- Использование микроорганизмов домена архей в промышленной биотехнологии

Содержание
- 2. Введение Одним из основных достижений биологии конца XX века явилось выявление трех эволюционных ветвей жизни, сформировавшихся
- 3. Общая характеристика Архе́и (лат. Archaea от др.греч. - ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых
- 4. Морфология архей Средний диаметр клеток архей 1 мкм, типичный для большинства известных прокариот. Клетки Nanoarchaeum equitans
- 5. Сравнительная таблица архей и других доменов
- 6. Экология архей Археи распространены повсеместно. Начиная от гидротермальных источников, кратеров вулканов и дна Северного Ледовитого океана
- 7. Типы питания архей
- 8. Взаимодействие с другими организмами Метанообразующие археи вступают в симбиоз с термитами Хорошо изученные отношения между археями
- 9. Типы архей Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии; Euryarchaeota (Эвриархеоты) — метаногенные и галофильные археи;
- 10. Характеристика некоторых типов архей Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии Включает 28 родов, относящихся к
- 11. Euryarchaeota (Эвриархеоты) — метаногенные и галофильные археи Эти организмы распространены повсеместно. Некоторые формы являются экстремально термофильными
- 12. Применение архей в биотехнологии В настоящее время некоторые метаболиты архейного происхождения нашли применение или рекомендованы для
- 13. Процессы с использованием архей ДЕСУЛЬФУРИЗАЦИЯ ВЫХЛОПНЫХ ГАЗОВ На мощных заводах или химических предприятиях в ряде технических
- 14. Архейные ферменты используемые в биотехнологии ТЕРМОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ Одна из важнейших проблем, связанных с применением ферментов, —
- 15. Алкогольдегидрогеназы Ферменты кальдерных архей представляют интерес для органической химии. Хиральные спирты, применяемые в фармацевтике, получают при
- 16. а-Глюкозидазы Фермент включается на конечной стадии деградации гликогена, отщепляя терминальные остатки глюкозы. а-Глюкозидаза, выделенная из S.solfataricus,
- 17. ПСИХРОФИЛЫ И ПСИХРОФИЛЬНЫЕЭКСТРЕМОЗИМЫ Места обитания организмов с высокой температурой не являются широко распространенными на Земле, в
- 18. ГАЛОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ Две группы галофильных архей: аэробные галоархеи и анаэробные галофильные метаноархеи используют различные способы существования
- 19. ЭКСТРЕМОЗИМЫ АРХЕЙНЫХ ПЬЕЗОФИЛОВ Пьезофильные (барофильные) микроорганизмы, включая термофильные и гипертермофильные архейные штаммы, обитают при давлении в
- 20. Исследования по использованию архей ЭЛЕКТРОДНЫЕ МАТЕРИАЛЫ НА ОСНОВЕ МОДИФИЦИРОВАННЫХ ЖГУТИКОВ АРХЕЙ ДЛЯ ЛИТИЙ-ИОННЫХ АККУМУЛЯТОРОВ С помощью
- 22. Скачать презентацию

Введение
Одним из основных достижений биологии конца XX века явилось выявление трех
Введение
Одним из основных достижений биологии конца XX века явилось выявление трех

Общая характеристика
Архе́и (лат. Archaea от др.греч. - ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых организмов (по трехдоменной системе Карла Вёзе наряду
Общая характеристика
Архе́и (лат. Archaea от др.греч. - ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых организмов (по трехдоменной системе Карла Вёзе наряду

Морфология архей
Средний диаметр клеток архей 1 мкм, типичный для большинства известных
Морфология архей
Средний диаметр клеток архей 1 мкм, типичный для большинства известных

Сравнительная таблица архей и других доменов
Сравнительная таблица архей и других доменов

Экология архей
Археи распространены повсеместно. Начиная от гидротермальных источников, кратеров вулканов
Экология архей
Археи распространены повсеместно. Начиная от гидротермальных источников, кратеров вулканов

Типы питания архей
Типы питания архей

Взаимодействие с другими организмами
Метанообразующие археи вступают в симбиоз с термитами Хорошо изученные отношения
Взаимодействие с другими организмами
Метанообразующие археи вступают в симбиоз с термитами Хорошо изученные отношения

Типы архей
Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии;
Euryarchaeota (Эвриархеоты) — метаногенные и галофильные
Типы архей
Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии;
Euryarchaeota (Эвриархеоты) — метаногенные и галофильные

Характеристика некоторых типов архей
Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии Включает 28 родов, относящихся
Характеристика некоторых типов архей
Crenarchaeota(Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии Включает 28 родов, относящихся
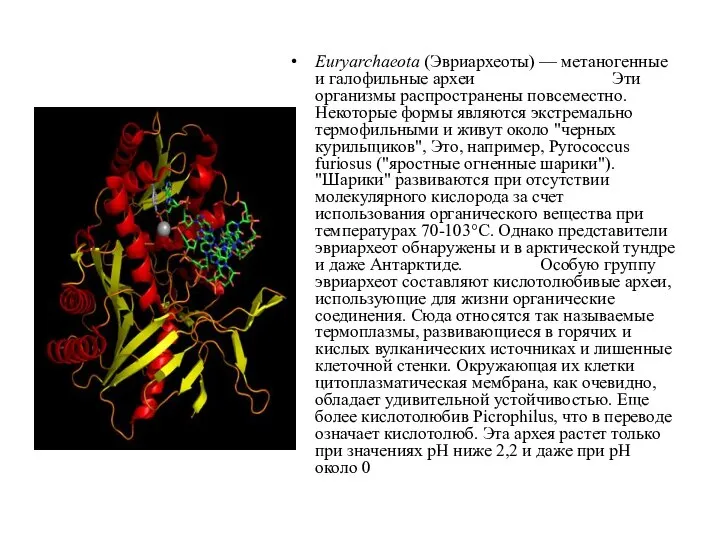
Euryarchaeota (Эвриархеоты) — метаногенные и галофильные археи Эти организмы распространены повсеместно. Некоторые формы
Euryarchaeota (Эвриархеоты) — метаногенные и галофильные археи Эти организмы распространены повсеместно. Некоторые формы

Применение архей в биотехнологии
В настоящее время некоторые метаболиты архейного происхождения нашли
Применение архей в биотехнологии
В настоящее время некоторые метаболиты архейного происхождения нашли

Процессы с использованием архей
ДЕСУЛЬФУРИЗАЦИЯ ВЫХЛОПНЫХ ГАЗОВ
На мощных заводах или химических предприятиях
Процессы с использованием архей
ДЕСУЛЬФУРИЗАЦИЯ ВЫХЛОПНЫХ ГАЗОВ
На мощных заводах или химических предприятиях

Архейные ферменты используемые в биотехнологии
ТЕРМОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ
Одна из важнейших проблем, связанных с
Архейные ферменты используемые в биотехнологии
ТЕРМОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ
Одна из важнейших проблем, связанных с

Алкогольдегидрогеназы
Ферменты кальдерных архей представляют интерес для органической химии. Хиральные спирты, применяемые
Алкогольдегидрогеназы
Ферменты кальдерных архей представляют интерес для органической химии. Хиральные спирты, применяемые
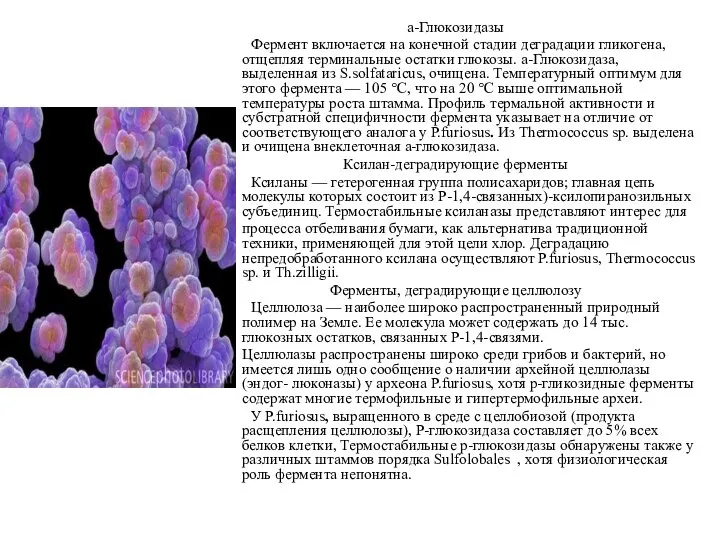
а-Глюкозидазы
Фермент включается на конечной стадии деградации гликогена, отщепляя терминальные остатки глюкозы.
а-Глюкозидазы
Фермент включается на конечной стадии деградации гликогена, отщепляя терминальные остатки глюкозы.

ПСИХРОФИЛЫ И ПСИХРОФИЛЬНЫЕЭКСТРЕМОЗИМЫ
Места обитания организмов с высокой температурой не являются широко
ПСИХРОФИЛЫ И ПСИХРОФИЛЬНЫЕЭКСТРЕМОЗИМЫ
Места обитания организмов с высокой температурой не являются широко

ГАЛОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ
Две группы галофильных архей: аэробные галоархеи и анаэробные галофильные метаноархеи
ГАЛОАРХЕЙНЫЕ ЭКСТРЕМОЗИМЫ
Две группы галофильных архей: аэробные галоархеи и анаэробные галофильные метаноархеи
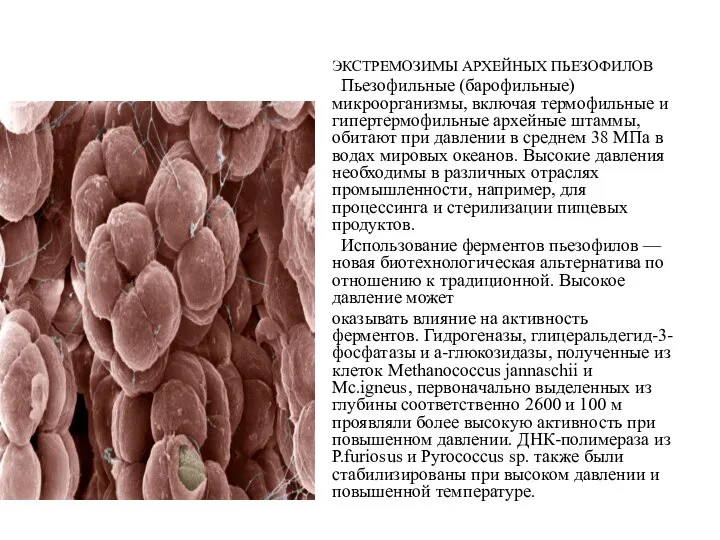
ЭКСТРЕМОЗИМЫ АРХЕЙНЫХ ПЬЕЗОФИЛОВ
Пьезофильные (барофильные) микроорганизмы, включая термофильные и гипертермофильные архейные штаммы,
ЭКСТРЕМОЗИМЫ АРХЕЙНЫХ ПЬЕЗОФИЛОВ
Пьезофильные (барофильные) микроорганизмы, включая термофильные и гипертермофильные архейные штаммы,

Исследования по использованию архей
ЭЛЕКТРОДНЫЕ МАТЕРИАЛЫ НА ОСНОВЕ МОДИФИЦИРОВАННЫХ ЖГУТИКОВ АРХЕЙ ДЛЯ
Исследования по использованию архей
ЭЛЕКТРОДНЫЕ МАТЕРИАЛЫ НА ОСНОВЕ МОДИФИЦИРОВАННЫХ ЖГУТИКОВ АРХЕЙ ДЛЯ
 Класс Двудольные: Семейства Пасленовые, Мотыльковые (Бобовые) и Сложноцветные (Астровые)
Класс Двудольные: Семейства Пасленовые, Мотыльковые (Бобовые) и Сложноцветные (Астровые) Сердце-центральный орган кровообращения.
Сердце-центральный орган кровообращения.  Презентация на тему Влияние солнечной активности на социальные процессы
Презентация на тему Влияние солнечной активности на социальные процессы  Презентация на тему Понятие о биоценозе
Презентация на тему Понятие о биоценозе  Жизнь в пресных водоемах
Жизнь в пресных водоемах Спинной мозг и спинномозговые нервы
Спинной мозг и спинномозговые нервы Экология животных. Тест
Экология животных. Тест Жизненные циклы простейших
Жизненные циклы простейших Патофизиология пәні; Қысқаша даму тарихы; Мақсаты мен міндеттері
Патофизиология пәні; Қысқаша даму тарихы; Мақсаты мен міндеттері Макромир: клетки, организмы, земные сообщества
Макромир: клетки, организмы, земные сообщества Высшая нервная деятельность
Высшая нервная деятельность Презентация на тему "ТИП ПРОСТЕЙШИЕ" - скачать бесплатно презентации по Биологии
Презентация на тему "ТИП ПРОСТЕЙШИЕ" - скачать бесплатно презентации по Биологии Презентация на тему Рыжий муравей
Презентация на тему Рыжий муравей Грибы - двойники. Как отличить ложные грибы-двойники от съедобных?
Грибы - двойники. Как отличить ложные грибы-двойники от съедобных? Высшие споровые растения
Высшие споровые растения Класс Земноводные
Класс Земноводные Своя игра. Рыбы, земноводные и пресмыкающиеся
Своя игра. Рыбы, земноводные и пресмыкающиеся Регуляция дыхания
Регуляция дыхания Степи, лесостепи
Степи, лесостепи Происхождение хордовых животных
Происхождение хордовых животных Аспирин: за и против
Аспирин: за и против Презентация на тему Три состояния воды
Презентация на тему Три состояния воды Принципы систематики и классификации растений
Принципы систематики и классификации растений Санитарная микробиология
Санитарная микробиология Хрящевые ткани
Хрящевые ткани Презентация на тему "Химия в сельском хозяйстве" - скачать презентации по Биологии
Презентация на тему "Химия в сельском хозяйстве" - скачать презентации по Биологии Долголетие и долгожитель
Долголетие и долгожитель Транспорт газов кровью
Транспорт газов кровью