- Оплодотворение и дробление

Содержание
- 2. 1 – цитоплазма яйцеклетки; 2 – ядро яйцеклетки; 3 – блестящая оболочка; 4 – фолликулярный эпителий;
- 3. 1 фаза – СБЛИЖЕНИЕ Сперматозоиды двигаются благодаря: Хемотаксису (pH=7-7,5 – быстро перемещаются по направлению к яйцеклеткам,
- 4. 2 фаза – ПРОНИКНОВЕНИЕ У млекопитающих в яйцеклетку проникает лишь один сперматозоид - моноспермия. У беспозвоночных
- 5. А-В – слияние наружной мембраны акросомы и мембраны сперматозоида; Г-Д – полимеризация актина и образование акросомного
- 6. В цитоплазму яйцеклетки проникает головка, шейка и часть хвостового отдела (митохондриальное влагалище). Проникновение сперматозоида значительно усиливает
- 7. Медленный блок Сутью этой реакции является зкзоцитоз содержимого кортикальных гранул, которое оказывается между мембраной яйцеклетки и
- 8. Быстрый блок (ББ описан у морских ежей и у амфибий) Сразу после того, как первый сперматозоид
- 9. 3 фаза – СИНКАРИОН Образуются ♂ и ♀ пронуклеусы с последующим их слиянием. У многих видов
- 10. Метаболические события в яйце: Увеличивается вязкость цитоплазмы и ее проницаемость; Интенсивно перемещаются составные части ооплазмы, иногда
- 11. Слева – яйцо лягушки до оплодотворения; справа – изменения сразу после Ооплазматическая сегрегация 1 - анимальный
- 12. ДРОБЛЕНИЕ ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца Правила: Правило Пфлюгера: веретено всегда
- 13. ДРОБЛЕНИЕ Клетки, возникающие в результате дробления – бластомеры, а перетяжки, по которым они отделяются одна от
- 14. Равномерное дробление (алецитальные, изолецитальные яйца) – бластомеры дробятся синхронно, имеют одинаковые размеры, их количество удваивается при
- 15. Частичное (меробластическое) дробление Дискоидальное дробление (полилецитальные яйца) – дробится только часть цитоплазмы с ядром (на анимальном
- 16. Радиальное дробление (иглокожие, хордовые) - после первых делений зиготы бластомеры анимального полушария лежат строго над бластомерами
- 17. Билатеральное дробление (нематоды, а также для многих низшие хордовые, в том числе для асцидии, аппендикулярии). Характерной
- 18. При радиальном и спиральном дроблениях обычно различаются и сроки определения будущей «судьбы» клеток, т. е. того,
- 19. Типы бластулы Целобластула (образуется при полном дроблении) – полый шар, стенка которого (бластодерма) состоит из одного
- 22. Классификация иц по количеству желтка Полилецитальные — большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых)
- 23. Классификация яиц по расположению желтка Телолецитальные желток смещен к вегетативному полюсу. Противоположный полюс - анимальный (некоторые
- 25. Скачать презентацию

1 – цитоплазма яйцеклетки; 2 – ядро яйцеклетки; 3 – блестящая
1 – цитоплазма яйцеклетки; 2 – ядро яйцеклетки; 3 – блестящая
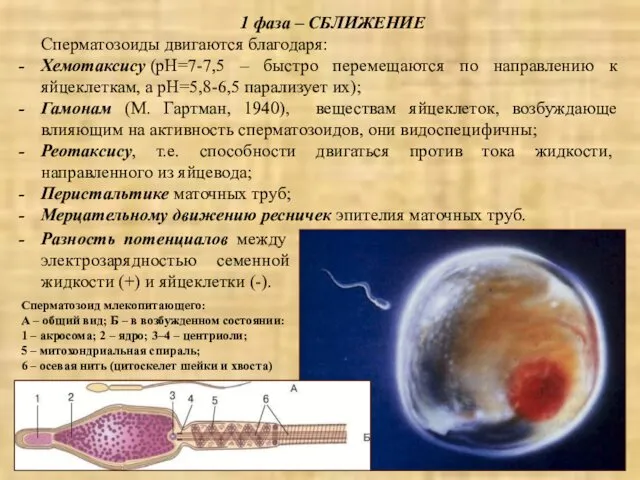
1 фаза – СБЛИЖЕНИЕ
Сперматозоиды двигаются благодаря:
Хемотаксису (pH=7-7,5 – быстро перемещаются по направлению
Сперматозоиды двигаются благодаря:
Хемотаксису (pH=7-7,5 – быстро перемещаются по направлению
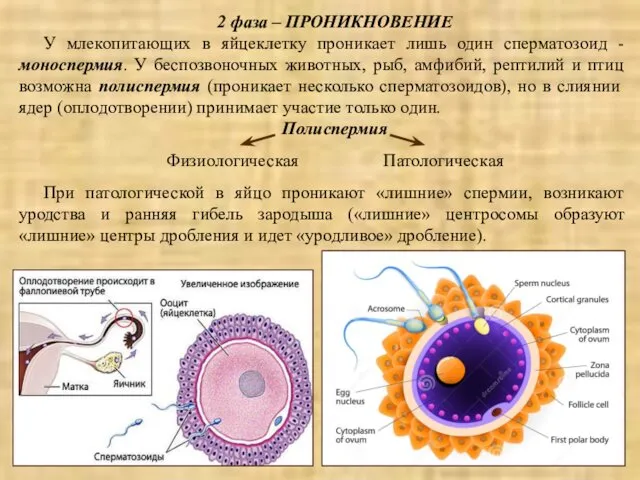
2 фаза – ПРОНИКНОВЕНИЕ
У млекопитающих в яйцеклетку проникает лишь один
2 фаза – ПРОНИКНОВЕНИЕ
У млекопитающих в яйцеклетку проникает лишь один

А-В – слияние наружной мембраны акросомы и мембраны сперматозоида;
Г-Д –
А-В – слияние наружной мембраны акросомы и мембраны сперматозоида;
Г-Д –
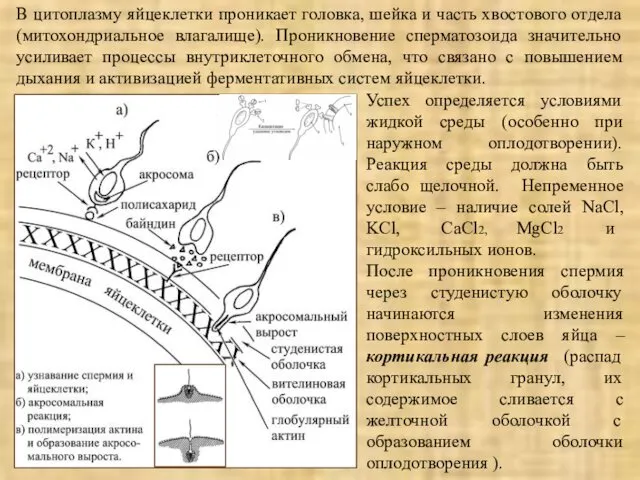
В цитоплазму яйцеклетки проникает головка, шейка и часть хвостового отдела (митохондриальное
В цитоплазму яйцеклетки проникает головка, шейка и часть хвостового отдела (митохондриальное
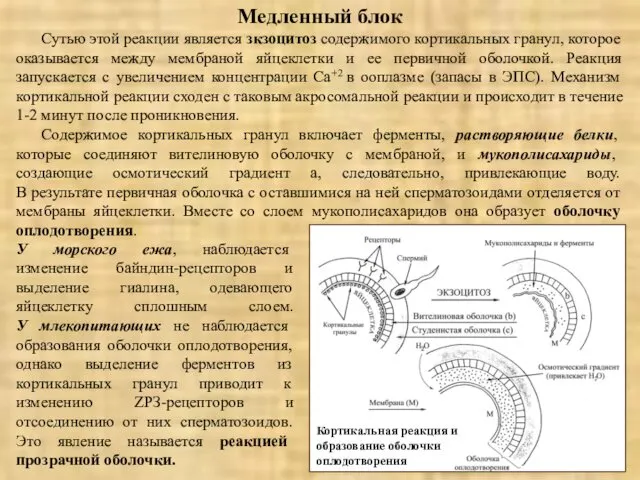
Медленный блок
Сутью этой реакции является зкзоцитоз содержимого кортикальных гранул, которое
Медленный блок
Сутью этой реакции является зкзоцитоз содержимого кортикальных гранул, которое
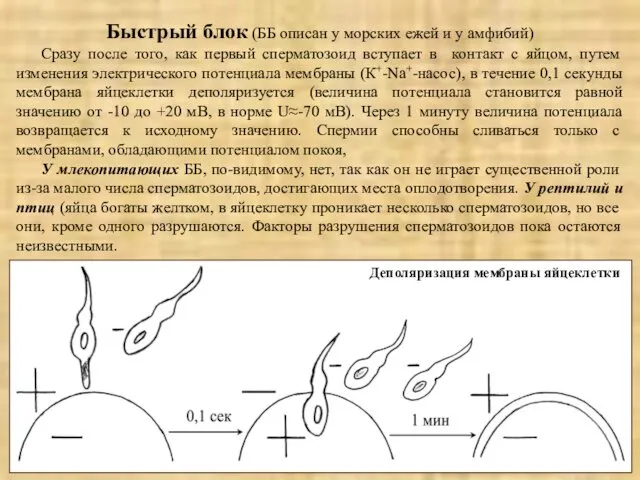
Быстрый блок (ББ описан у морских ежей и у амфибий)
Сразу после
Быстрый блок (ББ описан у морских ежей и у амфибий)
Сразу после

3 фаза – СИНКАРИОН
Образуются ♂ и ♀ пронуклеусы с последующим их
3 фаза – СИНКАРИОН
Образуются ♂ и ♀ пронуклеусы с последующим их

Метаболические события в яйце:
Увеличивается вязкость цитоплазмы и ее проницаемость;
Интенсивно перемещаются составные
Метаболические события в яйце:
Увеличивается вязкость цитоплазмы и ее проницаемость;
Интенсивно перемещаются составные

Слева – яйцо лягушки до оплодотворения;
справа – изменения сразу после
Ооплазматическая сегрегация
1
Слева – яйцо лягушки до оплодотворения;
справа – изменения сразу после
Ооплазматическая сегрегация
1

ДРОБЛЕНИЕ
ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца
Правила:
Правило Пфлюгера: веретено всегда
ДРОБЛЕНИЕ
ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца
Правила:
Правило Пфлюгера: веретено всегда

ДРОБЛЕНИЕ
Клетки, возникающие в результате дробления – бластомеры, а перетяжки, по которым
ДРОБЛЕНИЕ
Клетки, возникающие в результате дробления – бластомеры, а перетяжки, по которым
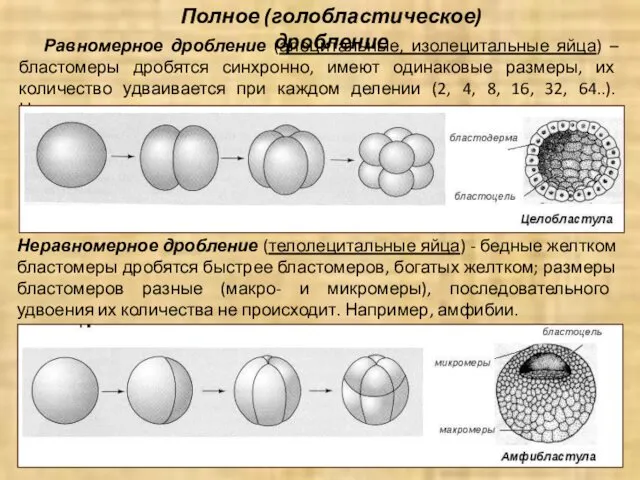
Равномерное дробление (алецитальные, изолецитальные яйца) – бластомеры дробятся синхронно, имеют одинаковые
Равномерное дробление (алецитальные, изолецитальные яйца) – бластомеры дробятся синхронно, имеют одинаковые

Частичное (меробластическое) дробление
Дискоидальное дробление (полилецитальные яйца) – дробится только часть
Частичное (меробластическое) дробление
Дискоидальное дробление (полилецитальные яйца) – дробится только часть

Радиальное дробление (иглокожие, хордовые) - после первых делений зиготы бластомеры анимального
Радиальное дробление (иглокожие, хордовые) - после первых делений зиготы бластомеры анимального

Билатеральное дробление (нематоды, а также для многих низшие хордовые, в том
Билатеральное дробление (нематоды, а также для многих низшие хордовые, в том

При радиальном и спиральном дроблениях обычно различаются и сроки определения будущей
При радиальном и спиральном дроблениях обычно различаются и сроки определения будущей
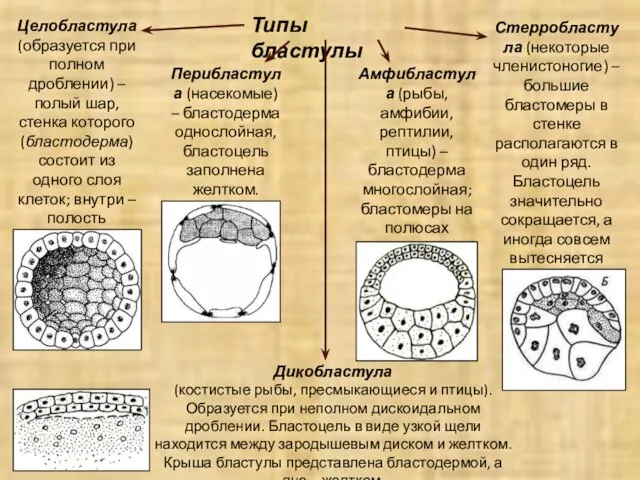
Типы бластулы
Целобластула (образуется при полном дроблении) – полый шар, стенка которого
Типы бластулы
Целобластула (образуется при полном дроблении) – полый шар, стенка которого



Классификация иц по количеству желтка
Полилецитальные — большое количество желтка (членистоногие, рептилии, птицы,
Классификация иц по количеству желтка
Полилецитальные — большое количество желтка (членистоногие, рептилии, птицы,

Классификация яиц по расположению желтка
Телолецитальные
желток смещен к вегетативному полюсу. Противоположный полюс
Классификация яиц по расположению желтка
Телолецитальные
желток смещен к вегетативному полюсу. Противоположный полюс
 Макроэволюция
Макроэволюция Строение костной системы организма и ее функции. Лекция №3
Строение костной системы организма и ее функции. Лекция №3 Опорно-двигательная система человека
Опорно-двигательная система человека Тесты по биологии
Тесты по биологии Рост и развитие животных
Рост и развитие животных Микробиологическая оценка обсемененности продуктов переработки яблок
Микробиологическая оценка обсемененности продуктов переработки яблок Хвойные растения
Хвойные растения  Строение и работа сердца
Строение и работа сердца Всем привет Поговорим о теориях происхождения жизни.
Всем привет Поговорим о теориях происхождения жизни. Теплорегуляция Интегрированный урок (физики и биологии)
Теплорегуляция Интегрированный урок (физики и биологии) Гены развития дрозофилы
Гены развития дрозофилы Презентация Полевые сорняки
Презентация Полевые сорняки Влияние породы коров на физико-химичесский состав молока и качество сливочного масла. Оценка племенных животных. Мечение скота
Влияние породы коров на физико-химичесский состав молока и качество сливочного масла. Оценка племенных животных. Мечение скота Природа вокруг нас
Природа вокруг нас Гены в хромосомах и популяциях
Гены в хромосомах и популяциях Презентация на тему «Лекарственные и ядовитые растения» Презентацию подготовила ученица 6 А класса МОУ-СОШ № 33 Резнеченко Ангел
Презентация на тему «Лекарственные и ядовитые растения» Презентацию подготовила ученица 6 А класса МОУ-СОШ № 33 Резнеченко Ангел Споры и Спорангии
Споры и Спорангии Презентация на тему "Семейство Крестоцветные" - скачать презентации по Биологии
Презентация на тему "Семейство Крестоцветные" - скачать презентации по Биологии Пищеварительный канал
Пищеварительный канал Селекция микроорганизмов
Селекция микроорганизмов Международный день птиц
Международный день птиц Рани. Їх класифікація. Ярової М. Авак’ян О.
Рани. Їх класифікація. Ярової М. Авак’ян О.  Генетика пола. Наследование, сцепленное с полом
Генетика пола. Наследование, сцепленное с полом История развития, основные достижения и проблемы медицинской генетики. Цитологические основы наследственности
История развития, основные достижения и проблемы медицинской генетики. Цитологические основы наследственности Презентация на тему Ежи морские (Echinoidea)
Презентация на тему Ежи морские (Echinoidea)  Мутагенные факторы
Мутагенные факторы Взаимоотношения между организмами хищничество
Взаимоотношения между организмами хищничество Выполнил: учитель биологии МБОУ ООШ п.Туголесский Бор Солдакова Марина Александровна
Выполнил: учитель биологии МБОУ ООШ п.Туголесский Бор Солдакова Марина Александровна