Основы электрофизиологии. Общая характеристика возбудимых тканей. Потенциал покоя. Потенциал действия
- Основы электрофизиологии. Общая характеристика возбудимых тканей. Потенциал покоя. Потенциал действия

Содержание
- 2. 2 Основные проявления жизнедеятельности Физиологический покой Физиологическая активность Раздражение Возбуждение Торможение
- 3. 3 Общая характеристика возбудимых тканей. Все клетки нашего организма обладают свойством раздражимости. Раздражимость – это способность
- 4. 4 Раздражение и виды раздражителей. Раздражение – это процесс воздействия на живой объект внешних по отношению
- 5. 5 По природе раздражителя: 1. физические ( температура, звук, свет, электрический ток и т.д.). 2. химические
- 6. 6 По биологическому значению: 1. адекватные – раздражитель к которому в естественных условиях у биосистем есть
- 7. 7 По количественному признаку или порогу раздражения: В зависимости от силы действия раздражители делятся на: 1.
- 8. 8 Все возбудимые ткани обладают общими свойствами: Возбудимостью Проводимостью Лабильностью Аккомодацией
- 9. 9 Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной вызывать возбуждение. Проводимость –
- 10. 10 Законы раздражения: Закон силы Закон времени Закон крутизны нарастания раздражителя
- 11. 11 Закон силы Чем сильней раздражение, тем до определенного предела сильнее ответная реакция биосистемы. Закон постулирован
- 12. 12 Закон времени Чем длительнее раздражение, тем сильней до определенных пределов ответная реакция живой ткани. В
- 13. 13 ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ» Этот закон был экспериментально установллен и сформулирован независимо друг от друга
- 14. 14 Закон крутизны нарастания раздражителя или градиента: Чем выше крутизна нарастания раздражителя во времени, тем больше
- 15. 15
- 16. История открытия биоэлектри чества.
- 18. 18 Первый опыт Гальвани При подвешивании нервно - мышечного препарата с помощью медного крючка, проходящего через
- 19. 19 ВОЛЬТОВ СТОЛБ Вольтов столб, состоящий из металлических дисков, разделенных кружками мокрой ткани. Однако ему возразил
- 20. 20 Второй опыт Гальвани Позднее, защищая свою точку зрения от возражений оппонентов, Гальвани воспроизводит различные модификации
- 21. 21 Опыт Маттеучи Открытия Гальвани были подтверждены в работах Маттеучи (1837). Однако, Маттеучи обнаружил явление вторичного
- 22. 22 Работы Э.Дюбуа-Реймона ( выполненны в 1840-1860 гг. ) В этих работах благодаря высокочувствительному гальванометру и
- 23. Природа и происхождение потенциал покоя (Ео) 0 -75 +30 мв Ео
- 24. 24 В 1896 г. В.Ю. Чаговец впервые высказал гипотезу о ионном механизме электрических потенциалов в живых
- 25. 25 Диффузионный концентрационный потенциал Представим сосуд, разделенный ис-кусственной полупроницаемой мембраной. Стенки пор этой мембраны заряжены электроотрицательно,
- 26. 26 Двойной электрический слой (мембрана поляризована)
- 27. 27 Уравнение НЕРНСТА RT [ K +o] Ек = ℓn nF [ K +i] Ек –
- 28. 28 Английский зоолог Д. Юнг (1936) обнаружил, что особые длинные тяжи у кальмаров и каракатиц являются
- 29. 29 Экспериментальная проверка гипотезы Бернштейна В 1962 г. Ходжкин с сотрудниками подтвердил гипотезу Бернштейна: Из аксона
- 30. 30 Современные представления о природе биоэлектрических явлений в тканях базируются на результатах работ Алана Ходжкина, Эндрью
- 32. 32 Мембранно-ионная теория происхождения потенциала покоя (ПП) Все клетки живого организма поляризованы. Между внутренней средой клетки
- 33. 33 Схема регистрации мембранного потенциала клетки
- 34. 34
- 35. 35 Концентрации Na+, K+ и Cl– внутри и вне клетки и пути проникновения этих ионов через
- 36. 36 Физиологический мембранный потенциал в возбудимых клетках называется потенциалом покоя (ПП)
- 37. 37
- 38. 38 Если подставить значения концентраций из таблицы, то: ЕK+ = - 97 мВ ЕNa+ = +
- 39. 39 Уравнение ГОЛЬДМАНА RT Pk [ Ko+] + PNa [ Nao+]+PCl [ Cli- ] Ем =
- 40. 40 Потенциал покоя создается: Неравновесным распределением калия, натрия, хлора и кальция внутри клетки по отношению к
- 41. 41 Три состояния мембраны 1. Поляризация - значение МП в состоянии покоя. 2. Деполяризация - уменьшение
- 42. 42 Два варианта ответной реакции возбудимой структуры на действие раздражителя: 1. локальный ответ (ЛО)- на действие
- 43. Потенциал действия
- 44. ПД – это быстрое изменение мембранного потенциала, которые возникают в месте нанесения раздражения и распространяются на
- 45. 45
- 46. 46 Натриевая гипотеза происхождения ПД При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко
- 47. 47 Экспериментальное доказательство натриевой природы деполяризации В 1939 г. Ходжкин и Хаксли установили, что мембранный потенциал
- 48. 48 Мембрана характеризуется повышенной проницаемостью для ионов Na+ лишь очень короткое время 0.2 - 0.5 мс.
- 49. 49 Проницаемость мембраны для ионов Na на пике ПД pK+ : pNa+ : pCl - =
- 50. 50
- 51. 51 ЦИКЛ ХОДЖКИНА-ХАКСЛИ Раздражитель деполяризация мембраны возрастание повышение Na+ входящего проницаемости Na+ - тока
- 52. m h m h h Р Е А К Т И В А Ц И Я
- 53. 53
- 54. 54 Потенциалзависимая инактивация Na+ –системы По оси абсцисс отложены величины отклонения мембранного потенциала от потенциала покоя
- 55. 55 Nа+ - потенциалозависимые каналы, в мембране нервного и скелетного мышечного волокна, характеризуются, как время -
- 56. Блокада калиевых каналов тетраэтиламмонием резко удлиняет процесс реполяризации m m m 56
- 57. 57
- 58. 58
- 59. 59 Потенциал действия: Возникает в ответ на пороговые и сверхпороговые раздражители. Обусловлен увеличением проницаемости мембраны для
- 61. Скачать презентацию
2
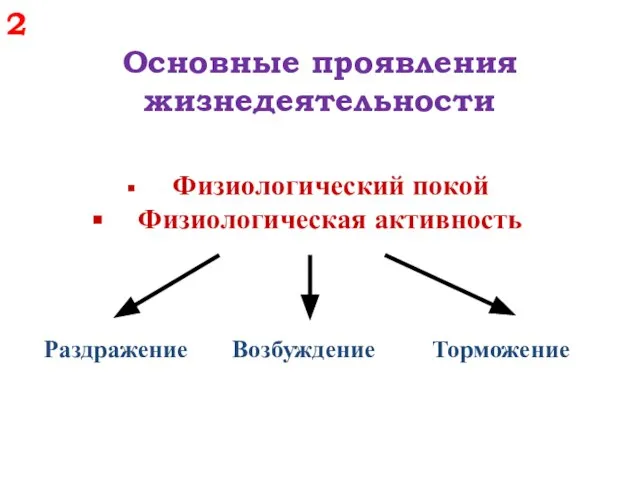
Основные проявления
жизнедеятельности
Физиологический покой
Физиологическая активность
Раздражение Возбуждение Торможение
2
Основные проявления
жизнедеятельности
Физиологический покой
Физиологическая активность
Раздражение Возбуждение Торможение

3
Общая характеристика возбудимых тканей.
Все клетки нашего организма обладают свойством раздражимости.
Раздражимость
3
Общая характеристика возбудимых тканей.
Все клетки нашего организма обладают свойством раздражимости.
Раздражимость

4
Раздражение и виды раздражителей.
Раздражение – это процесс воздействия на живой объект
4
Раздражение и виды раздражителей.
Раздражение – это процесс воздействия на живой объект

5
По природе раздражителя:
1. физические ( температура, звук, свет, электрический ток
5
По природе раздражителя:
1. физические ( температура, звук, свет, электрический ток

6
По биологическому значению:
1. адекватные – раздражитель к которому в естественных
6
По биологическому значению:
1. адекватные – раздражитель к которому в естественных

7
По количественному признаку или порогу раздражения:
В зависимости от силы действия раздражители
7
По количественному признаку или порогу раздражения:
В зависимости от силы действия раздражители

8
Все возбудимые ткани обладают
общими свойствами:
Возбудимостью
Проводимостью
Лабильностью
Аккомодацией
8
Все возбудимые ткани обладают
общими свойствами:
Возбудимостью
Проводимостью
Лабильностью
Аккомодацией

9
Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной
9
Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной

10
Законы раздражения:
Закон силы
Закон времени
Закон крутизны нарастания раздражителя
10
Законы раздражения:
Закон силы
Закон времени
Закон крутизны нарастания раздражителя

11
Закон силы
Чем сильней раздражение, тем до определенного предела сильнее ответная
11
Закон силы
Чем сильней раздражение, тем до определенного предела сильнее ответная

12
Закон времени
Чем длительнее раздражение, тем сильней до определенных пределов
12
Закон времени
Чем длительнее раздражение, тем сильней до определенных пределов

13
ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
Этот закон был экспериментально установллен и сформулирован
13
ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
Этот закон был экспериментально установллен и сформулирован

14
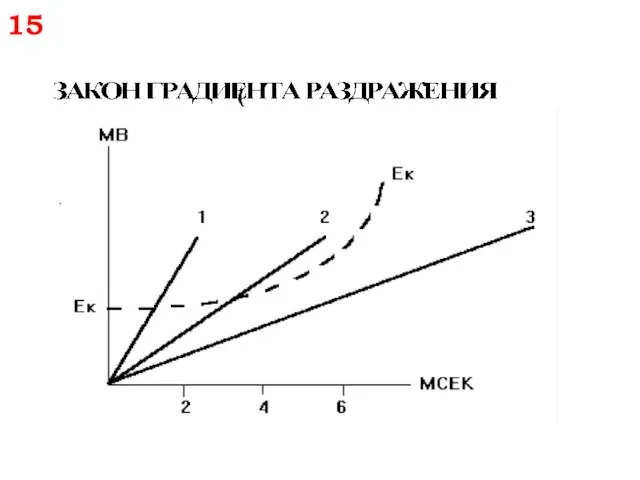
Закон крутизны нарастания раздражителя или градиента:
Чем выше крутизна нарастания раздражителя во
14
Закон крутизны нарастания раздражителя или градиента:
Чем выше крутизна нарастания раздражителя во
15
15

История открытия
биоэлектри чества.
История открытия
биоэлектри чества.


18
Первый опыт Гальвани
При подвешивании нервно - мышечного препарата с помощью
18
Первый опыт Гальвани
При подвешивании нервно - мышечного препарата с помощью

19 ВОЛЬТОВ СТОЛБ
Вольтов столб, состоящий из металлических дисков, разделенных кружками мокрой
19 ВОЛЬТОВ СТОЛБ
Вольтов столб, состоящий из металлических дисков, разделенных кружками мокрой

20
Второй опыт Гальвани
Позднее, защищая свою точку зрения от возражений оппонентов, Гальвани
20
Второй опыт Гальвани
Позднее, защищая свою точку зрения от возражений оппонентов, Гальвани
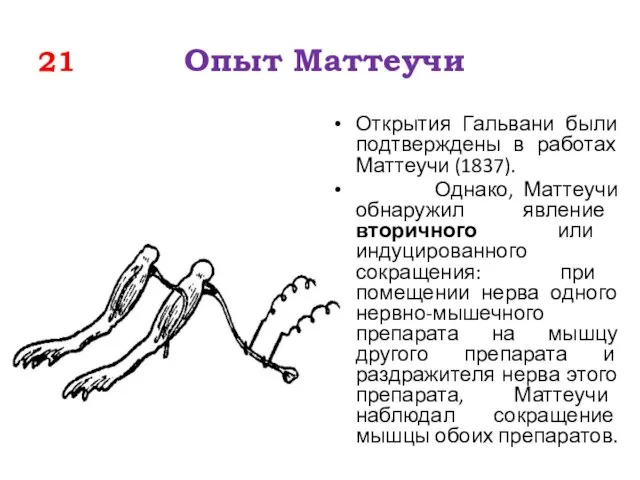
21 Опыт Маттеучи
Открытия Гальвани были подтверждены в работах Маттеучи (1837).
21 Опыт Маттеучи
Открытия Гальвани были подтверждены в работах Маттеучи (1837).

22
Работы Э.Дюбуа-Реймона
( выполненны в 1840-1860 гг. )
В этих
22
Работы Э.Дюбуа-Реймона
( выполненны в 1840-1860 гг. )
В этих

Природа и происхождение потенциал покоя (Ео)
0
-75
+30
мв
Ео
Природа и происхождение потенциал покоя (Ео)
0
-75
+30
мв
Ео

24
В 1896 г. В.Ю. Чаговец впервые высказал гипотезу о ионном
24
В 1896 г. В.Ю. Чаговец впервые высказал гипотезу о ионном

25 Диффузионный концентрационный потенциал
Представим сосуд, разделенный ис-кусственной полупроницаемой мембраной. Стенки пор
25 Диффузионный концентрационный потенциал
Представим сосуд, разделенный ис-кусственной полупроницаемой мембраной. Стенки пор

26
Двойной электрический слой
(мембрана поляризована)
26
Двойной электрический слой
(мембрана поляризована)
![27 Уравнение НЕРНСТА RT [ K +o] Ек = ℓn nF](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/533645/slide-26.jpg)
27
Уравнение НЕРНСТА
RT [ K +o]
Ек = ℓn
nF [
27
Уравнение НЕРНСТА
RT [ K +o]
Ек = ℓn
nF [

28
Английский зоолог
Д. Юнг (1936) обнаружил, что особые длинные тяжи у
28 Английский зоолог Д. Юнг (1936) обнаружил, что особые длинные тяжи у

29
Экспериментальная проверка гипотезы Бернштейна
В 1962 г. Ходжкин с сотрудниками подтвердил гипотезу
29
Экспериментальная проверка гипотезы Бернштейна
В 1962 г. Ходжкин с сотрудниками подтвердил гипотезу

30
Современные представления о природе биоэлектрических явлений в тканях базируются на результатах
30
Современные представления о природе биоэлектрических явлений в тканях базируются на результатах


32
Мембранно-ионная теория происхождения
потенциала покоя (ПП)
Все клетки живого организма поляризованы. Между
32
Мембранно-ионная теория происхождения
потенциала покоя (ПП)
Все клетки живого организма поляризованы. Между
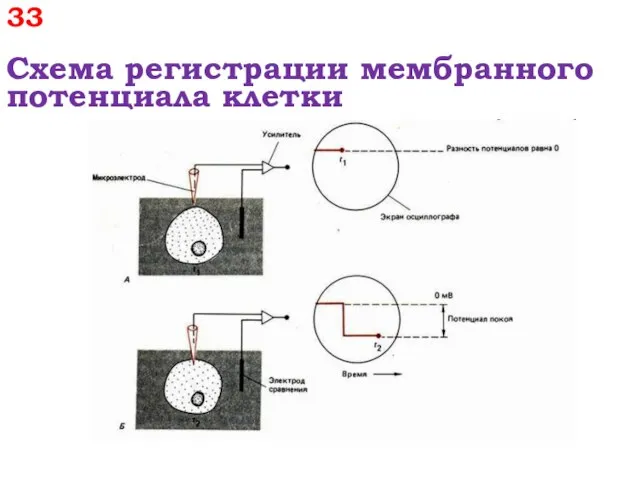
33
Схема регистрации мембранного потенциала клетки
33
Схема регистрации мембранного потенциала клетки
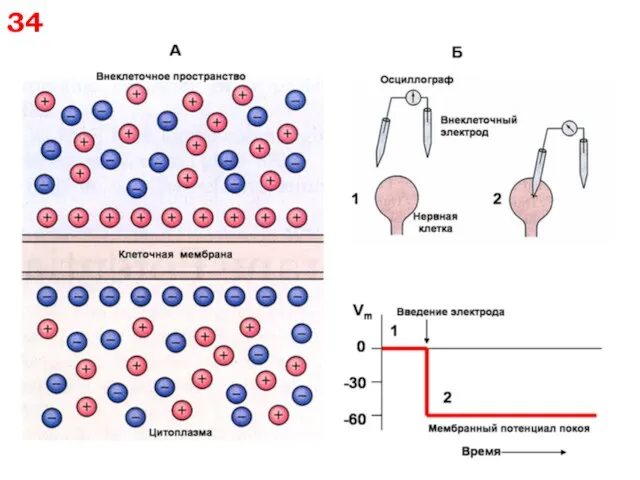
34
34

35
Концентрации Na+, K+ и Cl– внутри и вне клетки и пути
35 Концентрации Na+, K+ и Cl– внутри и вне клетки и пути

36
Физиологический мембранный потенциал
в возбудимых клетках называется потенциалом покоя (ПП)
36
Физиологический мембранный потенциал
в возбудимых клетках называется потенциалом покоя (ПП)
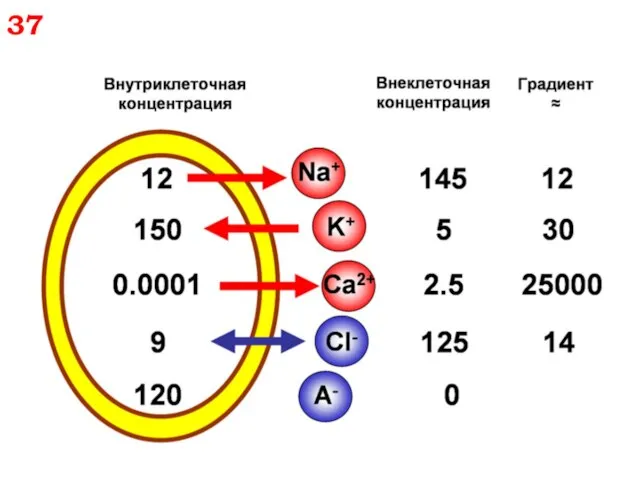
37
37

38
Если подставить значения концентраций из таблицы, то:
ЕK+ = - 97 мВ
ЕNa+
38
Если подставить значения концентраций из таблицы, то:
ЕK+ = - 97 мВ
ЕNa+
![39 Уравнение ГОЛЬДМАНА RT Pk [ Ko+] + PNa [ Nao+]+PCl](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/533645/slide-38.jpg)
39 Уравнение ГОЛЬДМАНА
RT Pk [ Ko+] + PNa [ Nao+]+PCl
39 Уравнение ГОЛЬДМАНА
RT Pk [ Ko+] + PNa [ Nao+]+PCl

40
Потенциал покоя создается:
Неравновесным распределением калия, натрия, хлора и кальция внутри
40
Потенциал покоя создается:
Неравновесным распределением калия, натрия, хлора и кальция внутри

41
Три состояния мембраны
1. Поляризация - значение МП в состоянии покоя.
2.
41
Три состояния мембраны
1. Поляризация - значение МП в состоянии покоя.
2.

42
Два варианта ответной реакции возбудимой структуры на действие раздражителя:
1. локальный ответ
42
Два варианта ответной реакции возбудимой структуры на действие раздражителя:
1. локальный ответ

Потенциал действия
Потенциал действия

ПД – это быстрое изменение мембранного потенциала, которые возникают в месте
ПД – это быстрое изменение мембранного потенциала, которые возникают в месте

45
45

46
Натриевая гипотеза происхождения ПД
При действии на клетку раздражителя проницаемость мембраны
46
Натриевая гипотеза происхождения ПД
При действии на клетку раздражителя проницаемость мембраны

47
Экспериментальное доказательство натриевой природы деполяризации
В 1939 г. Ходжкин и Хаксли установили,
47
Экспериментальное доказательство натриевой природы деполяризации
В 1939 г. Ходжкин и Хаксли установили,

48
Мембрана характеризуется повышенной проницаемостью для ионов Na+ лишь очень короткое
48
Мембрана характеризуется повышенной проницаемостью для ионов Na+ лишь очень короткое

49
Проницаемость мембраны для ионов Na на пике ПД
pK+ : pNa+ :
49
Проницаемость мембраны для ионов Na на пике ПД
pK+ : pNa+ :

50
50
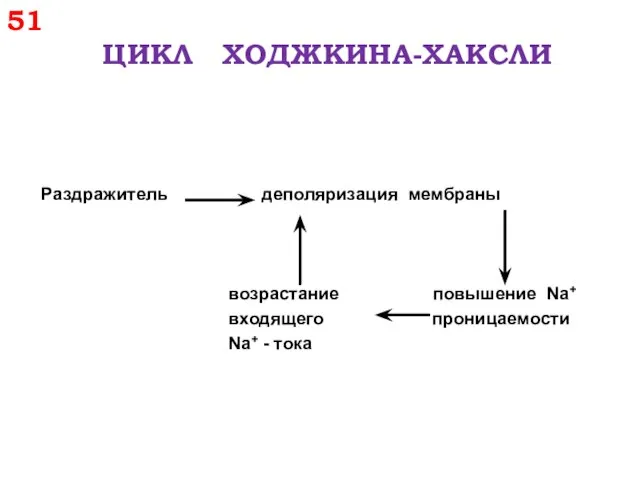
51
ЦИКЛ ХОДЖКИНА-ХАКСЛИ
Раздражитель деполяризация мембраны
возрастание повышение Na+
входящего проницаемости
Na+
51
ЦИКЛ ХОДЖКИНА-ХАКСЛИ
Раздражитель деполяризация мембраны
возрастание повышение Na+
входящего проницаемости
Na+
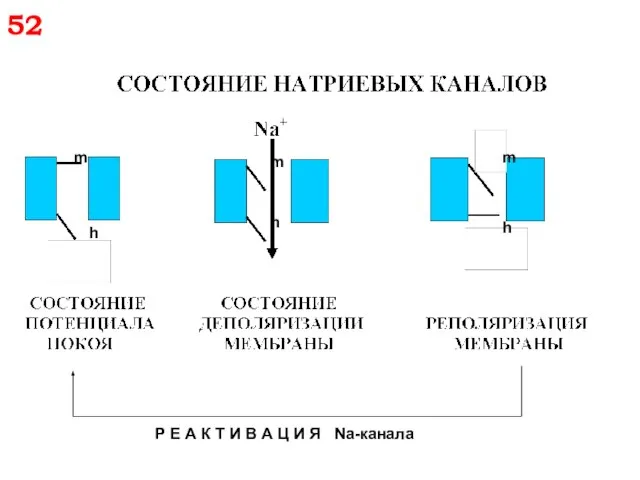
m
h
m
h
h
Р Е А К Т И В А Ц И Я
m
h
m
h
h
Р Е А К Т И В А Ц И Я

53
53
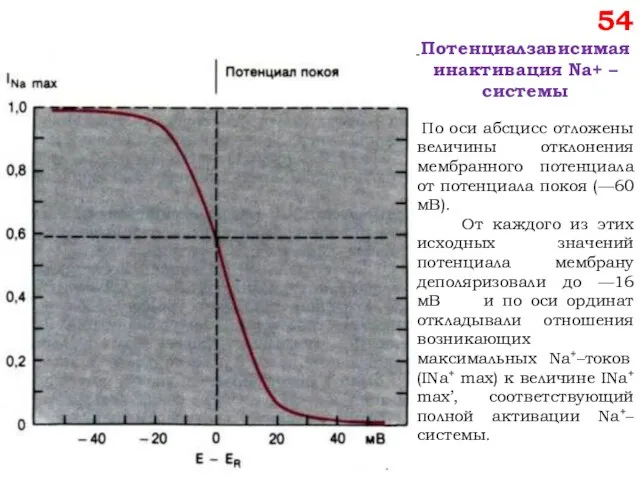
54
Потенциалзависимая инактивация Na+ –системы
По оси абсцисс отложены величины отклонения
54
Потенциалзависимая инактивация Na+ –системы
По оси абсцисс отложены величины отклонения

55
Nа+ - потенциалозависимые каналы, в мембране нервного и скелетного мышечного волокна,
55
Nа+ - потенциалозависимые каналы, в мембране нервного и скелетного мышечного волокна,

Блокада калиевых каналов тетраэтиламмонием
резко удлиняет процесс реполяризации
m
m
m
56
Блокада калиевых каналов тетраэтиламмонием
резко удлиняет процесс реполяризации
m
m
m
56
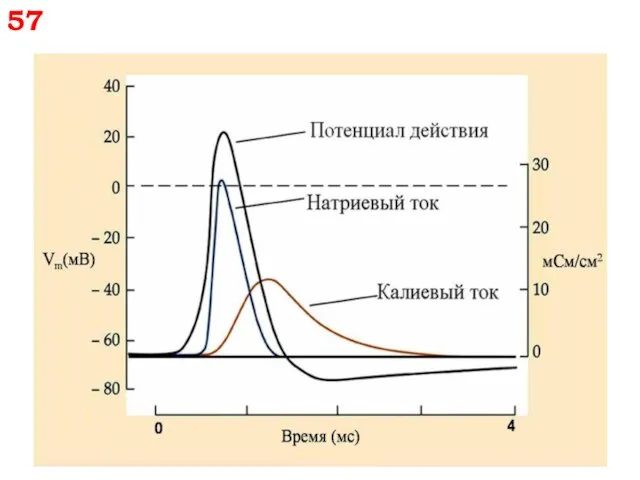
57
57

58
58

59
Потенциал действия:
Возникает в ответ на пороговые и сверхпороговые раздражители.
Обусловлен увеличением
59
Потенциал действия:
Возникает в ответ на пороговые и сверхпороговые раздражители.
Обусловлен увеличением
 Презентация Живые организмы
Презентация Живые организмы Презентация Космический мусор
Презентация Космический мусор Высшие споровые растения. Мхи
Высшие споровые растения. Мхи Царство – растения Отдел – цветковые (покрытосеменные) Класс – двудольные Семейство – Крестоцветные.
Царство – растения Отдел – цветковые (покрытосеменные) Класс – двудольные Семейство – Крестоцветные. ЦИТОЛОГИЯ Наука о клетке История открытия клетки Строение клеток
ЦИТОЛОГИЯ Наука о клетке История открытия клетки Строение клеток Творчество Виктора Петровича Астафьева
Творчество Виктора Петровича Астафьева Презентация на тему "Простейшие (Корненожки, Радиолярии, Солнечники, Споровики)" - скачать презентации по Биологии
Презентация на тему "Простейшие (Корненожки, Радиолярии, Солнечники, Споровики)" - скачать презентации по Биологии КРОВЬ ЧЕЛОВЕКА И ЛЯГУШКИ, СТРОЕНИЕ КРОВЕТВОРНЫХ ОРГАНОВ, ГЕМОПОЭЗ
КРОВЬ ЧЕЛОВЕКА И ЛЯГУШКИ, СТРОЕНИЕ КРОВЕТВОРНЫХ ОРГАНОВ, ГЕМОПОЭЗ  Морфология побега высших растений. Побеговые системы. (Лекция 7)
Морфология побега высших растений. Побеговые системы. (Лекция 7) Презентация на тему Зелёная аптека
Презентация на тему Зелёная аптека Вітамин С
Вітамин С Митохондрии, пероксисомы и пластиды
Митохондрии, пероксисомы и пластиды Скелет человека
Скелет человека Лёгкие — органы воздушного дыхания у человека
Лёгкие — органы воздушного дыхания у человека Основы анатомии и физиологии сельскохозяйствен ных животных. Тема 2
Основы анатомии и физиологии сельскохозяйствен ных животных. Тема 2 Польові дослідження
Польові дослідження Презентация на тему "Архейская эра" - скачать презентации по Биологии
Презентация на тему "Архейская эра" - скачать презентации по Биологии Какая у нас память Исследовательская работа
Какая у нас память Исследовательская работа  Презентация на тему "Каменный лес" - скачать презентации по Биологии
Презентация на тему "Каменный лес" - скачать презентации по Биологии Сенсорная система. Органы чувств
Сенсорная система. Органы чувств Презентация на тему "звезды" - скачать бесплатно презентации по Биологии
Презентация на тему "звезды" - скачать бесплатно презентации по Биологии Презентация на тему "Влияние алкоголя на беременную женщину" - скачать бесплатно презентации по Биологии
Презентация на тему "Влияние алкоголя на беременную женщину" - скачать бесплатно презентации по Биологии Презентация на тему "Животные, о существовании которых вы может даже и не знали" - скачать бесплатно презентации по Биологии
Презентация на тему "Животные, о существовании которых вы может даже и не знали" - скачать бесплатно презентации по Биологии Влияние света на интенсивность дыхания
Влияние света на интенсивность дыхания Конкуренция белок!
Конкуренция белок! Селекция животных “Борька” – гибрид домашней козы и сибирского козерога.
Селекция животных “Борька” – гибрид домашней козы и сибирского козерога. Проведение потенциалов в мембранах нервных клеток
Проведение потенциалов в мембранах нервных клеток Среды обитания организмов
Среды обитания организмов