- Сократительная деятельность мышц

Содержание
- 2. План 1. Теории мышечного сокращения. 2. Одиночное и тетаническое сокращение. 3. Теории тетануса. 4. Формы и
- 4. Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон Каждое волокно – это клетка
- 6. В миофибриллах скелетных мышц наблюдается правильное чередование более светлых и более темных участков. Поэтому скелетные мышцы
- 8. Саркоплазматический ретикулум состоит из системы поперечных, продольных трубочек и цистерн. Поперечные трубочки – это впячивания саркоплазмы
- 9. Саркомер ограничен с двух сторон так называемыми Z-дисками. К этим дискам с двух сторон прикреплены тонкие
- 11. В А-дисках миозиновые нити образуют гексагональную упаковку, именно они обусловливают высокую оптическую плотность дисков. Актиновые нити
- 15. В одной половине толстой протофибриллы (и в А-диске) все молекулы ориентированы в одну сторону, а во
- 16. Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
- 17. Хаксли и Хэнсон – теория скольжения нитей.
- 18. В ходе сокращения миозин становится способным взаимодействовать с актином (нити актина тянет к центру). При этом
- 19. Схема сокращения 1 – актиновый филламент, 2 – центр связывания, 3 – миозиновый филламент, 4 –
- 20. Сокращение мышцы
- 22. Главной «моторной» частью миозина скелетных мышц является головка тяжелой цепи миозина. Эти головки могут дотягиваться до
- 25. Предполагается, что поперечные мостики, отходящие от толстой нити прикрепляются к тонкой нити под определенным углом, а
- 26. Механизм сопряжения возбуждения и сокращения 1 – поперечная трубочка саркоплазматической мембраны, 2 –саркоплазматический ретикулум, 3 –
- 27. ATP – АТФ ADP - АДФ Pi – ион фосфора
- 29. Actin Myosin Crossbridge 3D Animation*
- 30. Поступление потенциала действия в моторные концевые пластинки аксона приводи к высвобождению ацетилхолина из запасных пузырьков в
- 31. Нервный импульс передается поперечными трубочкам, где он трансформируется во взаимодействия между дигидропиридиновыми и рианодиновыми рецепторами, что
- 32. Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере
- 33. При одиночном мышечном сокращении различают 3 фазы: латентный период продолжительностью около 5 мс – промежуток между
- 36. Одиночное сокращение и тетанус При воздействии импульсов, наносимых с частотой более 20 в 1 с, в
- 37. Если период следования импульсов больше продолжительности одиночного сокращения (100 мс), то возникает серия одиночных мышечных сокращений
- 38. Used by permission of John W. Kimball Н.Е.Введенский проводя опыты на нервно-мышечном препарате лягушки установил, что
- 40. Происхождение зубчатого и гладкого тетануса. Б – фазы и периоды мышечного сокращения, Б – режимы мышечного
- 42. Скачать презентацию

План
1. Теории мышечного сокращения.
2. Одиночное и тетаническое сокращение.
3. Теории тетануса.
4.
План
1. Теории мышечного сокращения.
2. Одиночное и тетаническое сокращение.
3. Теории тетануса.
4.


Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон
Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон


В миофибриллах скелетных мышц наблюдается правильное чередование более светлых и более
В миофибриллах скелетных мышц наблюдается правильное чередование более светлых и более


Саркоплазматический ретикулум состоит из системы поперечных, продольных трубочек и цистерн. Поперечные
Саркоплазматический ретикулум состоит из системы поперечных, продольных трубочек и цистерн. Поперечные

Саркомер ограничен с двух сторон так называемыми Z-дисками. К этим дискам
Саркомер ограничен с двух сторон так называемыми Z-дисками. К этим дискам


В А-дисках миозиновые нити образуют гексагональную упаковку, именно они обусловливают высокую
В А-дисках миозиновые нити образуют гексагональную упаковку, именно они обусловливают высокую




В одной половине толстой протофибриллы (и в А-диске) все молекулы ориентированы
В одной половине толстой протофибриллы (и в А-диске) все молекулы ориентированы
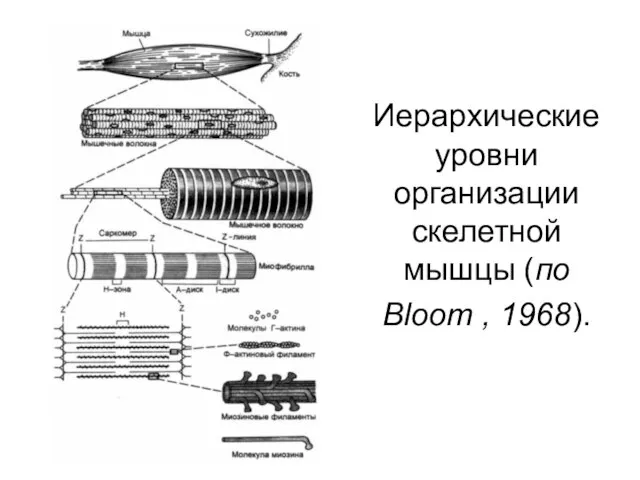
Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
Иерархические уровни организации скелетной мышцы (по Bloom , 1968).

Хаксли и Хэнсон –
теория скольжения нитей.
Хаксли и Хэнсон –
теория скольжения нитей.

В ходе сокращения миозин становится способным взаимодействовать с актином (нити актина
В ходе сокращения миозин становится способным взаимодействовать с актином (нити актина

Схема сокращения
1 – актиновый филламент,
2 – центр связывания,
3
Схема сокращения
1 – актиновый филламент, 2 – центр связывания, 3

Сокращение мышцы
Сокращение мышцы


Главной «моторной» частью миозина скелетных мышц является головка тяжелой цепи миозина.
Главной «моторной» частью миозина скелетных мышц является головка тяжелой цепи миозина.



Предполагается, что поперечные мостики, отходящие от толстой нити прикрепляются к тонкой
Предполагается, что поперечные мостики, отходящие от толстой нити прикрепляются к тонкой

Механизм сопряжения возбуждения и сокращения
1 – поперечная трубочка саркоплазматической мембраны,
Механизм сопряжения возбуждения и сокращения
1 – поперечная трубочка саркоплазматической мембраны,

ATP – АТФ
ADP - АДФ
Pi – ион фосфора
ATP – АТФ
ADP - АДФ
Pi – ион фосфора


Actin Myosin Crossbridge 3D Animation*
Actin Myosin Crossbridge 3D Animation*

Поступление потенциала действия в моторные концевые пластинки аксона приводи к высвобождению
Поступление потенциала действия в моторные концевые пластинки аксона приводи к высвобождению
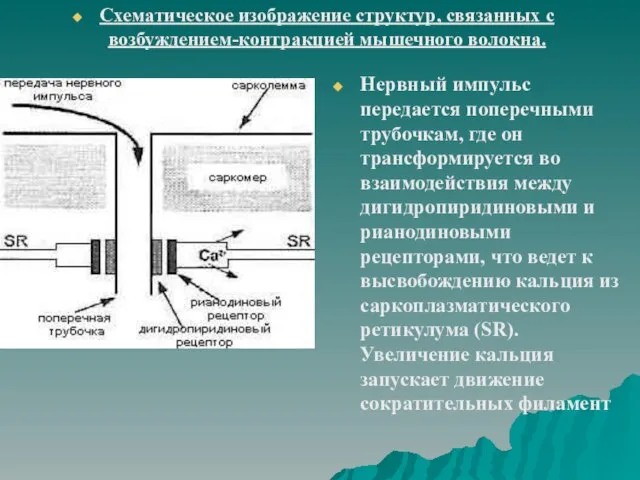
Нервный импульс передается поперечными трубочкам, где он трансформируется во взаимодействия между
Нервный импульс передается поперечными трубочкам, где он трансформируется во взаимодействия между

Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в
Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в

При одиночном мышечном сокращении различают 3 фазы:
латентный период продолжительностью
При одиночном мышечном сокращении различают 3 фазы:
латентный период продолжительностью



Одиночное сокращение и тетанус
При воздействии импульсов, наносимых с частотой более 20
Одиночное сокращение и тетанус
При воздействии импульсов, наносимых с частотой более 20

Если период следования импульсов больше продолжительности одиночного сокращения (100 мс), то
Если период следования импульсов больше продолжительности одиночного сокращения (100 мс), то

Used by permission of John W. Kimball
Н.Е.Введенский проводя опыты на
Used by permission of John W. Kimball
Н.Е.Введенский проводя опыты на

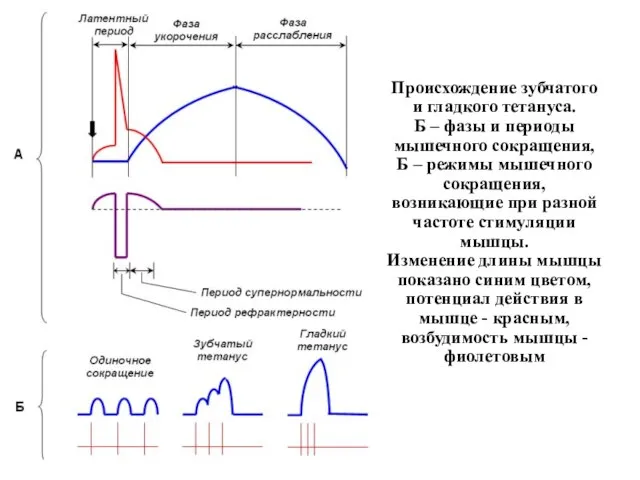
Происхождение зубчатого и гладкого тетануса.
Б – фазы и периоды мышечного сокращения,
Б
Происхождение зубчатого и гладкого тетануса. Б – фазы и периоды мышечного сокращения, Б
 Презентация Живые организмы
Презентация Живые организмы Презентация Космический мусор
Презентация Космический мусор Высшие споровые растения. Мхи
Высшие споровые растения. Мхи Царство – растения Отдел – цветковые (покрытосеменные) Класс – двудольные Семейство – Крестоцветные.
Царство – растения Отдел – цветковые (покрытосеменные) Класс – двудольные Семейство – Крестоцветные. ЦИТОЛОГИЯ Наука о клетке История открытия клетки Строение клеток
ЦИТОЛОГИЯ Наука о клетке История открытия клетки Строение клеток Творчество Виктора Петровича Астафьева
Творчество Виктора Петровича Астафьева Презентация на тему "Простейшие (Корненожки, Радиолярии, Солнечники, Споровики)" - скачать презентации по Биологии
Презентация на тему "Простейшие (Корненожки, Радиолярии, Солнечники, Споровики)" - скачать презентации по Биологии КРОВЬ ЧЕЛОВЕКА И ЛЯГУШКИ, СТРОЕНИЕ КРОВЕТВОРНЫХ ОРГАНОВ, ГЕМОПОЭЗ
КРОВЬ ЧЕЛОВЕКА И ЛЯГУШКИ, СТРОЕНИЕ КРОВЕТВОРНЫХ ОРГАНОВ, ГЕМОПОЭЗ  Морфология побега высших растений. Побеговые системы. (Лекция 7)
Морфология побега высших растений. Побеговые системы. (Лекция 7) Презентация на тему Зелёная аптека
Презентация на тему Зелёная аптека Вітамин С
Вітамин С Митохондрии, пероксисомы и пластиды
Митохондрии, пероксисомы и пластиды Скелет человека
Скелет человека Лёгкие — органы воздушного дыхания у человека
Лёгкие — органы воздушного дыхания у человека Основы анатомии и физиологии сельскохозяйствен ных животных. Тема 2
Основы анатомии и физиологии сельскохозяйствен ных животных. Тема 2 Польові дослідження
Польові дослідження Презентация на тему "Архейская эра" - скачать презентации по Биологии
Презентация на тему "Архейская эра" - скачать презентации по Биологии Какая у нас память Исследовательская работа
Какая у нас память Исследовательская работа  Презентация на тему "Каменный лес" - скачать презентации по Биологии
Презентация на тему "Каменный лес" - скачать презентации по Биологии Сенсорная система. Органы чувств
Сенсорная система. Органы чувств Презентация на тему "звезды" - скачать бесплатно презентации по Биологии
Презентация на тему "звезды" - скачать бесплатно презентации по Биологии Презентация на тему "Влияние алкоголя на беременную женщину" - скачать бесплатно презентации по Биологии
Презентация на тему "Влияние алкоголя на беременную женщину" - скачать бесплатно презентации по Биологии Презентация на тему "Животные, о существовании которых вы может даже и не знали" - скачать бесплатно презентации по Биологии
Презентация на тему "Животные, о существовании которых вы может даже и не знали" - скачать бесплатно презентации по Биологии Влияние света на интенсивность дыхания
Влияние света на интенсивность дыхания Конкуренция белок!
Конкуренция белок! Селекция животных “Борька” – гибрид домашней козы и сибирского козерога.
Селекция животных “Борька” – гибрид домашней козы и сибирского козерога.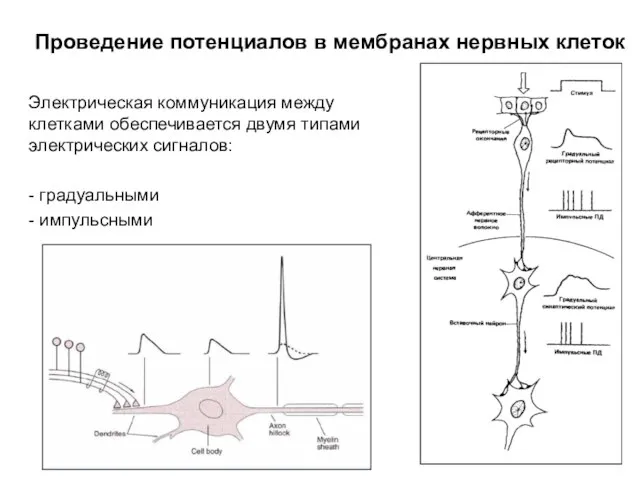 Проведение потенциалов в мембранах нервных клеток
Проведение потенциалов в мембранах нервных клеток Среды обитания организмов
Среды обитания организмов