- Клеточный цикл. Деление клетки. Митоз

Содержание
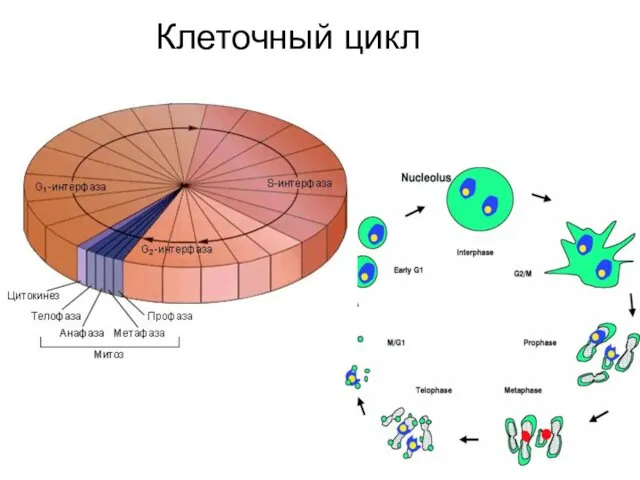
- 2. Клеточный цикл
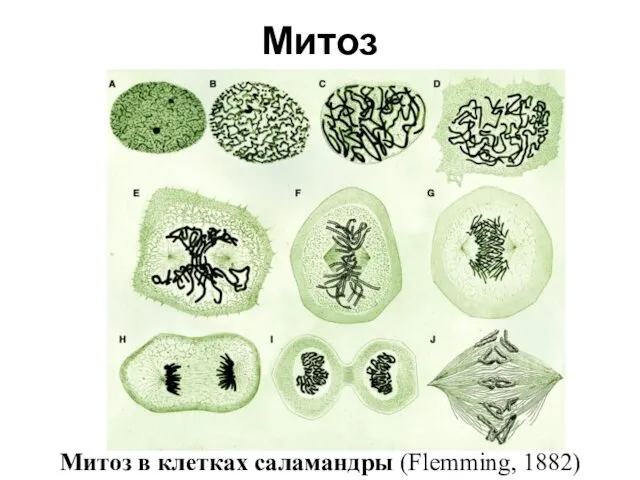
- 3. Митоз в клетках саламандры (Flemming, 1882) Митоз
- 4. 2. Конденсация хромосом – конденсин: -в основном - в митозе Когезия хромосом - когезин: -связывание в
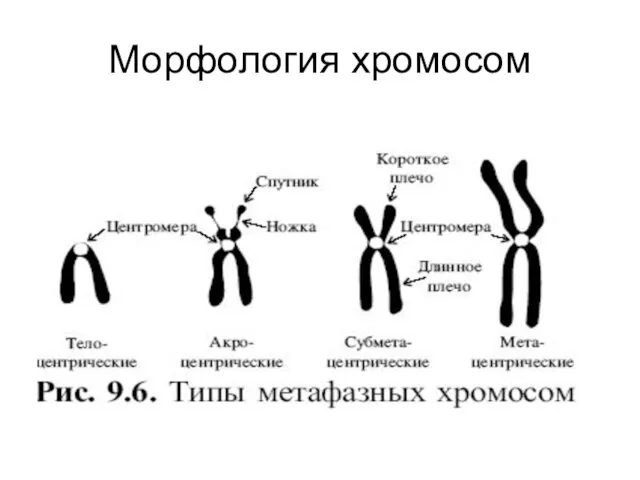
- 5. Морфология хромосом
- 7. ПРОФАЗА. В профазе происходят следующие события: 1.конденсация хромосом. Каждая хромосома двойная (2n4с), они тесно соприкасаются и
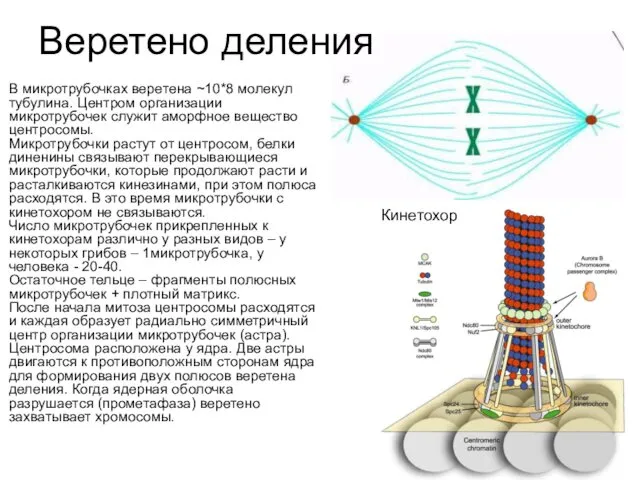
- 8. Веретено деления В микротрубочках веретена ~10*8 молекул тубулина. Центром организации микротрубочек служит аморфное вещество центросомы. Микротрубочки
- 9. Кинетохор Центромера и внутренний кинетохор Центральный кинетохор Внешний кинетохор Гетерохроматин Хромосомные плечи Кинетохор – мультибелковый комплекс,
- 10. Белковые моторы в составе кинетохора 1. Цитоплазматический динеин: движение к минус-концу МТ 2. CENP-E - кинезин:
- 11. МЕТАФАЗА 1. Завершается формирование веретена деления, 2. хромосомы перестают двигаться и выстраиваются по экватору веретена (экваториальная
- 12. АНАФАЗА Разделение хромосом Хромосомы внезапно одновременно отделяются друг от друга и начинают движение к полюсам. Центромеры
- 13. ТЕЛОФАЗА. Телофаза длится с момента прекращения движения хромосом. Происходит реконструкция ядер - образование ядерной оболочки, В
- 16. Мейоз Мейоз – происходит при образовании гамет - гаметный, при образовании спор - споровый и мейоз
- 20. Принципиальная схема мейоза. В соматических диплоидных клетках содержат две гомологичные хромосомы, одна отцовская и одна материнская.
- 21. Первое деление мейоза - редукционное Продолжительность мейоза напрямую зависит от количества ДНК в ядре. Также продолжительность
- 22. Лептотеннa (от лат. leptos - тонкий) - стадия длинных, тонких, слабо спирализованных хромосом, на которых видны
- 23. Профаза I Пахитена стадия толстых нитей (pachis - толстый). Гомологичные хромосомы, составляющие биваленты частично разделяются, как
- 24. Метафаза I завершается формирование веретена деления, его нити прикрепляются к центромерам хромосом, в результате чего биваленты
- 25. Анафаза I Связи в бивалентах ослабляются и гомологичные хромосомы отходят друг от друга, направляясь к противоположным
- 26. Телофаза I У полюсов веретена деления собирается одинарный гаплоидный набор хромосом, каждая из них содержит удвоенное
- 27. Второе деление мейоза - уравнительное (эквационное) Интеркинез - временной промежуток между первым и вторым делениями мейоза
- 28. Профаза II В профазу II исчезают ядрышки и кариолемма, хромосомы укорачиваются и утолщаются. Центриоли перемещаются к
- 29. Метафаза II Метафаза II. Хромосомы выстраиваются по экватору клетки. При этом хроматиды случайно взаимодействуют с одним
- 30. Анафаза II Анафаза II. Происходит деление центромер, хроматиды отделяются друг от друга и теперь называются хромосомами,
- 31. Телофаза II Телофаза II Очень сходна с телофазой митоза. Генетическая характеристика образующихся клеток 1n1c.
- 32. Значение мейоза: У организмов, размножающихся половым путем, в результате мейоза образуются гаплоидные клетки, хромосомные наборы которых
- 33. Деление бактериальной клетки
- 35. Скачать презентацию
Клеточный цикл
Клеточный цикл
Митоз в клетках саламандры (Flemming, 1882)
Митоз
Митоз в клетках саламандры (Flemming, 1882)
Митоз

2. Конденсация хромосом –
конденсин: -в основном - в митозе
2. Конденсация хромосом –
конденсин: -в основном - в митозе
Морфология хромосом
Морфология хромосом


ПРОФАЗА.
В профазе происходят следующие события:
1.конденсация хромосом. Каждая хромосома двойная (2n4с),
ПРОФАЗА.
В профазе происходят следующие события:
1.конденсация хромосом. Каждая хромосома двойная (2n4с),
Веретено деления
В микротрубочках веретена ~10*8 молекул тубулина. Центром организации микротрубочек служит
Веретено деления
В микротрубочках веретена ~10*8 молекул тубулина. Центром организации микротрубочек служит
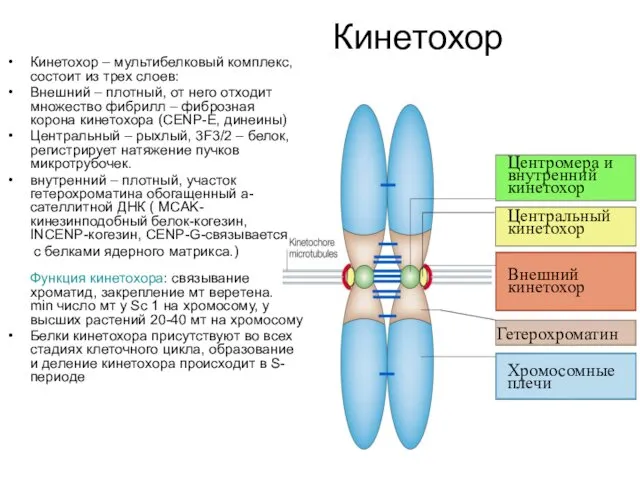
Кинетохор
Центромера и
внутренний
кинетохор
Центральный
кинетохор
Внешний
кинетохор
Гетерохроматин
Хромосомные
плечи
Кинетохор – мультибелковый комплекс, состоит из трех слоев:
Внешний
Кинетохор
Центромера и
внутренний
кинетохор
Центральный
кинетохор
Внешний
кинетохор
Гетерохроматин
Хромосомные
плечи
Кинетохор – мультибелковый комплекс, состоит из трех слоев:
Внешний
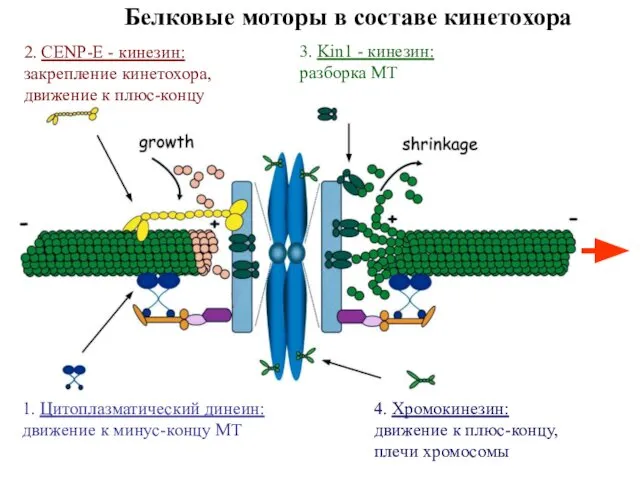
Белковые моторы в составе кинетохора
1. Цитоплазматический динеин:
движение к минус-концу МТ
2. CENP-E
Белковые моторы в составе кинетохора
1. Цитоплазматический динеин:
движение к минус-концу МТ
2. CENP-E

МЕТАФАЗА
1. Завершается формирование веретена деления,
2. хромосомы перестают двигаться и выстраиваются
МЕТАФАЗА
1. Завершается формирование веретена деления,
2. хромосомы перестают двигаться и выстраиваются

АНАФАЗА
Разделение хромосом
Хромосомы внезапно одновременно отделяются друг от друга и начинают
АНАФАЗА
Разделение хромосом
Хромосомы внезапно одновременно отделяются друг от друга и начинают

ТЕЛОФАЗА.
Телофаза длится с момента прекращения движения хромосом.
Происходит реконструкция ядер
ТЕЛОФАЗА.
Телофаза длится с момента прекращения движения хромосом.
Происходит реконструкция ядер


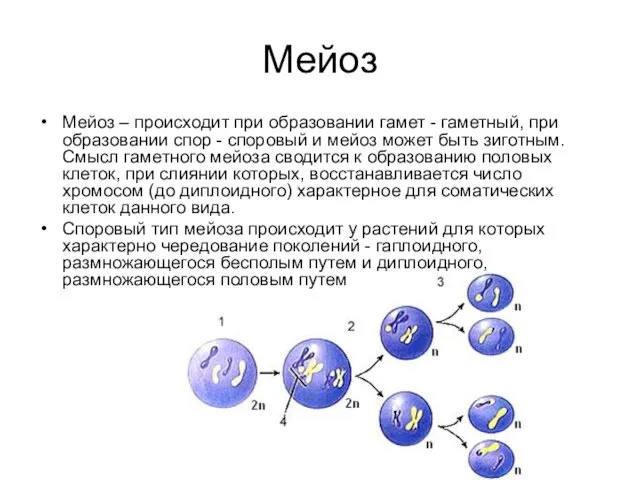
Мейоз
Мейоз – происходит при образовании гамет - гаметный, при образовании спор
Мейоз
Мейоз – происходит при образовании гамет - гаметный, при образовании спор



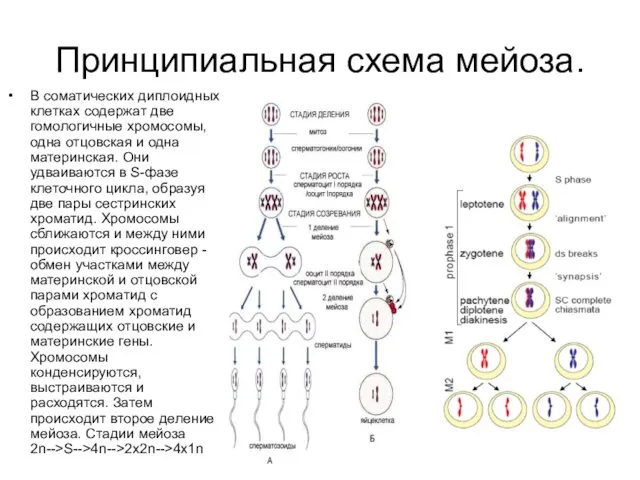
Принципиальная схема мейоза.
В соматических диплоидных клетках содержат две гомологичные хромосомы, одна
Принципиальная схема мейоза.
В соматических диплоидных клетках содержат две гомологичные хромосомы, одна
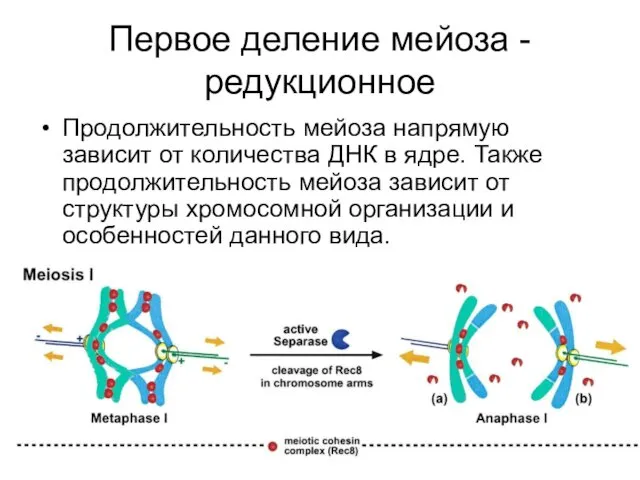
Первое деление мейоза - редукционное
Продолжительность мейоза напрямую зависит от количества ДНК
Первое деление мейоза - редукционное
Продолжительность мейоза напрямую зависит от количества ДНК
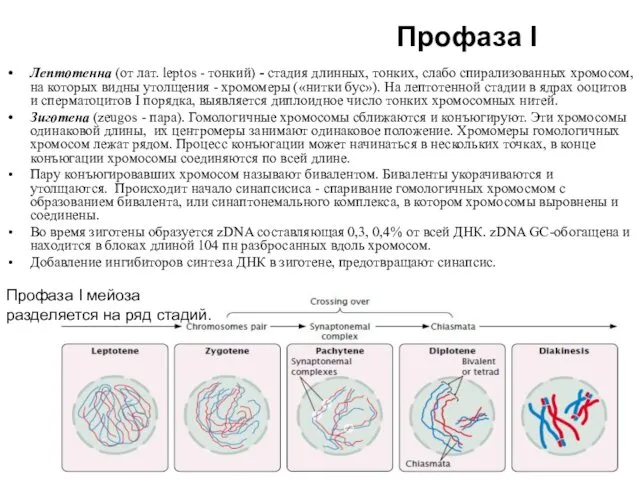
Лептотеннa (от лат. leptos - тонкий) - стадия длинных, тонких, слабо
Лептотеннa (от лат. leptos - тонкий) - стадия длинных, тонких, слабо

Профаза I
Пахитена стадия толстых нитей (pachis - толстый). Гомологичные хромосомы, составляющие
Профаза I
Пахитена стадия толстых нитей (pachis - толстый). Гомологичные хромосомы, составляющие

Метафаза I
завершается формирование веретена деления, его нити прикрепляются к центромерам хромосом,
Метафаза I
завершается формирование веретена деления, его нити прикрепляются к центромерам хромосом,

Анафаза I
Связи в бивалентах ослабляются и гомологичные хромосомы отходят друг от
Анафаза I
Связи в бивалентах ослабляются и гомологичные хромосомы отходят друг от

Телофаза I
У полюсов веретена деления собирается одинарный гаплоидный набор хромосом, каждая
Телофаза I
У полюсов веретена деления собирается одинарный гаплоидный набор хромосом, каждая
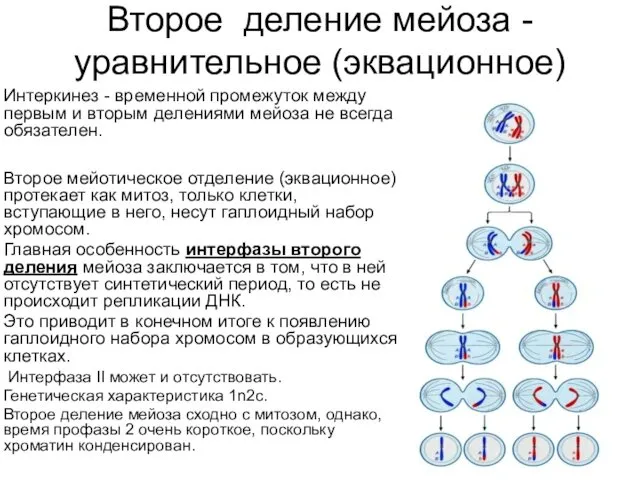
Второе деление мейоза - уравнительное (эквационное)
Интеркинез - временной промежуток между первым
Второе деление мейоза - уравнительное (эквационное)
Интеркинез - временной промежуток между первым
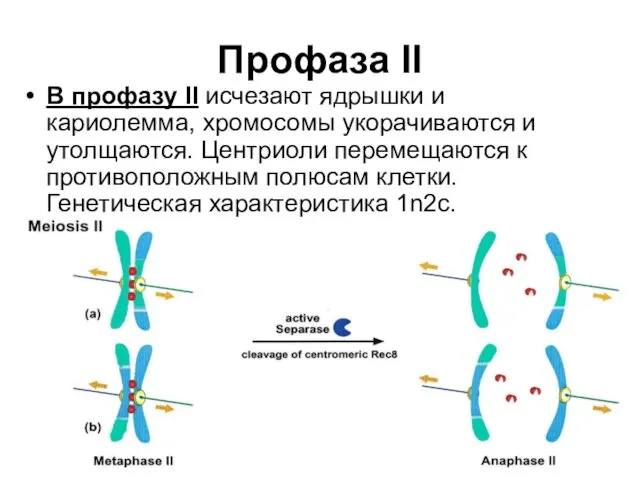
Профаза II
В профазу II исчезают ядрышки и кариолемма, хромосомы укорачиваются и
Профаза II
В профазу II исчезают ядрышки и кариолемма, хромосомы укорачиваются и

Метафаза II
Метафаза II. Хромосомы выстраиваются по экватору клетки. При этом хроматиды
Метафаза II
Метафаза II. Хромосомы выстраиваются по экватору клетки. При этом хроматиды

Анафаза II
Анафаза II. Происходит деление центромер, хроматиды отделяются друг от друга
Анафаза II
Анафаза II. Происходит деление центромер, хроматиды отделяются друг от друга
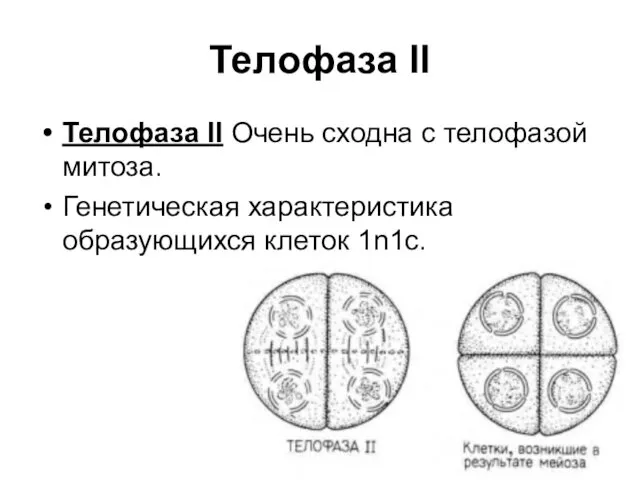
Телофаза II
Телофаза II Очень сходна с телофазой митоза.
Генетическая характеристика образующихся
Телофаза II
Телофаза II Очень сходна с телофазой митоза.
Генетическая характеристика образующихся
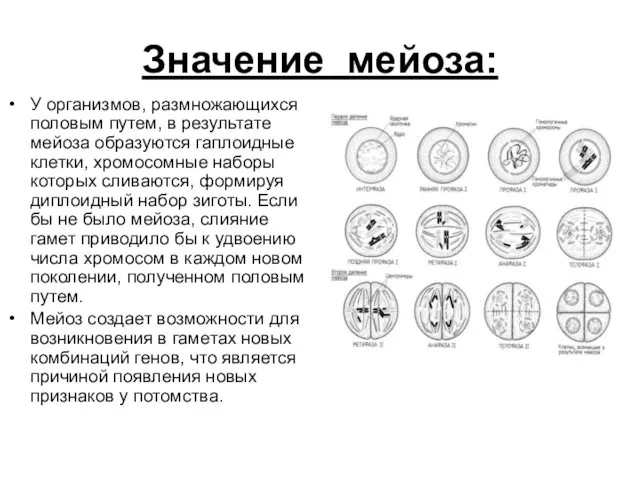
Значение мейоза:
У организмов, размножающихся половым путем, в результате мейоза образуются гаплоидные
Значение мейоза:
У организмов, размножающихся половым путем, в результате мейоза образуются гаплоидные
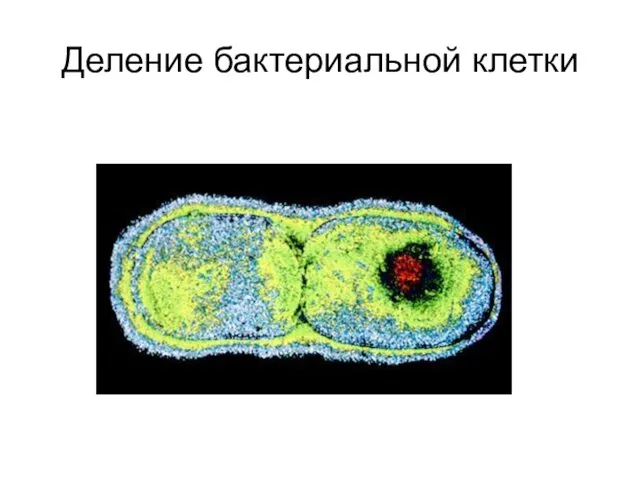
Деление бактериальной клетки
Деление бактериальной клетки
 Судың радиационды-биологиялық
Судың радиационды-биологиялық Зоологія. Холоднокровні
Зоологія. Холоднокровні Шар пей
Шар пей Жыныс генетикасы
Жыныс генетикасы Презентация на тему "Сукцессия" - скачать презентации по Биологии
Презентация на тему "Сукцессия" - скачать презентации по Биологии Витамин PP
Витамин PP Презентация на тему "Необычные растения" - скачать бесплатно презентации по Биологии
Презентация на тему "Необычные растения" - скачать бесплатно презентации по Биологии Последствия влияния алкоголя, никотина и наркотических веществ на развитие зародыша человека
Последствия влияния алкоголя, никотина и наркотических веществ на развитие зародыша человека Свойства живого вещества
Свойства живого вещества Строение тела человека. (9 класс)
Строение тела человека. (9 класс) Северный калан
Северный калан Высшая нервная деятельность по Павлову И.П
Высшая нервная деятельность по Павлову И.П Нейро-эндокринная регуляция иммунного ответа
Нейро-эндокринная регуляция иммунного ответа Неорганические вещества, входящие в состав клетки
Неорганические вещества, входящие в состав клетки  Бактерии. Полезные и вредные бактерии
Бактерии. Полезные и вредные бактерии Объёмный ёжик
Объёмный ёжик Развитие жизни на Земле: архей и протерозой
Развитие жизни на Земле: архей и протерозой Мышечная и нервная ткани. Нервные волокна и окончания
Мышечная и нервная ткани. Нервные волокна и окончания ВИДЫ ПИНГВИНОВ
ВИДЫ ПИНГВИНОВ  Ископаемые предки птиц
Ископаемые предки птиц Биоинженерия. Основные направления
Биоинженерия. Основные направления Реконструктивные посадки
Реконструктивные посадки Ехінококоз тварин та поширення його на території України
Ехінококоз тварин та поширення його на території України Детям о собаках
Детям о собаках Тип Arthropoda Членистоногие. Класс Ostracoda Остракоды (кембрий – кватер) (Ракушняковые раки)
Тип Arthropoda Членистоногие. Класс Ostracoda Остракоды (кембрий – кватер) (Ракушняковые раки) Популяционная экология животных
Популяционная экология животных Птицы Кузбасса
Птицы Кузбасса Части растения Окружающий мир ( 2 класс )
Части растения Окружающий мир ( 2 класс )