- Генетика бактерий. Экспрессия генетической информации у бактерий

Содержание
- 2. Геном бактерий Геном – совокупность всех генов бактерий; Бактерии – гаплоидные организмы, имеют один набор генов;
- 3. Организация генетического аппарата бактерий Нуклеоид - одна двунитевая молекула ДНК кольцевой формы; Содержание ДНК непостоянно, может
- 4. Нуклеоид содержит информацию необходимую для обеспечения основных процессов жизнедеятельности - кодирует синтез ферментов, участвующих в пластическом
- 5. Организация нуклеоида кольцевая молекула ДНК; длина 1,6 мм, диаметр 1 мкм; суперскрученное состояние обеспечивается топоизомеразой; петли
- 6. Общая характеристика генома бактерий структурные гены – кодирующие области – 85% последовательностей ДНК бактерий; регуляторные области;
- 7. Оперонная организация бактериальных генов Оперон – группа структурных генов (кодируют признаки), находящихся под общим контролем; В
- 8. Внехромосомные генетические элементы - плазмиды, транспозоны, вставочные последовательности Различаются по молекулярной массе, объему закодированной информации, способности
- 9. Плазмиды автономно реплицирующиеся двухцепочечные молекулы ДНК
- 10. Классификация плазмид по свойствам F – плазмиды; R – плазмиды; Col – плазмиды; Ent – плазмиды;
- 11. Классификация плазмид по способу межклеточной передачи Конъюгативные (трансмиссивные) – осуществляют собственный перенос путем конъюгации. Содержат tra-опероны
- 12. Классификация плазмид по совместимости Несовместимость – родственные плазмиды, обладающие высоким сходством репликонов неспособны существовать в одной
- 13. Значение плазмид обусловливают гетерогенность микробных популяций; контролируют обмен генетическим материалом; контролируют синтез факторов (в том числе
- 14. Транспозоны (Tn)- мобильные генетические элементы Фрагменты ДНК, состоящие из генов, кодирующих транспозицию (перемещение) и признаки; Способны
- 15. Вставочные последовательности (IS-элементы) Фрагменты ДНК, несущие только гены, кодирующие собственное перемещение (транспозицию) - фермент транспозазу и
- 16. Функции IS-элементов Координируют взаимодействие мобильных генетических элементов между собой и бактериальной хромосомой; Регулируют экспрессию структурных генов;
- 17. «Островки» патогенности –фрагменты ДНК, кодирующие факторы болезнетворности Обнаружены в геноме болезнетворных бактерий; Располагаются отдельными кластерами в
- 18. Структура «островков» патогенности Содержат мобильные элементы (IS), гены подвижности (интегразу, транспозазу), гены вирулентности (V1-V4); По обоим
- 19. Передача генетической информации у бактерий по вертикали (по наследству) – обеспечивает передачу всех генов исходной особи
- 20. Передача генетической информации у бактерий по горизонтали – способствует возникновению новых признаков – изменчивости; основной механизм
- 21. Конъюгация - перенос генетического материала посредством конъюгативных пилей хеликаза осуществляет разрывы водородных связей в двухцепочечной ДНК;
- 22. Трансмиссивная F-плазмида находится в клетке в автономном состоянии; при участии хеликазы и эндонуклеазы образуется однонитевая ДНК;
- 23. Интегративная F-плазмида F-плазмида и хромосома бактериальной клетки вместе образуют единый трансмиссивный репликон; клетки со встроенной F-плазмидой
- 24. Трансформация поглощение фрагментов ДНК и включение в хромосому бактерии реципиента; ДНК должна быть двунитевой; длина фрагмента
- 25. Механизм трансформации Аутолизин разрушает КС, обнажает ДНК-связывающий белок и эндонуклеазу I; ДНК-связывающий белок абсорбирует фрагменты донорной
- 26. Трансдукция передача генетического материала от одной бактерии другой посредством бактериофага общая (неспецифическая) - перенос бактериофагом любого
- 27. Экспрессия генетической информации у бактерий
- 28. Бактерии – универсальные существа Отдельная бактериальная клетка – полноценный самодостаточный организм; Клетки многоклеточных входят в состав
- 29. Парадокс бактериальной клетки Высокая метаболическая активность (правило Рубнера); Высокая ферментативная насыщенность (до 109-1012 реакций в минуту);
- 30. Быстрая адаптация к изменяющимся условиям среды - необходимое условие выживания Бактерии должны иметь все ферменты, необходимые
- 31. Бактерии нуждаются в получении информации о своем окружении Способы получения информации бактериями: непосредственный контакт при конъюгации
- 32. Внешние сигналы определяют экспрессию генов Сигналы распознаются и преобразуются; Передаются генетическим структурам; Эта информация реализуется на
- 33. Регуляторы экспрессии генетической информации двухкомпонентные сигнальные системы. Широко распространенны среди прокариотических организмов. Сенсорная киназа– ключевой компонент
- 34. Уровни регуляции метаболизма На уровне транскрипции (путь от ДНК к РНК); Регуляция на начальных этапах экспрессии
- 35. Регуляторы экспрессии оперонов Регуляторные белки Продукты генов-регуляторов Связываются с участком ДНК -оператором; Осуществляют позитивный или негативный
- 36. Эффекты связывания регуляторного белка с оператором Негативный контроль регуляторный белок препятствует транскрипции Фермент не синтезируется Позитивный
- 37. Индуцибельные опероны Катаболитные опероны; Цель регуляции – включить синтез ферментов, которые ранее не требовались; Оперон «включается»
- 38. Катаболитная репрессия Если в среде присутствует несколько субстратов (глюкоза и лактоза), то сначала утилизируется субстрат, поддерживающий
- 39. Механизм катаболитной репрессии Эффектором выступает цАМФ; Белок активатор катаболитных оперонов (БАК) неактивен в свободном состоянии; Комплекс
- 40. Циклический АМФ регулятор активности БАК Образуется из АТФ с помощью аденилатциклазы; При недостатке глюкозы компоненты ее
- 41. Арабинозный оперон Гены araА, araВ, araD структурные – кодируют синтез ферментов, образуют оперон araВАD ; Ген
- 42. Структура арабинозного оперона В присутствии арабинозы AraC превращается в активатор, присоединяется к инициатору (I); БАК в
- 43. Работа арабинозного оперона в присутствии арабинозы Репрессор арабиноза Актива тор БАК цАМФ Белок активатор катаболитных оперонов
- 44. Работа арабинозного оперона в отсутствии арабинозы Репрессор
- 45. Репрессибельные опероны Характерны для анаболических путей. Цель регуляции – прекращение синтеза; Оперон «выключается» в присутствии молекул
- 46. Триптофановый оперон При наличии в среде триптофана, его синтез прекращается (репрессия конечным продуктом); Эффектор (корепрессор) –
- 47. Принцип работы триптофанового оперона
- 48. Аттенуация – механизм тонкой регуляции экспрессии структурных генов Позволяет регулировать количество синтезирующегося триптофана; При избытке триптофана
- 49. Аттенуатор - последовательность нуклеотидов в регуляторной области перед первым структурным геном Р - промотор; О -
- 51. Скачать презентацию

Геном бактерий
Геном – совокупность всех генов бактерий;
Бактерии – гаплоидные организмы, имеют
Геном бактерий
Геном – совокупность всех генов бактерий;
Бактерии – гаплоидные организмы, имеют

Организация генетического аппарата бактерий
Нуклеоид - одна двунитевая молекула ДНК кольцевой формы;
Содержание
Организация генетического аппарата бактерий
Нуклеоид - одна двунитевая молекула ДНК кольцевой формы;
Содержание
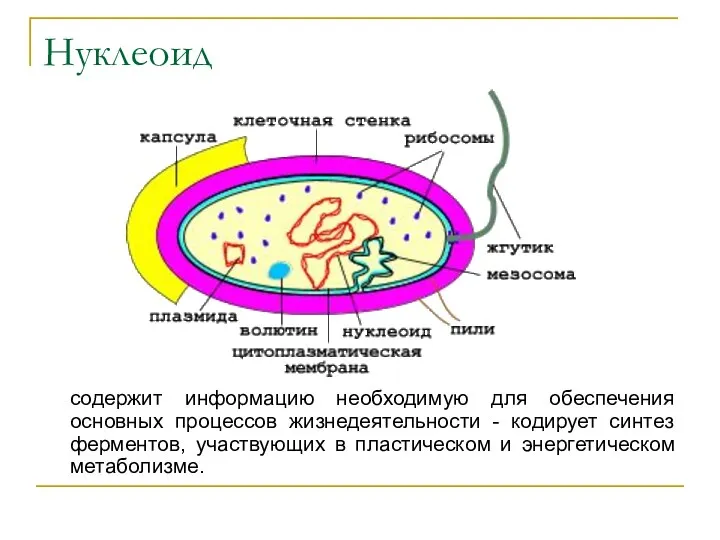
Нуклеоид
содержит информацию необходимую для обеспечения основных процессов жизнедеятельности - кодирует синтез
Нуклеоид
содержит информацию необходимую для обеспечения основных процессов жизнедеятельности - кодирует синтез
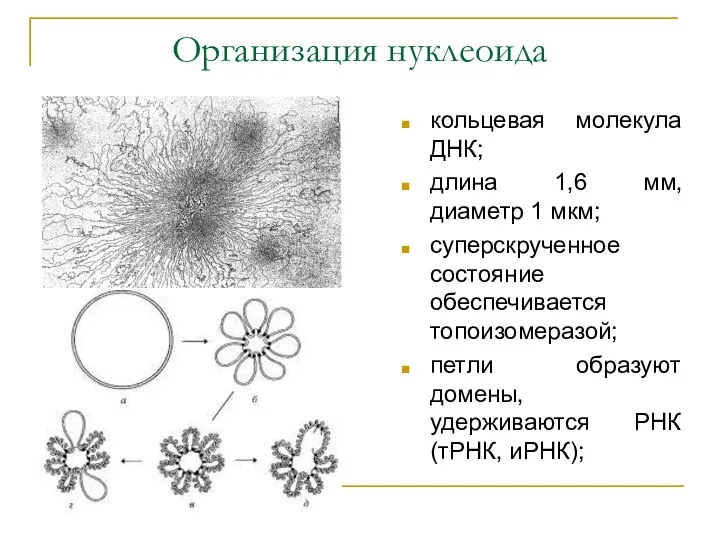
Организация нуклеоида
кольцевая молекула ДНК;
длина 1,6 мм, диаметр 1 мкм;
суперскрученное состояние обеспечивается
Организация нуклеоида
кольцевая молекула ДНК;
длина 1,6 мм, диаметр 1 мкм;
суперскрученное состояние обеспечивается

Общая характеристика генома бактерий
структурные гены – кодирующие области – 85% последовательностей
Общая характеристика генома бактерий
структурные гены – кодирующие области – 85% последовательностей

Оперонная организация бактериальных генов
Оперон – группа структурных генов (кодируют признаки), находящихся
Оперонная организация бактериальных генов
Оперон – группа структурных генов (кодируют признаки), находящихся

Внехромосомные генетические элементы - плазмиды, транспозоны, вставочные последовательности
Различаются по молекулярной массе,
Внехромосомные генетические элементы - плазмиды, транспозоны, вставочные последовательности
Различаются по молекулярной массе,
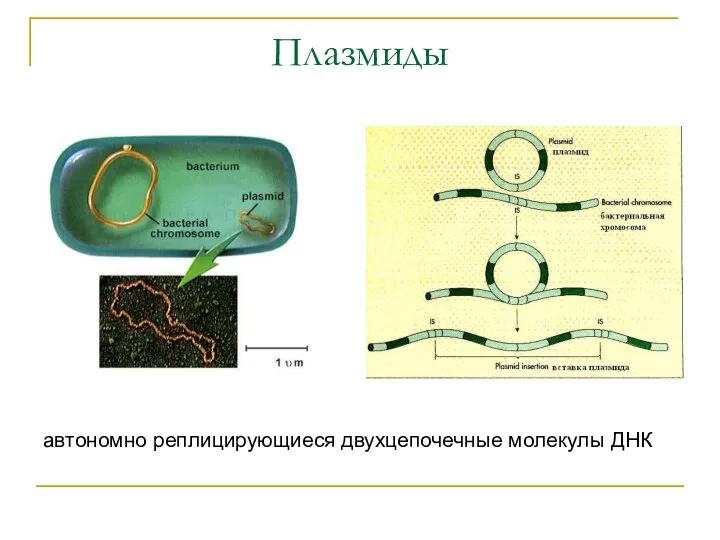
Плазмиды
автономно реплицирующиеся двухцепочечные молекулы ДНК
Плазмиды
автономно реплицирующиеся двухцепочечные молекулы ДНК

Классификация плазмид по свойствам
F – плазмиды;
R – плазмиды;
Col – плазмиды;
Ent –
Классификация плазмид по свойствам
F – плазмиды;
R – плазмиды;
Col – плазмиды;
Ent –

Классификация плазмид по способу межклеточной передачи
Конъюгативные (трансмиссивные) – осуществляют собственный перенос
Классификация плазмид по способу межклеточной передачи
Конъюгативные (трансмиссивные) – осуществляют собственный перенос

Классификация плазмид по совместимости
Несовместимость – родственные плазмиды, обладающие высоким сходством репликонов
Классификация плазмид по совместимости
Несовместимость – родственные плазмиды, обладающие высоким сходством репликонов

Значение плазмид
обусловливают гетерогенность микробных популяций;
контролируют обмен генетическим материалом;
контролируют синтез факторов (в
Значение плазмид
обусловливают гетерогенность микробных популяций;
контролируют обмен генетическим материалом;
контролируют синтез факторов (в
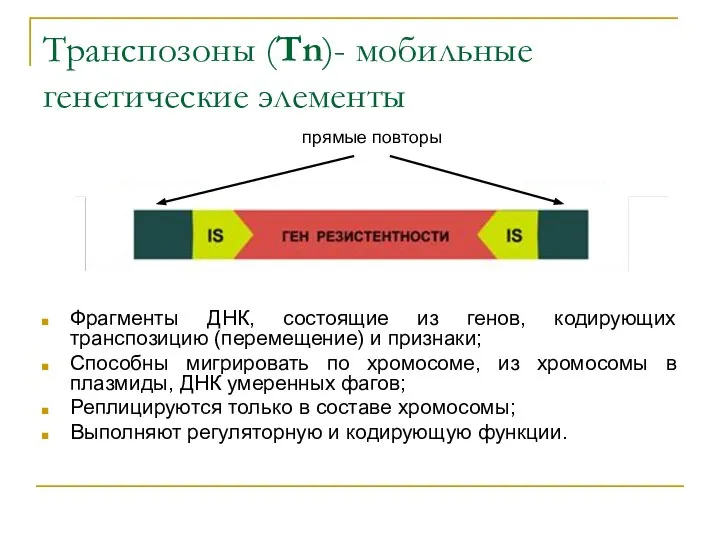
Транспозоны (Tn)- мобильные генетические элементы
Фрагменты ДНК, состоящие из генов, кодирующих транспозицию
Транспозоны (Tn)- мобильные генетические элементы
Фрагменты ДНК, состоящие из генов, кодирующих транспозицию
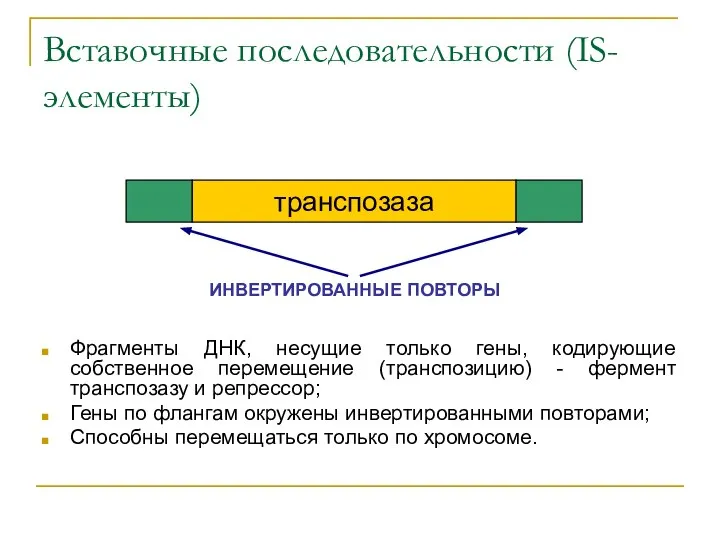
Вставочные последовательности (IS-элементы)
Фрагменты ДНК, несущие только гены, кодирующие собственное перемещение (транспозицию)
Вставочные последовательности (IS-элементы)
Фрагменты ДНК, несущие только гены, кодирующие собственное перемещение (транспозицию)

Функции IS-элементов
Координируют взаимодействие мобильных генетических элементов между собой и бактериальной
Функции IS-элементов
Координируют взаимодействие мобильных генетических элементов между собой и бактериальной

«Островки» патогенности –фрагменты ДНК, кодирующие факторы болезнетворности
Обнаружены в геноме болезнетворных бактерий;
Располагаются
«Островки» патогенности –фрагменты ДНК, кодирующие факторы болезнетворности
Обнаружены в геноме болезнетворных бактерий;
Располагаются
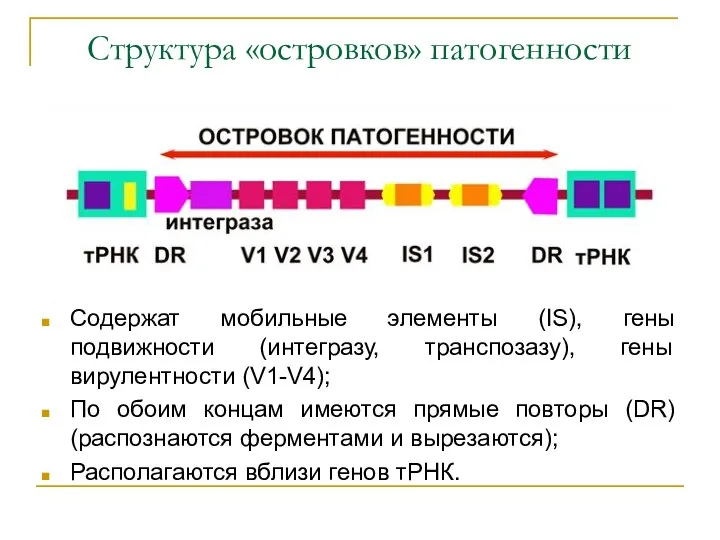
Структура «островков» патогенности
Содержат мобильные элементы (IS), гены подвижности (интегразу, транспозазу), гены
Структура «островков» патогенности
Содержат мобильные элементы (IS), гены подвижности (интегразу, транспозазу), гены
Передача генетической информации у бактерий
по вертикали (по наследству) – обеспечивает передачу
Передача генетической информации у бактерий
по вертикали (по наследству) – обеспечивает передачу

Передача генетической информации у бактерий
по горизонтали – способствует возникновению новых
Передача генетической информации у бактерий
по горизонтали – способствует возникновению новых

Конъюгация - перенос генетического материала посредством конъюгативных пилей
хеликаза осуществляет разрывы водородных
Конъюгация - перенос генетического материала посредством конъюгативных пилей
хеликаза осуществляет разрывы водородных
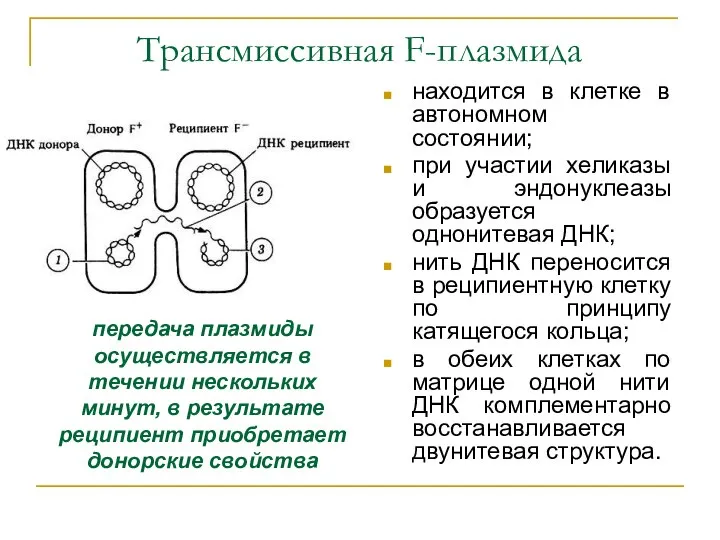
Трансмиссивная F-плазмида
находится в клетке в автономном состоянии;
при участии хеликазы и эндонуклеазы
Трансмиссивная F-плазмида
находится в клетке в автономном состоянии;
при участии хеликазы и эндонуклеазы
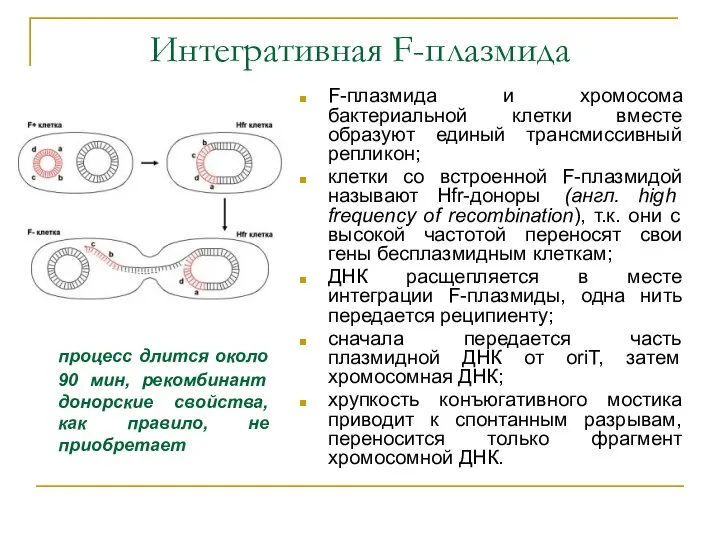
Интегративная F-плазмида
F-плазмида и хромосома бактериальной клетки вместе образуют единый трансмиссивный репликон;
клетки
Интегративная F-плазмида
F-плазмида и хромосома бактериальной клетки вместе образуют единый трансмиссивный репликон;
клетки

Трансформация
поглощение фрагментов ДНК и включение в хромосому бактерии реципиента;
ДНК должна быть
Трансформация
поглощение фрагментов ДНК и включение в хромосому бактерии реципиента;
ДНК должна быть

Механизм трансформации
Аутолизин разрушает КС, обнажает ДНК-связывающий белок и эндонуклеазу I;
ДНК-связывающий белок
Механизм трансформации
Аутолизин разрушает КС, обнажает ДНК-связывающий белок и эндонуклеазу I;
ДНК-связывающий белок

Трансдукция
передача генетического материала от одной бактерии другой посредством бактериофага
общая
Трансдукция
передача генетического материала от одной бактерии другой посредством бактериофага
общая

Экспрессия генетической информации у бактерий
Экспрессия генетической информации у бактерий

Бактерии – универсальные существа
Отдельная бактериальная клетка – полноценный самодостаточный организм;
Клетки
Бактерии – универсальные существа
Отдельная бактериальная клетка – полноценный самодостаточный организм;
Клетки

Парадокс бактериальной клетки
Высокая метаболическая активность (правило Рубнера);
Высокая ферментативная насыщенность (до 109-1012
Парадокс бактериальной клетки
Высокая метаболическая активность (правило Рубнера);
Высокая ферментативная насыщенность (до 109-1012

Быстрая адаптация к изменяющимся условиям среды - необходимое условие выживания
Бактерии должны
Быстрая адаптация к изменяющимся условиям среды - необходимое условие выживания
Бактерии должны

Бактерии нуждаются в получении информации о своем окружении
Способы получения информации бактериями:
непосредственный
Бактерии нуждаются в получении информации о своем окружении
Способы получения информации бактериями:
непосредственный

Внешние сигналы определяют экспрессию генов
Сигналы распознаются и преобразуются;
Передаются генетическим структурам;
Эта
Внешние сигналы определяют экспрессию генов
Сигналы распознаются и преобразуются;
Передаются генетическим структурам;
Эта

Регуляторы экспрессии генетической информации
двухкомпонентные сигнальные системы. Широко распространенны среди прокариотических организмов.
Регуляторы экспрессии генетической информации
двухкомпонентные сигнальные системы. Широко распространенны среди прокариотических организмов.

Уровни регуляции метаболизма
На уровне транскрипции (путь от ДНК к РНК);
Регуляция
Уровни регуляции метаболизма
На уровне транскрипции (путь от ДНК к РНК);
Регуляция

Регуляторы экспрессии оперонов
Регуляторные белки
Продукты генов-регуляторов
Связываются с участком ДНК -оператором;
Осуществляют позитивный или
Регуляторы экспрессии оперонов
Регуляторные белки
Продукты генов-регуляторов
Связываются с участком ДНК -оператором;
Осуществляют позитивный или

Эффекты связывания регуляторного белка с оператором
Негативный контроль регуляторный белок препятствует транскрипции
Фермент
Эффекты связывания регуляторного белка с оператором
Негативный контроль регуляторный белок препятствует транскрипции
Фермент

Индуцибельные опероны
Катаболитные опероны;
Цель регуляции – включить синтез ферментов, которые ранее не
Индуцибельные опероны
Катаболитные опероны;
Цель регуляции – включить синтез ферментов, которые ранее не

Катаболитная репрессия
Если в среде присутствует несколько субстратов (глюкоза и лактоза), то
Катаболитная репрессия
Если в среде присутствует несколько субстратов (глюкоза и лактоза), то

Механизм катаболитной репрессии
Эффектором выступает цАМФ;
Белок активатор катаболитных оперонов (БАК) неактивен в
Механизм катаболитной репрессии
Эффектором выступает цАМФ;
Белок активатор катаболитных оперонов (БАК) неактивен в

Циклический АМФ регулятор активности БАК
Образуется из АТФ с помощью аденилатциклазы;
При недостатке
Циклический АМФ регулятор активности БАК
Образуется из АТФ с помощью аденилатциклазы;
При недостатке

Арабинозный оперон
Гены araА, araВ, araD структурные – кодируют синтез ферментов, образуют
Арабинозный оперон
Гены araА, araВ, araD структурные – кодируют синтез ферментов, образуют
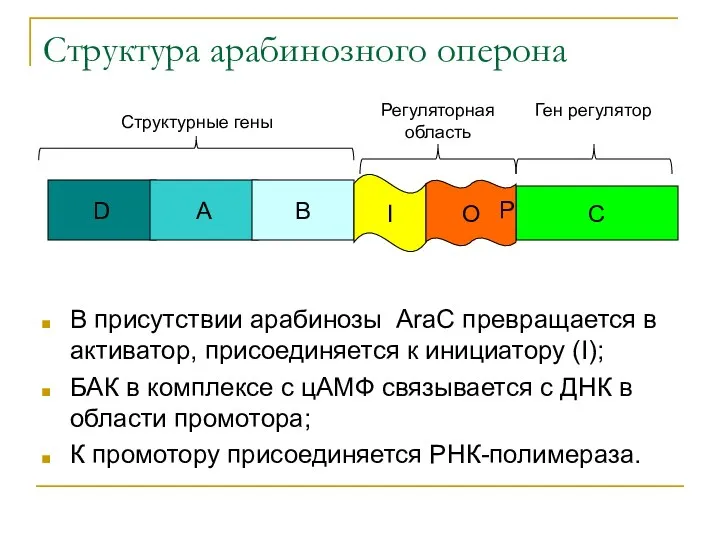
Структура арабинозного оперона
В присутствии арабинозы AraC превращается в активатор, присоединяется к
Структура арабинозного оперона
В присутствии арабинозы AraC превращается в активатор, присоединяется к
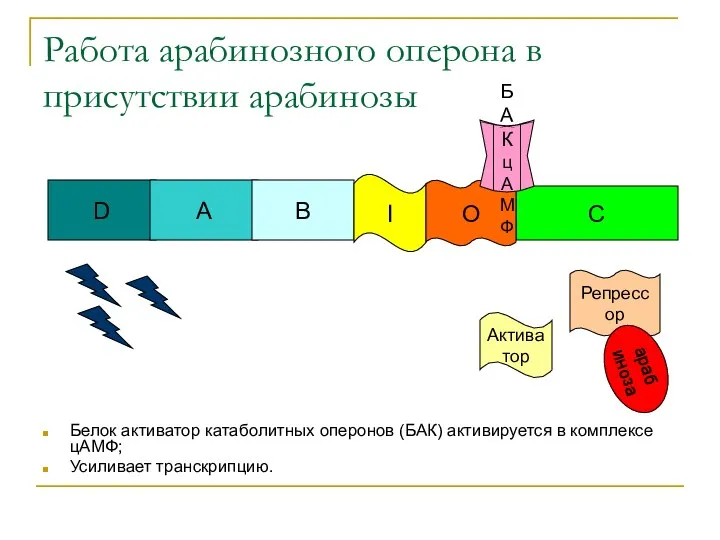
Работа арабинозного оперона в присутствии арабинозы
Репрессор
арабиноза
Актива
тор
БАК
цАМФ
Белок активатор катаболитных оперонов (БАК) активируется
Работа арабинозного оперона в присутствии арабинозы
Репрессор
арабиноза
Актива
тор
БАК
цАМФ
Белок активатор катаболитных оперонов (БАК) активируется

Работа арабинозного оперона в отсутствии арабинозы
Репрессор
Работа арабинозного оперона в отсутствии арабинозы
Репрессор

Репрессибельные опероны
Характерны для анаболических путей.
Цель регуляции – прекращение синтеза;
Оперон
Репрессибельные опероны
Характерны для анаболических путей.
Цель регуляции – прекращение синтеза;
Оперон

Триптофановый оперон
При наличии в среде триптофана, его синтез прекращается (репрессия конечным
Триптофановый оперон
При наличии в среде триптофана, его синтез прекращается (репрессия конечным
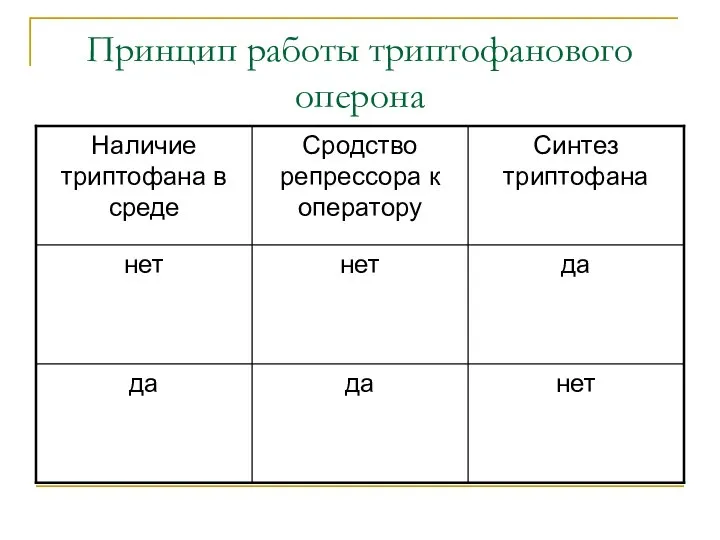
Принцип работы триптофанового оперона
Принцип работы триптофанового оперона

Аттенуация – механизм тонкой регуляции экспрессии структурных генов
Позволяет регулировать количество синтезирующегося
Аттенуация – механизм тонкой регуляции экспрессии структурных генов
Позволяет регулировать количество синтезирующегося
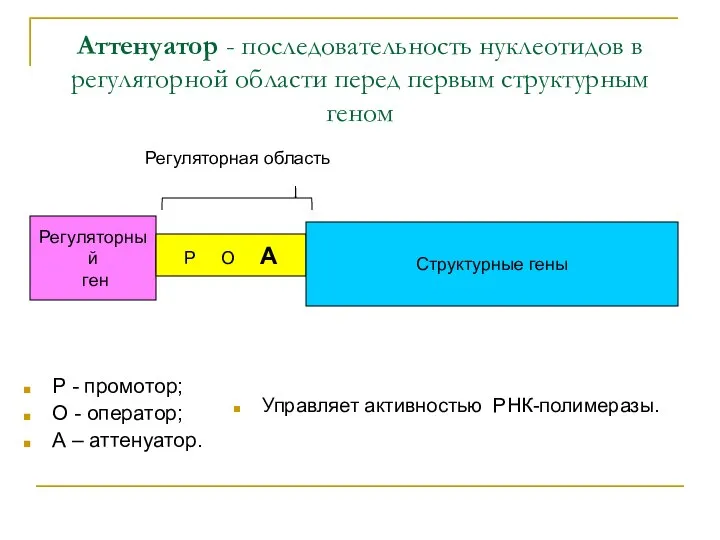
Аттенуатор - последовательность нуклеотидов в регуляторной области перед первым структурным геном
Р
Аттенуатор - последовательность нуклеотидов в регуляторной области перед первым структурным геном
Р
 Организация обучения биологии на основе вариативности УМК. Из опыта работы учителя биологии МОУ Маслянинской средней общеобраз
Организация обучения биологии на основе вариативности УМК. Из опыта работы учителя биологии МОУ Маслянинской средней общеобраз Автор Долгорукова С.В. учитель биологии и экологии, высшей категории
Автор Долгорукова С.В. учитель биологии и экологии, высшей категории Исследовательский проект. Сезонные явления в природе
Исследовательский проект. Сезонные явления в природе Черепно-мозговые нервы № 2
Черепно-мозговые нервы № 2 В экваториальном лесу Южной Америки
В экваториальном лесу Южной Америки Презентация на тему "Опыление (6 класс)" - скачать бесплатно презентации по Биологии
Презентация на тему "Опыление (6 класс)" - скачать бесплатно презентации по Биологии Спортивные газоны. Партерные газоны
Спортивные газоны. Партерные газоны Пищеварительная система
Пищеварительная система Птицы - лесные жители
Птицы - лесные жители Human Body
Human Body Дальтонизм. История термина
Дальтонизм. История термина Эстрогены.Биологические эффекты, механизм действия, биосинтез. Регуляция биосинтеза эстрогенов
Эстрогены.Биологические эффекты, механизм действия, биосинтез. Регуляция биосинтеза эстрогенов Rosengewächse (Pfirsich)
Rosengewächse (Pfirsich) Филогенез систем органов
Филогенез систем органов Презентация на тему Движение крови и лимфы в организме
Презентация на тему Движение крови и лимфы в организме  Общая ангиология
Общая ангиология Интродукция. Инвазивные виды
Интродукция. Инвазивные виды Отряд млекопитающие. Китообразные
Отряд млекопитающие. Китообразные Исчезнувшие животные Презентация по экологии на тему: «Исчезнувшие животные» Составила учитель биологии МОУ СОШ с. Апалиха, Х
Исчезнувшие животные Презентация по экологии на тему: «Исчезнувшие животные» Составила учитель биологии МОУ СОШ с. Апалиха, Х Значение выделения. 8ласс
Значение выделения. 8ласс Негативное воздействие металлов на организм человека Руководитель проекта: Иванова Е.В. Тебенькова Татьяна Михайловна учитель химии и биологии школы 255 кандидат биологических наук почетный работник образования РФ © 2007
Негативное воздействие металлов на организм человека Руководитель проекта: Иванова Е.В. Тебенькова Татьяна Михайловна учитель химии и биологии школы 255 кандидат биологических наук почетный работник образования РФ © 2007 Неандертальцы
Неандертальцы Особенности организации птиц
Особенности организации птиц Лекарственные и ядовитые растения
Лекарственные и ядовитые растения Insects № 4. Муравей
Insects № 4. Муравей Презентация на тему Кинология
Презентация на тему Кинология  Многообразие кишечнополостных, значение в природе, жизни человека
Многообразие кишечнополостных, значение в природе, жизни человека Микроскопические грибы
Микроскопические грибы