотидних ланок у полінуклеотидному ланцюзі називається первинною структурою нуклеїнових кислот. (Просторова орієнтація полінуклеотидних ланцюгів
- 18. За моделлю Уотсона та Крика діаметр спіралі 1,8-2,0 нм. Кожний виток спіралі містить 10 пар основ.
- 19. Утворення нуклеотидів Нуклеотиди утворюються при взаємодії пентози з азотистою основою (утворюючи N-глікозидний зв'язок при взаємодії з
- 20. Структура Первина
- 22. Вторинна структура ДНК
- 24. Схема багатостадійного упакування молекули ДНК в хромосому
- 26. Еукаріотична клітина має три основні частини: 1) ядро; 2) цитоплазму; 3) оболонку
- 27. Ядро (лат. nucleus, грец. karion) забезпечує процеси біосинтезу й передачі спадкової інформації та складається із: ядерної
- 28. У 1825 р. Я. Пуркіньє вперше спостерігав клітинні ядра в яйцеклітині курки. У 1833 р. Р.
- 29. Функції ядра Збереження генетичної (спадкової) інформації серед клітинних поколінь; Підтримання постійної структури ДНК за допомогою так
- 30. Форма ядра: Найчастіше сферична; зустрічаються паличкоподібна, бобовидна, кільцеподібна, сегментована. Форма ядра залежить від: форми клітини (видовжені
- 31. В еукаріотичних клітинах основний генетичний матеріал зосереджений в ядрі. До складу сухої речовини ядра входить 80%
- 32. Остеокласт Адипоцити
- 33. Клітини крові (різна форма ядер) Тромбоцити Еритроцит Базофільний гранулоцит Лімфоцит Моноцит Нейтрофільний гранулоцит Еозинофільний гранулоцит Кісткова
- 34. Ядро локалізується завжди у певному місці клітини. Наприклад, у циліндричних клітинах шлунка, кишки воно займай базальне
- 35. Структура ядра: 1 – каріотека; 2 – ядерна пора; 3 – гетерохроматин; 4 –еухроматин; 5 –
- 36. Електронна мікрофотографія інтерфазного ядра
- 37. Ядерна оболонка (нуклеолема, каріолема, каріотека) Структура, що відокремлює ядро від цитоплазми, дає змогу здійснюватися обміну між
- 38. Зовнішня мембрана зовнішньої оболонки часто переходить у мембрани гранулярного ендоплазматичного ретикулума. Цитоплазматичний бік ядра часто вкритий
- 39. Функції ядерної оболонки бар'єрна функція: ядерна оболонка відокремлює вміст ядра, його генетичний матеріал від цитоплазми, обмежує
- 40. структура ядерної оболонки залежить від фази клітинного циклу на початку профази ядерна оболонка розчиняється в кінці
- 41. Структура ядерної пори: 1 – перинуклеарний простір; 2 – внутрішня мембрана; 3 – зовнішня мембрана; 4
- 42. Каріоплазма, або ядерний сік рідка частина ядра, желеподібний колоїд середовище в якому перебувають ядерця та хромосоми
- 43. Хроматин інтерфазна форма існування хромосом (структурний аналог хромосом) хромосоми видно лише під час поділу клітини це
- 44. Рівні організації хроматину Метафазна хромосома – 1400 нм Ділянка метафазної хромосоми – 700нм Нуклеомери - субодиниці
- 45. Будова хроматину
- 46. Нуклеосома – це часточка, що має вигляд нанизаних на нитку намистин. Щільно упаковані у вигляді спіралі
- 47. Види хроматину Гетерохроматин відповідає конденсованим під час інтерфази ділянкам хромосом; він є функціонально неактивним. Цей хроматин
- 48. Рівні спіралізації хромосом ДНК Нуклеосома Хроматинова фібрила Хроматида Конденсована хромосома
- 49. Взаємозв’язок між структурою хроматину та організацією хромосом
- 50. Хромосоми Хромосоми — це щільні паличко- або ниткоподібні тільця діаметром 0,2-2 мкм і довжиною у людини
- 51. Структурна організація метафазних хромосом Кожна хромосома складається з двох хроматид – конденсованих подвійних ланцюгів ДНК Сестринські
- 52. Структурна організація метафазних хромосом 1 - довге плече; 2 - коротке плече; 3 - центромера; 4-
- 54. Типи метафазних хромосом людини 1 – метацентрична; 2 – субметацентрична; 3 – акроцентрична з супутником; 4
- 55. Каріотип – диплоїдний набір хромосом клітини, який характеризується: кількістю хромосом, певними розмірами, формою, будовою Диплоїдний набір
- 56. ДНК в хромосомах ДНК в складі хромосом зв’язана з білками-гістонами Один комплекс із гістонів і ДНК
- 57. Каріотип людини
- 58. Ідіограма – розташування пар хромосом у порядку зменшення їх розмірів Гомологічні хромосоми – хромосоми однієї пари,
- 59. Хромосомний набір (каріотип) чоловіка
- 60. В каріотипі різностатевих організмів розрізняють статеві хромосоми (у людини це X-хромосома та Y-хромосома) і аутосоми (всі
- 61. ПОРУШЕННЯ СТРУКТУРИ ХРОМОСОМ Порушення структури хромосом відбувається в результаті спонтанних змін: Генні мутації (зміни на молекулярному
- 62. Хромосомна мутація: ДЕЛЕЦІЯ від лат. deletio — знищення — хромосомна аберація (перебудова), при якій відбувається втрата
- 63. Хромосомна мутація: ДУПЛІКАЦІЯ від лат. duplicatio — подвоєння — структурна хромосомна мутація, полягає в подвоєнні ділянки
- 64. Хромосомна мутація: ТРАНСЛОКАЦІЯ В процесі транслокації відбувається обмін ділянками негомологічних хромосом, але загальне число генів не
- 65. Хромосомна мутація: ІНВЕРСІЯ Зміна структури хромосоми, яка викликана поворотом на 180° однієї з внутрішніх її ділянок.
- 66. ЯДЕРЦЕ Ядерце — це найбільш щільна структура ядра (щільність ядерця в 1,5 раза перевищує щільність ядра),
- 67. Ядерця щільні, сильно забарвлені гранулярні утворення, які не мають мембрани відбувається синтез рибосомальної РНК утворення субодиниць
- 68. Електронна мікрофотографія ядерця
- 70. Скачать презентацию

План:
Будова нуклеїнових кислот.
Будова клітинного ядра.
Будова, типи і властивості метафазних хромосом.
План:
Будова нуклеїнових кислот.
Будова клітинного ядра.
Будова, типи і властивості метафазних хромосом.

Будова нуклеїнових кислот
У залежності від природи вуглеводу, що входить до складу
Будова нуклеїнових кислот
У залежності від природи вуглеводу, що входить до складу

Порівняння структури дволанцюгової ДНК
та одноланцюгової РНК та азотистих основ
Порівняння структури дволанцюгової ДНК
та одноланцюгової РНК та азотистих основ
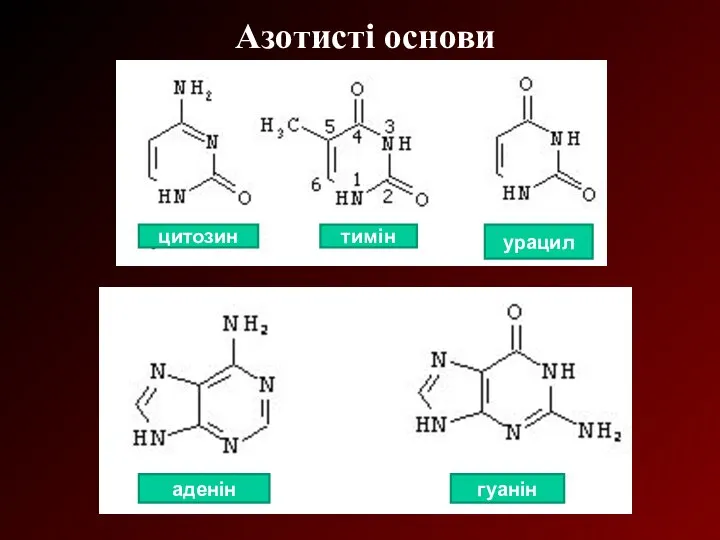
Азотисті основи
тимін
цитозин
урацил
аденін
гуанін
Азотисті основи
тимін
цитозин
урацил
аденін
гуанін

Гетероциклічні основи, що входять до складу нуклеїнових кислот, є похідними пурину
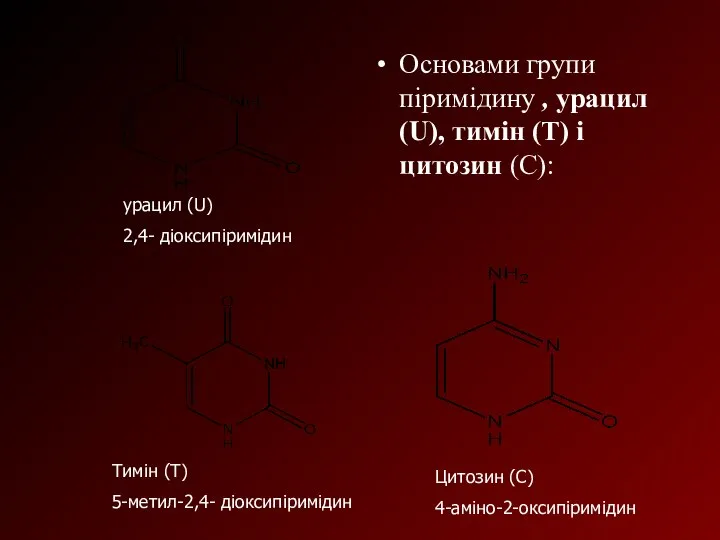
Основами групи піримідину , урацил (U), тимін (Т) і цитозин (С):
Основами групи піримідину , урацил (U), тимін (Т) і цитозин (С):
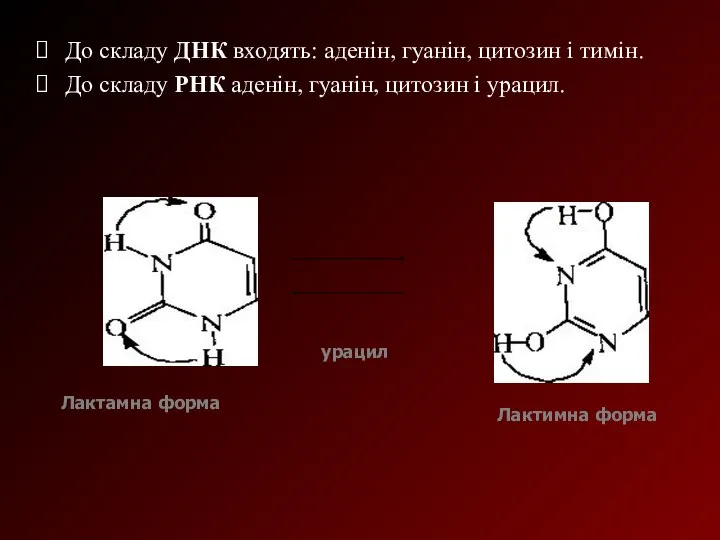
До складу ДНК входять: аденін, гуанін, цитозин і тимін.
До складу
До складу ДНК входять: аденін, гуанін, цитозин і тимін.
До складу

У нуклеїнових кислотах органічні основи сполучені N-глікозидним зв'язком з залишком D-рибози
У нуклеїнових кислотах органічні основи сполучені N-глікозидним зв'язком з залишком D-рибози
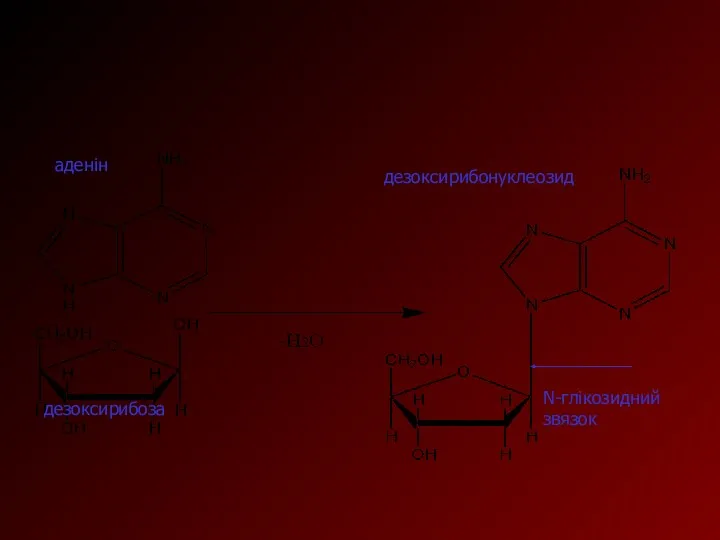
аденін
дезоксирибоза
дезоксирибонуклеозид
N-глікозидний звязок
аденін
дезоксирибоза
дезоксирибонуклеозид
N-глікозидний звязок
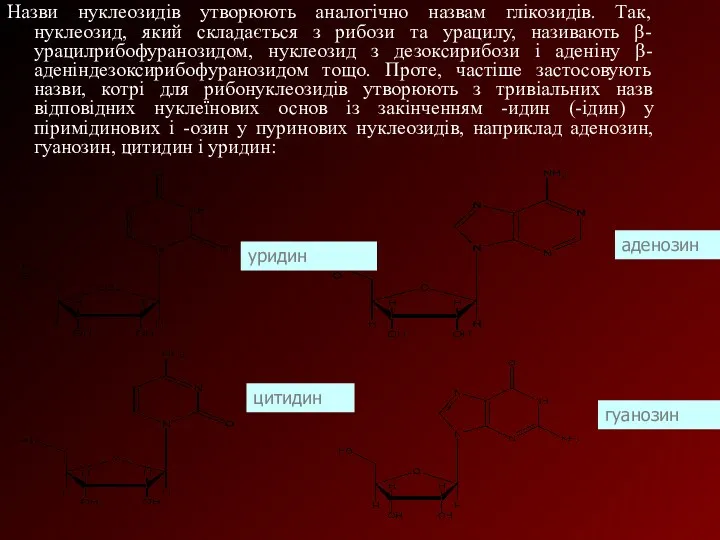
Назви нуклеозидів утворюють аналогічно назвам глікозидів. Так, нуклеозид, який складається з
Назви нуклеозидів утворюють аналогічно назвам глікозидів. Так, нуклеозид, який складається з

У назвах дезоксирибонуклеозидів додатково вводиться префікс дезокси- (дезоксі-), наприклад: дезоксіаденозин, дезоксигуанозин,
У назвах дезоксирибонуклеозидів додатково вводиться префікс дезокси- (дезоксі-), наприклад: дезоксіаденозин, дезоксигуанозин,

У нуклеїнових кислотах гідроксильна група біля С5' або С3' пентозного залишку
У нуклеїнових кислотах гідроксильна група біля С5' або С3' пентозного залишку

Номенклатура нуклеотидів
Номенклатура нуклеотидів

Нуклеїнові кислоти являють собою продукти полімеризації моно-нуклеотидів. Нуклеотиди сполучаються в довгі
Нуклеїнові кислоти являють собою продукти полімеризації моно-нуклеотидів. Нуклеотиди сполучаються в довгі
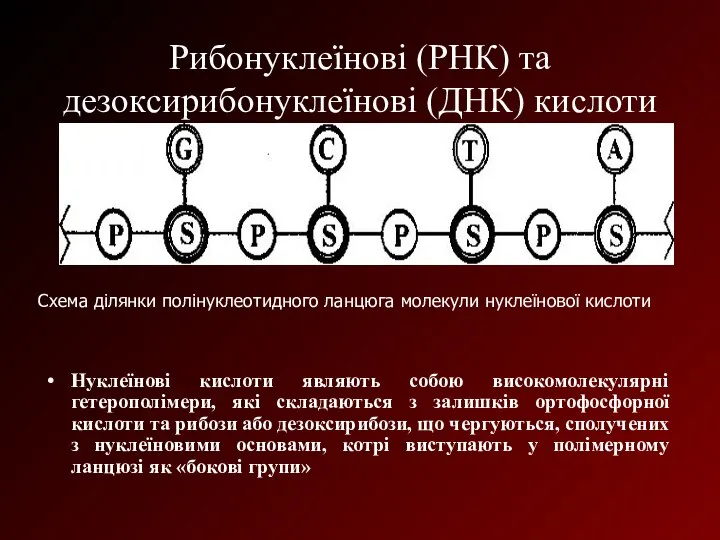
Рибонуклеїнові (РНК) та дезоксирибонуклеїнові (ДНК) кислоти
Нуклеїнові кислоти являють собою високомолекулярні гетерополімери,
Рибонуклеїнові (РНК) та дезоксирибонуклеїнові (ДНК) кислоти
Нуклеїнові кислоти являють собою високомолекулярні гетерополімери,

Певна послідовність нуклеотидних ланок у полінуклеотидному ланцюзі називається первинною структурою нуклеїнових
Певна послідовність нуклеотидних ланок у полінуклеотидному ланцюзі називається первинною структурою нуклеїнових

За моделлю Уотсона та Крика діаметр спіралі 1,8-2,0 нм. Кожний виток
За моделлю Уотсона та Крика діаметр спіралі 1,8-2,0 нм. Кожний виток

Утворення нуклеотидів
Нуклеотиди утворюються при взаємодії пентози з азотистою основою (утворюючи
Утворення нуклеотидів
Нуклеотиди утворюються при взаємодії пентози з азотистою основою (утворюючи
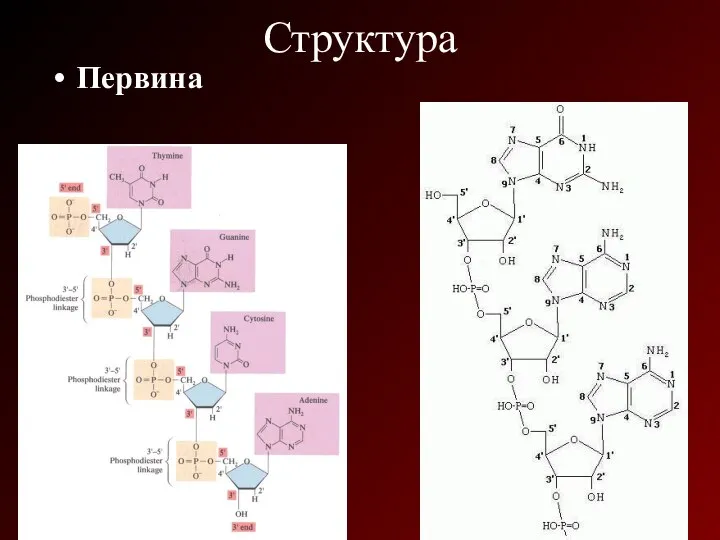
Структура
Первина
Структура
Первина


Вторинна структура ДНК
Вторинна структура ДНК

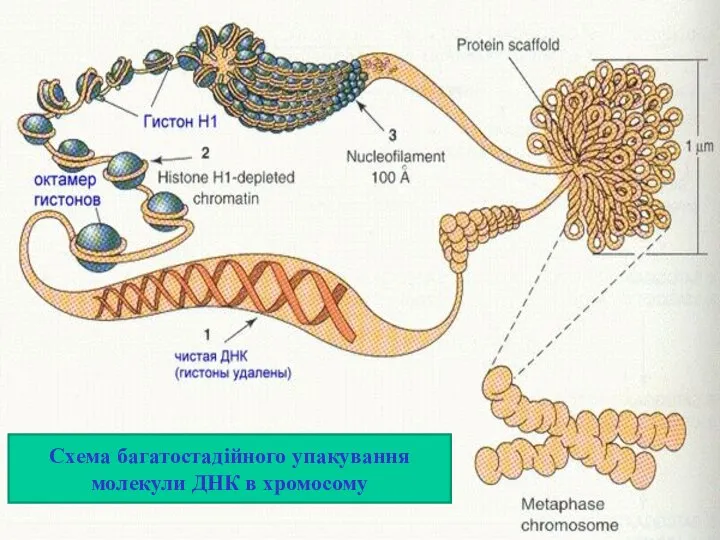
Схема багатостадійного упакування молекули ДНК в хромосому
Схема багатостадійного упакування молекули ДНК в хромосому


Еукаріотична клітина має три основні частини: 1) ядро; 2) цитоплазму; 3)
Еукаріотична клітина має три основні частини: 1) ядро; 2) цитоплазму; 3)
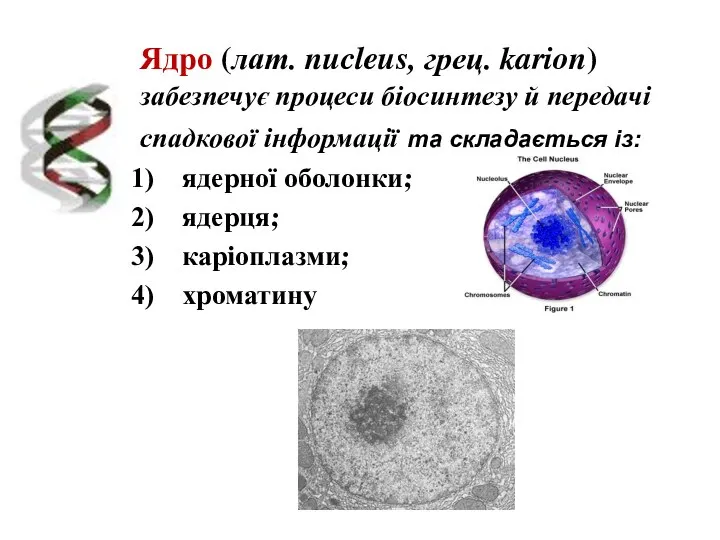
Ядро (лат. nucleus, грец. karion) забезпечує процеси біосинтезу й передачі спадкової
Ядро (лат. nucleus, грец. karion) забезпечує процеси біосинтезу й передачі спадкової

У 1825 р. Я. Пуркіньє вперше спостерігав клітинні ядра в яйцеклітині
У 1825 р. Я. Пуркіньє вперше спостерігав клітинні ядра в яйцеклітині

Функції ядра
Збереження генетичної (спадкової) інформації серед клітинних поколінь;
Підтримання постійної структури ДНК
Функції ядра
Збереження генетичної (спадкової) інформації серед клітинних поколінь;
Підтримання постійної структури ДНК

Форма ядра:
Найчастіше сферична;
зустрічаються паличкоподібна, бобовидна, кільцеподібна, сегментована.
Форма ядра залежить від:
форми клітини (видовжені
Форма ядра:
Найчастіше сферична;
зустрічаються паличкоподібна, бобовидна, кільцеподібна, сегментована.
Форма ядра залежить від:
форми клітини (видовжені

В еукаріотичних клітинах основний генетичний матеріал зосереджений в ядрі.
До складу
В еукаріотичних клітинах основний генетичний матеріал зосереджений в ядрі.
До складу

Остеокласт
Адипоцити
Остеокласт
Адипоцити
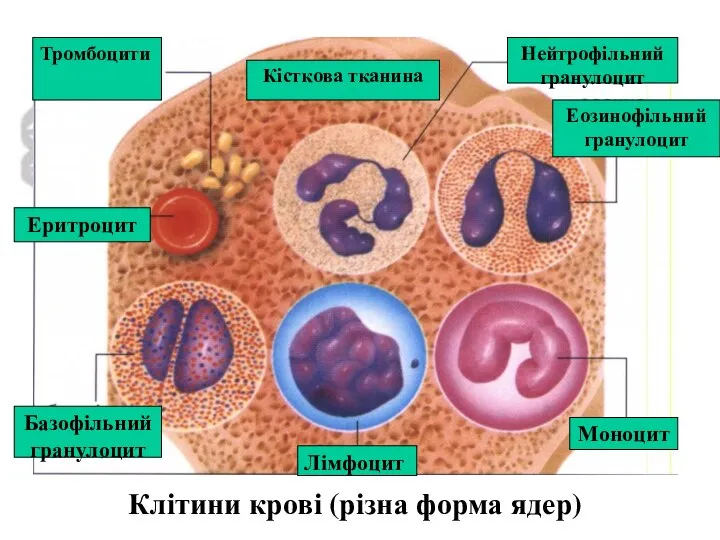
Клітини крові (різна форма ядер)
Тромбоцити
Еритроцит
Базофільний гранулоцит
Лімфоцит
Моноцит
Нейтрофільний гранулоцит
Еозинофільний гранулоцит
Кісткова тканина
Клітини крові (різна форма ядер)
Тромбоцити
Еритроцит
Базофільний гранулоцит
Лімфоцит
Моноцит
Нейтрофільний гранулоцит
Еозинофільний гранулоцит
Кісткова тканина

Ядро локалізується завжди у певному місці клітини. Наприклад, у циліндричних клітинах шлунка,
Ядро локалізується завжди у певному місці клітини. Наприклад, у циліндричних клітинах шлунка,

Структура ядра:
1 – каріотека;
2 – ядерна пора;
3 – гетерохроматин;
Структура ядра:
1 – каріотека;
2 – ядерна пора;
3 – гетерохроматин;

Електронна мікрофотографія інтерфазного ядра
Електронна мікрофотографія інтерфазного ядра

Ядерна оболонка
(нуклеолема, каріолема, каріотека)
Структура, що відокремлює ядро від цитоплазми,
Ядерна оболонка
(нуклеолема, каріолема, каріотека)
Структура, що відокремлює ядро від цитоплазми,

Зовнішня мембрана зовнішньої оболонки часто переходить у мембрани гранулярного ендоплазматичного ретикулума.
Цитоплазматичний
Зовнішня мембрана зовнішньої оболонки часто переходить у мембрани гранулярного ендоплазматичного ретикулума.
Цитоплазматичний

Функції ядерної оболонки
бар'єрна функція: ядерна оболонка відокремлює вміст ядра, його генетичний матеріал
Функції ядерної оболонки
бар'єрна функція: ядерна оболонка відокремлює вміст ядра, його генетичний матеріал

структура ядерної оболонки залежить від фази клітинного циклу
на початку профази ядерна
структура ядерної оболонки залежить від фази клітинного циклу
на початку профази ядерна

Структура ядерної пори:
1 – перинуклеарний
простір;
2 – внутрішня мембрана;
3 –
Структура ядерної пори:
1 – перинуклеарний
простір;
2 – внутрішня мембрана;
3 –

Каріоплазма, або ядерний сік
рідка частина ядра, желеподібний колоїд
середовище в якому перебувають
Каріоплазма, або ядерний сік
рідка частина ядра, желеподібний колоїд
середовище в якому перебувають

Хроматин
інтерфазна форма існування хромосом (структурний аналог хромосом)
хромосоми видно лише під
Хроматин
інтерфазна форма існування хромосом (структурний аналог хромосом)
хромосоми видно лише під

Рівні організації хроматину
Метафазна хромосома – 1400 нм
Ділянка метафазної хромосоми – 700нм
Нуклеомери
Рівні організації хроматину
Метафазна хромосома – 1400 нм
Ділянка метафазної хромосоми – 700нм
Нуклеомери
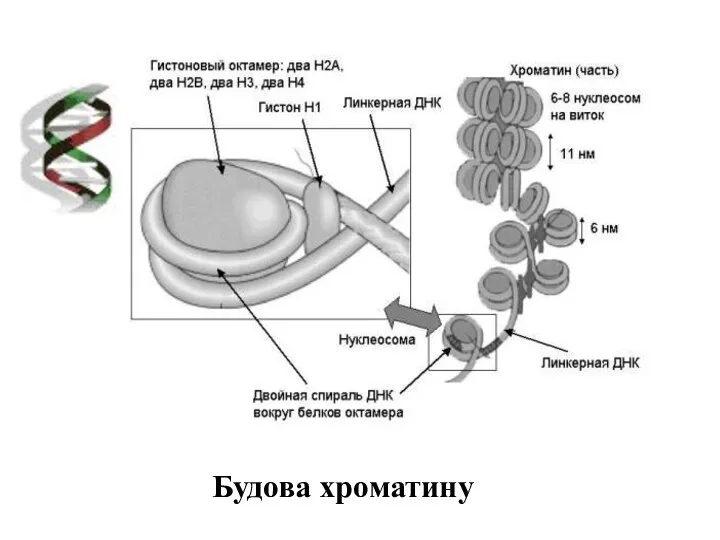
Будова хроматину
Будова хроматину

Нуклеосома – це часточка, що має вигляд нанизаних на нитку намистин.
Нуклеосома – це часточка, що має вигляд нанизаних на нитку намистин.

Види хроматину
Гетерохроматин відповідає конденсованим під час інтерфази ділянкам хромосом; він є
Види хроматину
Гетерохроматин відповідає конденсованим під час інтерфази ділянкам хромосом; він є

Рівні спіралізації хромосом
ДНК
Нуклеосома
Хроматинова фібрила
Хроматида
Конденсована хромосома
Рівні спіралізації хромосом
ДНК
Нуклеосома
Хроматинова фібрила
Хроматида
Конденсована хромосома

Взаємозв’язок між структурою хроматину та
організацією хромосом
Взаємозв’язок між структурою хроматину та
організацією хромосом

Хромосоми
Хромосоми — це щільні паличко- або ниткоподібні тільця діаметром 0,2-2 мкм і довжиною
Хромосоми
Хромосоми — це щільні паличко- або ниткоподібні тільця діаметром 0,2-2 мкм і довжиною

Структурна організація
метафазних хромосом
Кожна хромосома складається з двох хроматид – конденсованих
Структурна організація
метафазних хромосом
Кожна хромосома складається з двох хроматид – конденсованих

Структурна організація
метафазних хромосом
1 - довге плече;
2 - коротке
Структурна організація
метафазних хромосом
1 - довге плече;
2 - коротке


Типи метафазних хромосом людини
1 – метацентрична;
2 – субметацентрична;
3 – акроцентрична
Типи метафазних хромосом людини
1 – метацентрична;
2 – субметацентрична;
3 – акроцентрична

Каріотип – диплоїдний набір хромосом клітини, який характеризується: кількістю хромосом, певними
Каріотип – диплоїдний набір хромосом клітини, який характеризується: кількістю хромосом, певними

ДНК в хромосомах
ДНК в складі хромосом зв’язана з білками-гістонами
Один комплекс із
ДНК в хромосомах
ДНК в складі хромосом зв’язана з білками-гістонами
Один комплекс із

Каріотип людини
Каріотип людини

Ідіограма – розташування пар хромосом у порядку зменшення їх розмірів
Гомологічні хромосоми
Ідіограма – розташування пар хромосом у порядку зменшення їх розмірів
Гомологічні хромосоми

Хромосомний набір (каріотип) чоловіка
Хромосомний набір (каріотип) чоловіка

В каріотипі різностатевих організмів розрізняють статеві хромосоми (у людини це X-хромосома
В каріотипі різностатевих організмів розрізняють статеві хромосоми (у людини це X-хромосома

ПОРУШЕННЯ СТРУКТУРИ ХРОМОСОМ
Порушення структури хромосом відбувається в результаті спонтанних змін:
Генні мутації
ПОРУШЕННЯ СТРУКТУРИ ХРОМОСОМ
Порушення структури хромосом відбувається в результаті спонтанних змін:
Генні мутації

Хромосомна мутація: ДЕЛЕЦІЯ
від лат. deletio — знищення — хромосомна аберація (перебудова),
Хромосомна мутація: ДЕЛЕЦІЯ
від лат. deletio — знищення — хромосомна аберація (перебудова),

Хромосомна мутація: ДУПЛІКАЦІЯ
від лат. duplicatio — подвоєння — структурна хромосомна
Хромосомна мутація: ДУПЛІКАЦІЯ
від лат. duplicatio — подвоєння — структурна хромосомна

Хромосомна мутація: ТРАНСЛОКАЦІЯ
В процесі транслокації відбувається обмін ділянками негомологічних хромосом, але
Хромосомна мутація: ТРАНСЛОКАЦІЯ
В процесі транслокації відбувається обмін ділянками негомологічних хромосом, але

Хромосомна мутація: ІНВЕРСІЯ
Зміна структури хромосоми, яка викликана поворотом на 180° однієї
Хромосомна мутація: ІНВЕРСІЯ
Зміна структури хромосоми, яка викликана поворотом на 180° однієї

ЯДЕРЦЕ
Ядерце — це найбільш щільна структура ядра (щільність ядерця в 1,5
ЯДЕРЦЕ
Ядерце — це найбільш щільна структура ядра (щільність ядерця в 1,5

Ядерця
щільні, сильно забарвлені гранулярні утворення, які не мають мембрани
відбувається синтез рибосомальної
Ядерця
щільні, сильно забарвлені гранулярні утворення, які не мають мембрани
відбувається синтез рибосомальної

Електронна мікрофотографія ядерця
Електронна мікрофотографія ядерця
 Макроэволюция
Макроэволюция Строение костной системы организма и ее функции. Лекция №3
Строение костной системы организма и ее функции. Лекция №3 Опорно-двигательная система человека
Опорно-двигательная система человека Тесты по биологии
Тесты по биологии Рост и развитие животных
Рост и развитие животных Микробиологическая оценка обсемененности продуктов переработки яблок
Микробиологическая оценка обсемененности продуктов переработки яблок Хвойные растения
Хвойные растения  Строение и работа сердца
Строение и работа сердца Всем привет Поговорим о теориях происхождения жизни.
Всем привет Поговорим о теориях происхождения жизни. Теплорегуляция Интегрированный урок (физики и биологии)
Теплорегуляция Интегрированный урок (физики и биологии) Гены развития дрозофилы
Гены развития дрозофилы Презентация Полевые сорняки
Презентация Полевые сорняки Влияние породы коров на физико-химичесский состав молока и качество сливочного масла. Оценка племенных животных. Мечение скота
Влияние породы коров на физико-химичесский состав молока и качество сливочного масла. Оценка племенных животных. Мечение скота Природа вокруг нас
Природа вокруг нас Гены в хромосомах и популяциях
Гены в хромосомах и популяциях Презентация на тему «Лекарственные и ядовитые растения» Презентацию подготовила ученица 6 А класса МОУ-СОШ № 33 Резнеченко Ангел
Презентация на тему «Лекарственные и ядовитые растения» Презентацию подготовила ученица 6 А класса МОУ-СОШ № 33 Резнеченко Ангел Споры и Спорангии
Споры и Спорангии Презентация на тему "Семейство Крестоцветные" - скачать презентации по Биологии
Презентация на тему "Семейство Крестоцветные" - скачать презентации по Биологии Пищеварительный канал
Пищеварительный канал Селекция микроорганизмов
Селекция микроорганизмов Международный день птиц
Международный день птиц Рани. Їх класифікація. Ярової М. Авак’ян О.
Рани. Їх класифікація. Ярової М. Авак’ян О.  Генетика пола. Наследование, сцепленное с полом
Генетика пола. Наследование, сцепленное с полом История развития, основные достижения и проблемы медицинской генетики. Цитологические основы наследственности
История развития, основные достижения и проблемы медицинской генетики. Цитологические основы наследственности Презентация на тему Ежи морские (Echinoidea)
Презентация на тему Ежи морские (Echinoidea)  Мутагенные факторы
Мутагенные факторы Взаимоотношения между организмами хищничество
Взаимоотношения между организмами хищничество Выполнил: учитель биологии МБОУ ООШ п.Туголесский Бор Солдакова Марина Александровна
Выполнил: учитель биологии МБОУ ООШ п.Туголесский Бор Солдакова Марина Александровна