- Сущность живого. Ядерный аппарат клетки

Содержание
- 2. План лекции 1. О сущности живого 2. Клетка – миниатюрная биосистема 3. Поверхностный аппарат клетки 4.
- 3. Сущность живого Жизнь существует в форме нуклеопротеидных комплексов. Это биосистемы характеризующиеся 5 признаками: открытые самообновляющиеся саморегулирующиеся
- 5. Активный транспорт К+-Na+ насос
- 6. Метаболический аппарат клетки Современные представления о связи мембранных органелл в клетке В клетке есть постоянные компартменты,
- 7. Метаболический аппарат клетки Изучен механизм внутриклеточного транспорта секретируемых соединений. Клатриновый эндоцитоз In vitro культура фибробластов активно
- 8. Лекция 2 Межклеточная химическая сигнализация План лекции: Понятие о биомолекулярных сигналах и рецепторах Стратегии межклеточной химической
- 9. Внеклеточные регуляторные вещества – биомолекулярные сигналы: нейромедиаторы и нейропептиды гормоны локальные химические агенты , такие как
- 10. Стратегии химической сигнализации
- 11. Основные этапы передачи сигнала
- 12. Рецепторная специфичность Каждая клетка синтезирует определенный набор рецепторов. Набор рецепторов клетки изменяется в процессе развития и
- 13. Связывание сигнальной молекулы с рецептором и активация рецептора. Механизм действия гидрофильных и липофильных сигнальных молекул Многообразие
- 14. Семейство липофильных рецепторов Лиганды: стероиды тироксин ретиноиды свободно проникают через плазмалемму в цитозоль и активируют рецепторы
- 15. Семейство рецепторов клеточной поверхности Лиганды: гидрофильные сигнальные молекулы (нейромедиаторы – ацетилхолин,норадреналин и т.п., гормоны пептидной природы
- 16. Механизмы преобразования сигнала в клетке Изменение конформации белков Изменение активности путем фосфорилирования и дефосфорилирования белков Преобразование
- 17. Лекция 3 - 4 Поток энергии. Молекулярные механизмы преобразования энергии в клетке. План лекции Понятие о
- 18. План лекции Понятие о потоке энергии. Метаболизм. Пластический и энергетический обмены. Этапы энергетического обмена. Фотосинтез. Связь
- 19. Поток энергии
- 20. Растения в процессе фотосинтеза преобразуют солнечную энергию в химическую. Гетеротрофные организмы потребляют продукты фотосинтеза и расходуют
- 21. Метаболизм – это совокупность пластического и энергетического обменов, связанных с друг другом и внешней средой. Пластический
- 22. Этапы энергетического обмена 1. Подготовительный 2. Анаэробный (бескислородный, гликолиз) 3. Аэробный (кислородный)
- 23. Подготовительный этап Протекает в пищеварительном тракте человека и животных лизосомах Крупные молекулы расщепляются на мелкие: крахмал,
- 24. Анаэробный этап Протекает в цитоплазме. С6Н12О6+2АДФ+2Фн+2НАД→2С3Н4О3+2АТФ+2НАД.Н2 глюкоза пируват (пировиноградная к-та, ПВК) Энергия, которая выделяется при расщеплении
- 25. Аэробный этап Необходим кислород. Протекает в митохондриях: в матриксе – окисление пирувата и цикл Кребса на
- 26. Строение митохондрии
- 27. Цикл Кребса (цикл лимонной кислоты)
- 29. Фотосинтез
- 30. Фотосинтез Световая фаза Преобразование энергии Солнца в химическую энергию, которая запасается в виде молекул АТФ и
- 31. Связь фотосинтеза и клеточного дыхания
- 32. Гены и геномы (5-10) План лекции 1. История изучения генов 2. Особенности генома прокариот 3. Регуляция
- 33. Особенности генома прокариот Р R Р О L 5 кодирующая последовательность 3 T Единица транскрипции участок
- 34. Особенности генома прокариот 1. Ген – экспрессируемая единица генома, включающая единицу транскрипции и регуляторные участки 2.
- 35. Особенности генома прокариот 4. У прокариот 3 вида регуляторных участков ДНК: - промоторный для связи с
- 36. Особенности генома прокариот 5. Принцип построения генома единый для всех прокариот 6. Регуляция экспрессии генов происходит,
- 37. Регуляция экспрессии генов прокариот Регуляторные участки – на расстоянии 50-75 н.п. левее сайта инициации (иногда еще
- 38. Lac – оперон
- 39. Особенности генома эукариот Р L кодирующая последовательность T Единица транскрипции участок ДНК между сайтами инициации и
- 40. Особенности генома эукариот 1. Ген – экспрессируемая единица генома, включающая единицу транскрипции и регуляторные участки 2.Гены
- 41. Особенности генома эукариот 4. Регуляторные участки обычно расположены левее сайта инициации - промоторный для связи с
- 42. Особенности генома эукариот 5. Принцип построения генома эукариот – три уровня - генный - хромосомный -
- 43. Регуляция экспрессии генов эукариот
- 44. Регуляция экспрессии генов эукариот Пример регуляция инициации транскрипции • регуляторные белки • Тепло, свет, металлы •
- 45. Регуляция экспрессии генов эукариот Регуляторные последовательности (участки) ДНК: Промоторы ≈ 100 н.п.левее сайта инициации Энхансеры от
- 46. Регуляция экспрессии генов эукариот • Механизм Регуляторные белки связываются с промотором и энхансером (активаторы и репрессоры),
- 47. План лекции 1. Международная программа «Геном человека» 2. Организация генома человека 3. Понятие о геномике и
- 48. Организация генома человека Ядро – 95% ДНК Общая протяженность ДНК ядра1,5 – 2 метра. Разделена на
- 49. Классификация генов по структуре Уники один или несколько повторов информация о структуре белков. Их мутации наиболее
- 50. Классификация генов по структуре Множественные повторы сотни тысяч, млн. копий отрезков ДНК. Теломерные и центромерные участки
- 51. Классификация генов по функции РНК – кодирующие гены Протеин – кодирующие гены Гены МТХ tРНК, rРНК,
- 52. План лекции 1. Генетический полиморфизм и разнообразие геномов человека 2. Биохимическая уникальность человека Гены предрасположенности 3.
- 53. Разнообразие геномов человека Генетическая вариабельность, ограниченная одним видом (Homo sapiens), получила название генетического полиморфизма – ГП
- 54. Гены предрасположенности Гены «внешней среды» Гены триггеры Гены рецепторов Гены предрасположенности 10%-20% всего генома
- 55. Реконструкция происхождения людей по мтДНК
- 56. Мутации и болезни Индуцированные Возникают под действием мутагенных факторов (физические, химические и биологические) Классификация точечные хромосомные
- 57. Биология развития (11-15) План лекции 1. Периоды онтогенеза человека. Пренатальное развитие 2. Метод экстракорпорального оплодотворения (ЭКО)
- 58. Критические периоды Критические периоды в развитии зародыша – это периоды наибольшей ранимости, чувствительности к действию различных
- 59. Классификация тератогенов Мутагены агенты, вызывающие мутации (ионизирующая радиация, лекарственные препараты и др.) Вирусы вирус краснухи (нем.
- 60. Классификация тератогенов Микроорганизмы бактерии – Treponema и простейшие простейшиеToxoplasma Лек.препараты * Veratrum californicum прир. хим.соед (алкалоиды)
- 61. План лекции 1. Эпигеномная изменчивость. 2. Молекулярные механизмы развития зародыша. Метилирование ДНК. 3. Закономерности развития зародыша.
- 62. Эпигеномная изменчивость. Процессы, управляющие развертыванием программы развития в клетках называют эпигенетическими (эпигеномная изменчивость)
- 63. Молекулярные механизмы развития зародыша. Метилирование ДНК. 1. Регуляция экспрессии генов основана на взаимодействии регуляторных участков ДНК
- 64. Метилирование ДНК. Метилирование цитозина – важный элемент генной активности (СН3- группы присоединяются к С в парах
- 65. Метилирование ДНК.
- 66. Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах) Два способа детерминации эмбриональных клеток Гены
- 67. Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах) мозаичный тип развития регуляционный тип развития
- 68. План лекции 1. Периоды постнатального развития. 2. «О любви не говорят, о ней все сказано…?» 3.
- 69. Проблемы старения организма. Максимальную продолжительность жизни нельзя увеличить, т.к. это признак вида (90-100 лет) Теоретически (библия)
- 70. Проблемы старения организма. Генетический контроль за продолжительностью жизни очевиден (мутация 1 гена, синдром преждевременного старения –
- 71. Теории старения (механизмы) Свободно-радикальная теория ( кол-ва свободных радикалов с возрастом) Укорачивание теломер при делении
- 72. План лекции 1. Терапевтическое клонирование 2. Вопросы трансплантации.
- 73. Терапевтическое клонирование и трансплантация Согласно современным представлениям, регенерация тканей взрослого организма и их репарация в случае
- 74. Терапевтическое клонирование и трансплантация Источники стволовых клеток: Эмбриональные стволовые клетки (бластоциста) Фетальные стволовые клетки (абортивный материал
- 75. Суть терапевтического клонирования Обычная соматическая клетка располагается около яйцеклетки, чья ядерная ДНК удалена. Под воздействием электрического
- 76. ТРАНСПЛАНТОЛОГИЯ – (греч. Trans – пере-, через; plantare – сажать, выращивать; -logia - наука) – раздел
- 77. Трансплантология Органная трансплантация. Аутотрансплантация – трансплантат того же пациента. Реплантация – пересадка органов и тканей на
- 78. Литература к лекциям I семестра 1) Мэтт Ридли «Геном»,М.Эксмо,2008 2)С.Г. Инге-Вечтомов «Генетика с основами селекции»,СПб ,
- 79. Вопросы иммунитета 2 лекции Доц. Косенкова Н.С..
- 80. 1798 — Э. Дженнер (Великобритания). Вакцинация. 1881 - Луи Пастер (Франция). Эффективная иммунизация. 1890 — Э.
- 81. Система защиты генетической целостности и качественного постоянства организма. Первая линия обороны. Физико-химические барьеры. Физиологические защитные механизмы.
- 82. КОМПОНЕНТЫ ИММУННОЙ СИСТЕМЫ Клеточные факторы (лейкоциты : лимфоциты(Т, В, NK), моноциты/макрофаги, нейтрофилы, эозинофилы и базофилы/тучные клетки).
- 83. Врожденный (Неспецифический иммунитет) Способность отличать «чужое» с помощью TLR и др. рецепторов Клеточные факторы врожденного иммунитета:
- 84. Приобретенный иммунитет (Адаптивный,Специфический ) Клеточный и гуморальный иммунный ответ Клоны Т и В лимфоцитов >1011 вариантов
- 85. T лимфоциты. Клеточный специфический иммунитет. Т-клеточные рецепторы (TCR) распознают фрагменты антигенных молекул в комплексе с МНС
- 86. В лимфоциты. Гуморальный иммунный ответ B лимфоциты распознают нерасщепленные антигены. Наивные В-лимфоциты. Плазматические B клетки –
- 87. Антитела (иммуноглобулины) Структурная единица AT — мономер из двух идентичных тяжёлых Н-цепей и двух идентичных лёгких
- 88. Основные функции антител. Эффекторные механизмы гуморального иммунитета преципитация агглютинация Нейтрализация опсонизация активация системы комплемента активация дегрануляции
- 89. 5 классов антител IgG, IgD, IgE – мономеры IgA- в секретах - димер IgM- мономер в
- 90. Моноклональные антитела. Способ создания моноклональных антител Георгом Кёлером и Цезарем Мильштейном (Нобелевская премия 1984 года). Гибридома
- 91. Геномика и биоинформатика: роль в медицине асс. каф. медицинской биологии Старунова З.И. План Введение в геномику
- 92. Влияние геномики на различные области современной медицины Биотехнология: ДНК-последовательности как инструменты диагностики, получение терапевтических белков, вакцин,
- 93. Получение терапевтических белков С помощью клонирующих векторов можно также экспрессировать клонированный ген и получать рекомбинантный белок
- 94. Главные направления научных исследований, связанных с проектом «Геном человека»: аннотирование (описание) генома – получение полезной биологической
- 95. Анализ последовательностей (генов и геномов) различных организмов показал значительную структурную и функциональную консервативность между генами и
- 97. Скачать презентацию

План лекции
1. О сущности живого
2. Клетка – миниатюрная биосистема
3. Поверхностный аппарат
План лекции
1. О сущности живого
2. Клетка – миниатюрная биосистема
3. Поверхностный аппарат

Сущность живого
Жизнь существует в форме нуклеопротеидных комплексов.
Это биосистемы характеризующиеся 5 признаками:
Сущность живого
Жизнь существует в форме нуклеопротеидных комплексов.
Это биосистемы характеризующиеся 5 признаками:
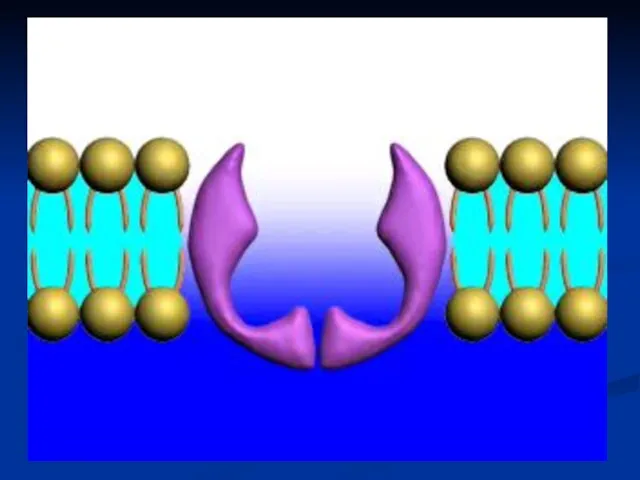
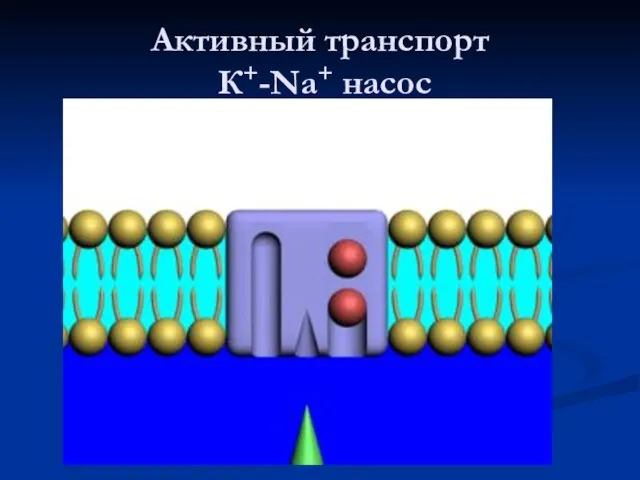
Активный транспорт
К+-Na+ насос
Активный транспорт
К+-Na+ насос

Метаболический аппарат клетки
Современные представления о связи мембранных органелл в клетке
В
Метаболический аппарат клетки
Современные представления о связи мембранных органелл в клетке
В

Метаболический аппарат клетки
Изучен механизм внутриклеточного транспорта секретируемых соединений.
Клатриновый эндоцитоз
Метаболический аппарат клетки
Изучен механизм внутриклеточного транспорта секретируемых соединений.
Клатриновый эндоцитоз

Лекция 2
Межклеточная химическая сигнализация
План лекции:
Понятие о биомолекулярных сигналах и рецепторах
Стратегии
Лекция 2
Межклеточная химическая сигнализация
План лекции:
Понятие о биомолекулярных сигналах и рецепторах
Стратегии

Внеклеточные регуляторные вещества – биомолекулярные сигналы:
нейромедиаторы и нейропептиды
гормоны
локальные химические агенты ,
Внеклеточные регуляторные вещества – биомолекулярные сигналы:
нейромедиаторы и нейропептиды
гормоны
локальные химические агенты ,
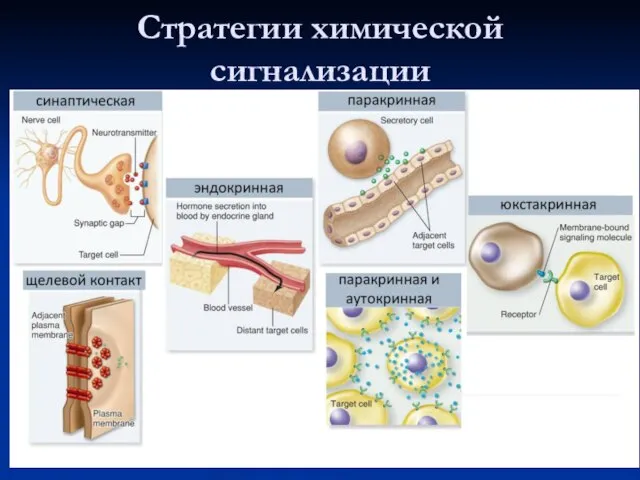
Стратегии химической сигнализации
Стратегии химической сигнализации
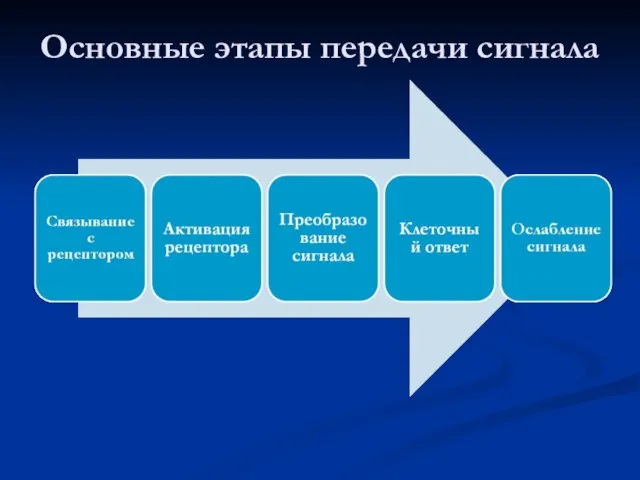
Основные этапы передачи сигнала
Основные этапы передачи сигнала

Рецепторная специфичность
Каждая клетка синтезирует определенный набор рецепторов.
Набор рецепторов клетки изменяется в
Рецепторная специфичность
Каждая клетка синтезирует определенный набор рецепторов.
Набор рецепторов клетки изменяется в
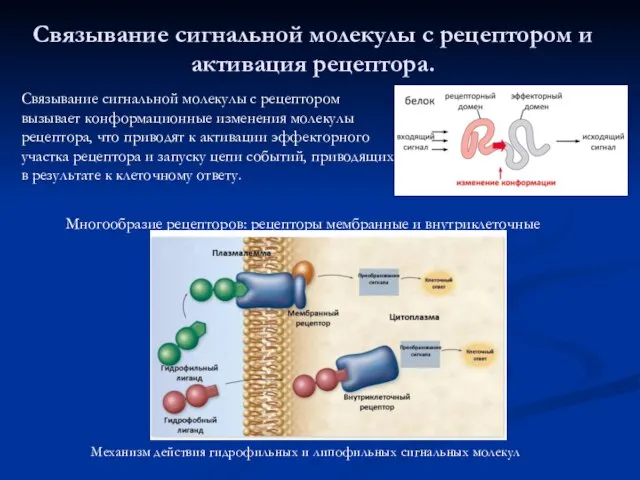
Связывание сигнальной молекулы с рецептором и активация рецептора.
Механизм действия гидрофильных и
Связывание сигнальной молекулы с рецептором и активация рецептора.
Механизм действия гидрофильных и

Семейство липофильных рецепторов
Лиганды:
стероиды
тироксин
ретиноиды
свободно проникают через плазмалемму в цитозоль и
Семейство липофильных рецепторов
Лиганды:
стероиды
тироксин
ретиноиды
свободно проникают через плазмалемму в цитозоль и

Семейство рецепторов клеточной поверхности
Лиганды:
гидрофильные сигнальные молекулы (нейромедиаторы – ацетилхолин,норадреналин и т.п.,
Семейство рецепторов клеточной поверхности
Лиганды:
гидрофильные сигнальные молекулы (нейромедиаторы – ацетилхолин,норадреналин и т.п.,

Механизмы преобразования сигнала в клетке
Изменение конформации белков
Изменение активности путем фосфорилирования и
Механизмы преобразования сигнала в клетке
Изменение конформации белков
Изменение активности путем фосфорилирования и

Лекция 3 - 4
Поток энергии. Молекулярные механизмы преобразования энергии в клетке.
План
Лекция 3 - 4 Поток энергии. Молекулярные механизмы преобразования энергии в клетке. План

План лекции
Понятие о потоке энергии.
Метаболизм. Пластический и энергетический обмены.
Этапы энергетического обмена.
Фотосинтез.
План лекции
Понятие о потоке энергии.
Метаболизм. Пластический и энергетический обмены.
Этапы энергетического обмена.
Фотосинтез.
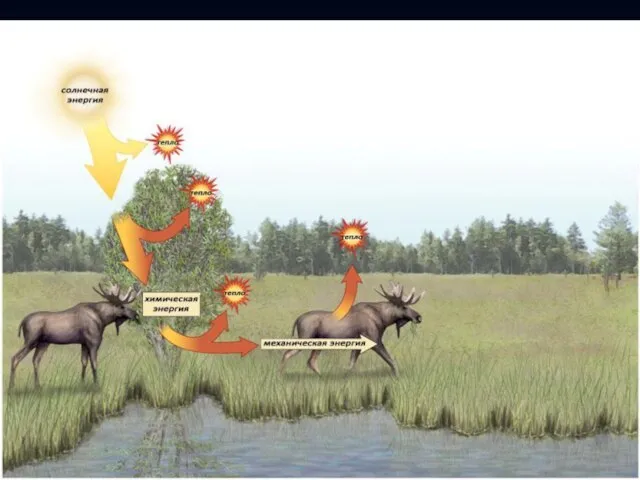
Поток энергии
Поток энергии

Растения в процессе фотосинтеза преобразуют солнечную энергию в химическую.
Гетеротрофные организмы потребляют
Растения в процессе фотосинтеза преобразуют солнечную энергию в химическую.
Гетеротрофные организмы потребляют

Метаболизм – это совокупность пластического и энергетического обменов, связанных с друг
Метаболизм – это совокупность пластического и энергетического обменов, связанных с друг

Этапы энергетического обмена
1. Подготовительный
2. Анаэробный (бескислородный, гликолиз)
3. Аэробный (кислородный)
Этапы энергетического обмена
1. Подготовительный
2. Анаэробный (бескислородный, гликолиз)
3. Аэробный (кислородный)

Подготовительный этап
Протекает в
пищеварительном тракте человека и животных
лизосомах
Крупные молекулы расщепляются на
Подготовительный этап
Протекает в
пищеварительном тракте человека и животных
лизосомах
Крупные молекулы расщепляются на

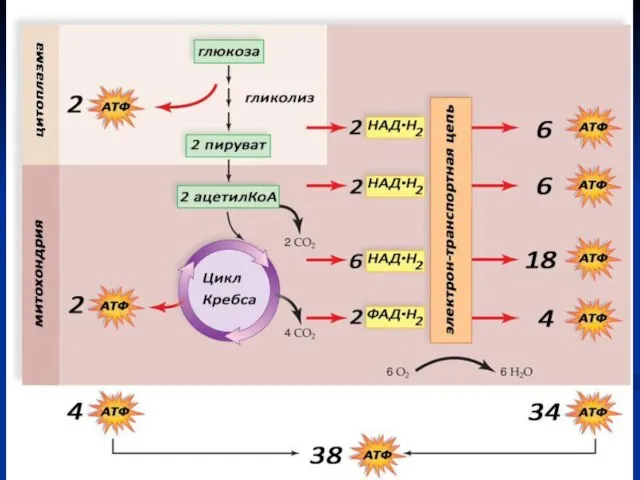
Анаэробный этап
Протекает в цитоплазме.
С6Н12О6+2АДФ+2Фн+2НАД→2С3Н4О3+2АТФ+2НАД.Н2
глюкоза
пируват
(пировиноградная
к-та, ПВК)
Энергия, которая выделяется при расщеплении молекулы
Анаэробный этап
Протекает в цитоплазме.
С6Н12О6+2АДФ+2Фн+2НАД→2С3Н4О3+2АТФ+2НАД.Н2
глюкоза
пируват
(пировиноградная
к-та, ПВК)
Энергия, которая выделяется при расщеплении молекулы

Аэробный этап
Необходим кислород.
Протекает в митохондриях:
в матриксе – окисление пирувата и
Аэробный этап
Необходим кислород.
Протекает в митохондриях:
в матриксе – окисление пирувата и
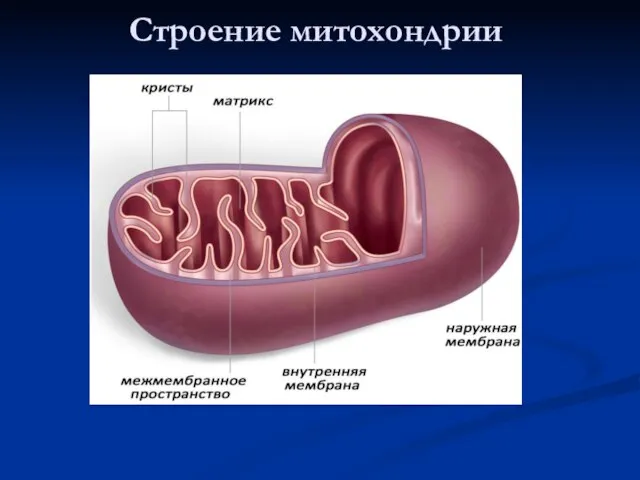
Строение митохондрии
Строение митохондрии
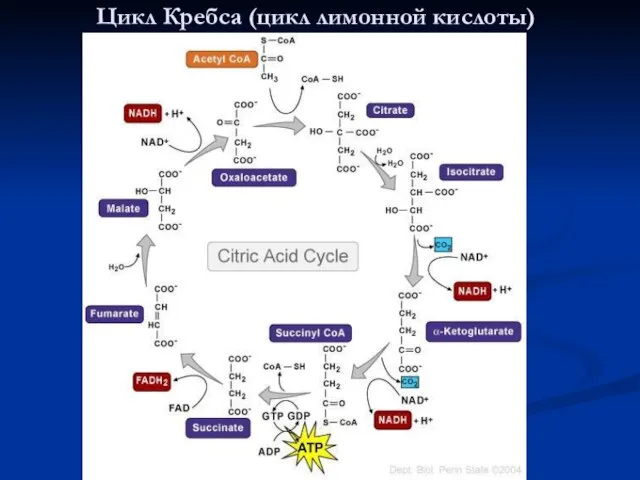
Цикл Кребса (цикл лимонной кислоты)
Цикл Кребса (цикл лимонной кислоты)
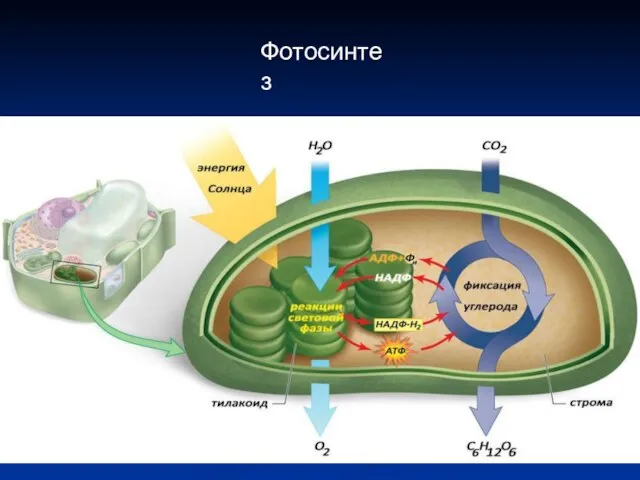
Фотосинтез
Фотосинтез

Фотосинтез
Световая фаза
Преобразование энергии Солнца в химическую энергию, которая запасается в виде
Фотосинтез
Световая фаза Преобразование энергии Солнца в химическую энергию, которая запасается в виде
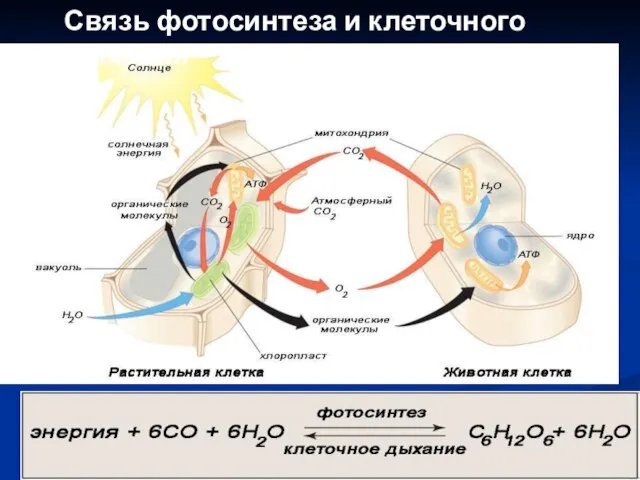
Связь фотосинтеза и клеточного дыхания
Связь фотосинтеза и клеточного дыхания

Гены и геномы (5-10)
План лекции
1. История изучения генов
2. Особенности генома
Гены и геномы (5-10)
План лекции
1. История изучения генов
2. Особенности генома
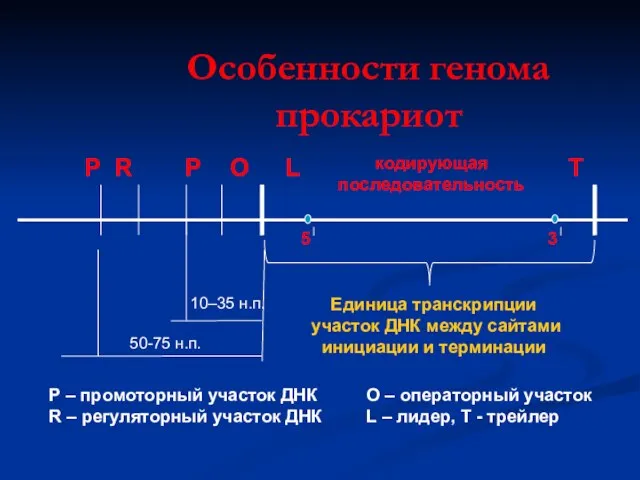
Особенности генома прокариот
Р
R
Р
О
L
5
кодирующая
последовательность
3
T
Единица транскрипции
участок ДНК между сайтами
инициации и
Особенности генома прокариот
Р
R
Р
О
L
5
кодирующая
последовательность
3
T
Единица транскрипции участок ДНК между сайтами инициации и

Особенности генома прокариот
1. Ген – экспрессируемая единица генома, включающая единицу транскрипции
Особенности генома прокариот
1. Ген – экспрессируемая единица генома, включающая единицу транскрипции

Особенности генома прокариот
4. У прокариот 3 вида регуляторных участков ДНК:
- промоторный
Особенности генома прокариот
4. У прокариот 3 вида регуляторных участков ДНК:
- промоторный

Особенности генома прокариот
5. Принцип построения генома единый для всех прокариот
6. Регуляция
Особенности генома прокариот
5. Принцип построения генома единый для всех прокариот
6. Регуляция

Регуляция экспрессии генов прокариот
Регуляторные участки – на расстоянии 50-75 н.п.
Регуляция экспрессии генов прокариот
Регуляторные участки – на расстоянии 50-75 н.п.
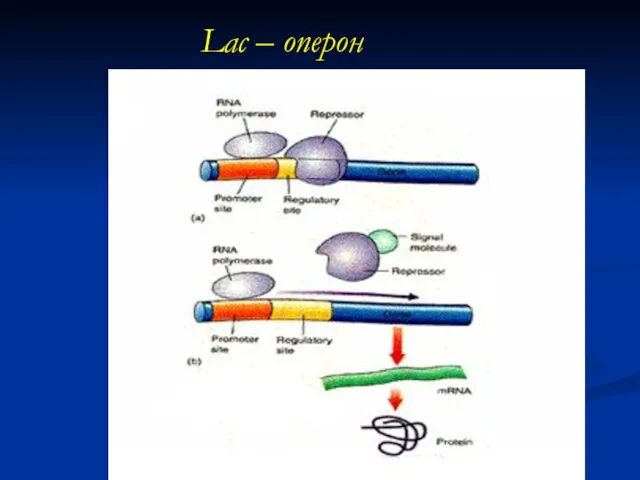
Lac – оперон
Lac – оперон
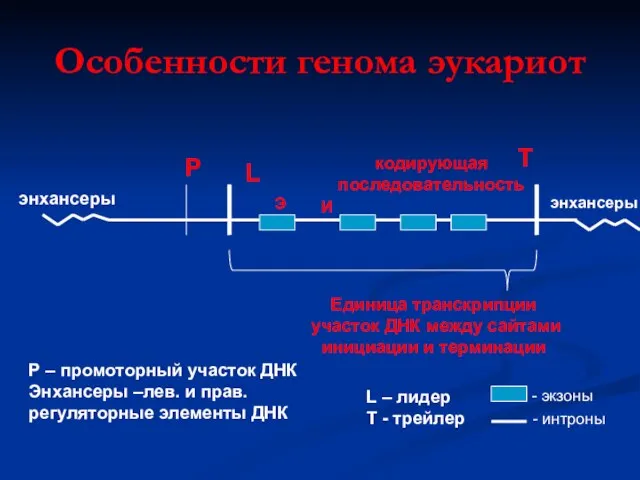
Особенности генома эукариот
Р
L
кодирующая
последовательность
T
Единица транскрипции
участок ДНК между сайтами
инициации и
Особенности генома эукариот
Р
L
кодирующая
последовательность
T
Единица транскрипции участок ДНК между сайтами инициации и

Особенности генома эукариот
1. Ген – экспрессируемая единица генома, включающая единицу транскрипции
Особенности генома эукариот
1. Ген – экспрессируемая единица генома, включающая единицу транскрипции

Особенности генома эукариот
4. Регуляторные участки обычно расположены левее сайта инициации
- промоторный
Особенности генома эукариот
4. Регуляторные участки обычно расположены левее сайта инициации - промоторный

Особенности генома эукариот
5. Принцип построения генома эукариот – три уровня
- генный
-
Особенности генома эукариот
5. Принцип построения генома эукариот – три уровня
- генный
-
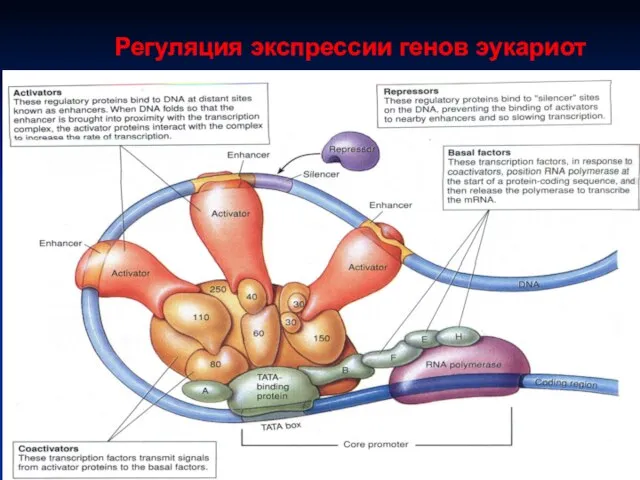
Регуляция экспрессии генов эукариот
Регуляция экспрессии генов эукариот

Регуляция экспрессии генов эукариот
Пример регуляция инициации транскрипции
• регуляторные белки
•
Регуляция экспрессии генов эукариот
Пример регуляция инициации транскрипции
• регуляторные белки
•

Регуляция экспрессии генов эукариот
Регуляторные последовательности (участки) ДНК:
Промоторы ≈ 100 н.п.левее сайта
Регуляция экспрессии генов эукариот
Регуляторные последовательности (участки) ДНК:
Промоторы ≈ 100 н.п.левее сайта

Регуляция экспрессии генов эукариот
• Механизм
Регуляторные белки связываются с промотором и энхансером
Регуляция экспрессии генов эукариот
• Механизм
Регуляторные белки связываются с промотором и энхансером

План лекции
1. Международная программа
«Геном человека»
2. Организация генома человека
3.
План лекции
1. Международная программа
«Геном человека»
2. Организация генома человека
3.

Организация генома человека
Ядро – 95% ДНК Общая протяженность ДНК ядра1,5 –
Организация генома человека
Ядро – 95% ДНК Общая протяженность ДНК ядра1,5 –

Классификация генов
по структуре
Уники один или несколько повторов
информация о структуре белков.
Классификация генов
по структуре
Уники один или несколько повторов информация о структуре белков.

Классификация генов
по структуре
Множественные повторы сотни тысяч, млн. копий
отрезков ДНК. Теломерные
Классификация генов
по структуре
Множественные повторы сотни тысяч, млн. копий
отрезков ДНК. Теломерные
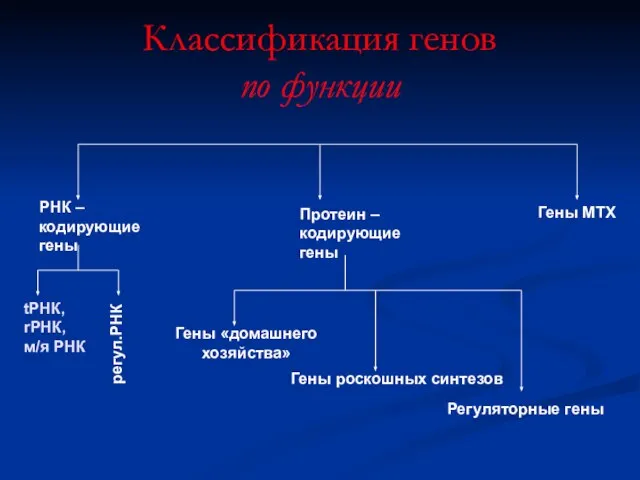
Классификация генов
по функции
РНК – кодирующие гены
Протеин –
кодирующие
гены
Гены МТХ
tРНК,
rРНК,
м/я РНК
регул.РНК
Гены
Классификация генов
по функции
РНК – кодирующие гены
Протеин –
кодирующие
гены
Гены МТХ
tРНК,
rРНК,
м/я РНК
регул.РНК
Гены

План лекции
1. Генетический полиморфизм и
разнообразие геномов человека
2. Биохимическая уникальность человека
План лекции
1. Генетический полиморфизм и
разнообразие геномов человека
2. Биохимическая уникальность человека

Разнообразие геномов человека
Генетическая вариабельность, ограниченная одним видом (Homo sapiens), получила название
Разнообразие геномов человека
Генетическая вариабельность, ограниченная одним видом (Homo sapiens), получила название

Гены предрасположенности
Гены «внешней среды»
Гены триггеры
Гены рецепторов
Гены предрасположенности
10%-20% всего генома
Гены предрасположенности
Гены «внешней среды»
Гены триггеры
Гены рецепторов
Гены предрасположенности
10%-20% всего генома
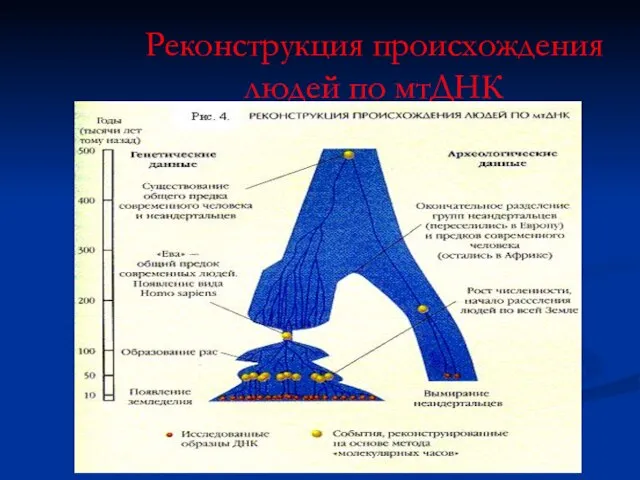
Реконструкция происхождения людей по мтДНК
Реконструкция происхождения людей по мтДНК

Мутации и болезни
Индуцированные
Возникают под действием мутагенных факторов (физические, химические и биологические)
Классификация
точечные
хромосомные
динамические
геномные
Мутации и болезни
Индуцированные
Возникают под действием мутагенных факторов (физические, химические и биологические)
Классификация
точечные
хромосомные
динамические
геномные

Биология развития (11-15)
План лекции
1. Периоды онтогенеза человека.
Пренатальное развитие
2. Метод экстракорпорального
Биология развития (11-15)
План лекции
1. Периоды онтогенеза человека.
Пренатальное развитие
2. Метод экстракорпорального

Критические периоды
Критические периоды в развитии зародыша – это периоды наибольшей ранимости,
Критические периоды
Критические периоды в развитии зародыша – это периоды наибольшей ранимости,

Классификация тератогенов
Мутагены агенты, вызывающие
мутации (ионизирующая
радиация, лекарственные
препараты и др.)
Вирусы вирус краснухи (нем.
Классификация тератогенов
Мутагены агенты, вызывающие
мутации (ионизирующая
радиация, лекарственные
препараты и др.)
Вирусы вирус краснухи (нем.

Классификация тератогенов
Микроорганизмы бактерии – Treponema
и простейшие простейшиеToxoplasma
Лек.препараты * Veratrum californicum
прир.
Классификация тератогенов
Микроорганизмы бактерии – Treponema
и простейшие простейшиеToxoplasma
Лек.препараты * Veratrum californicum
прир.

План лекции
1. Эпигеномная изменчивость.
2. Молекулярные механизмы развития зародыша. Метилирование ДНК.
План лекции
1. Эпигеномная изменчивость.
2. Молекулярные механизмы развития зародыша. Метилирование ДНК.
Эпигеномная изменчивость.
Процессы, управляющие развертыванием программы развития в клетках называют эпигенетическими (эпигеномная
Эпигеномная изменчивость.
Процессы, управляющие развертыванием программы развития в клетках называют эпигенетическими (эпигеномная

Молекулярные механизмы развития зародыша. Метилирование ДНК.
1. Регуляция экспрессии генов основана на
Молекулярные механизмы развития зародыша. Метилирование ДНК.
1. Регуляция экспрессии генов основана на

Метилирование ДНК.
Метилирование цитозина – важный элемент генной активности (СН3- группы присоединяются
Метилирование ДНК.
Метилирование цитозина – важный элемент генной активности (СН3- группы присоединяются
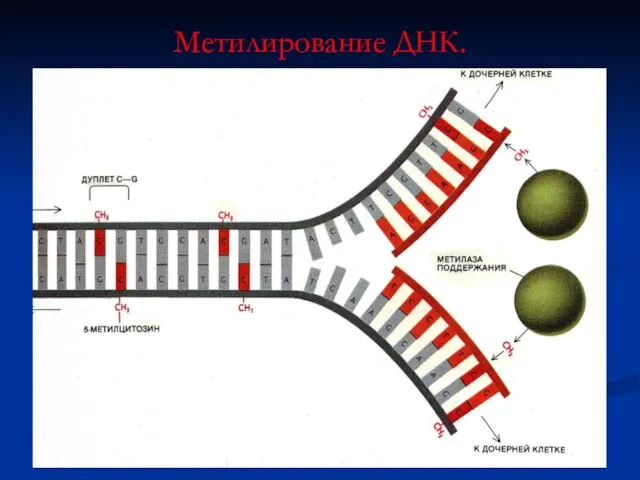
Метилирование ДНК.
Метилирование ДНК.

Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах)
Два способа
Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах)
Два способа
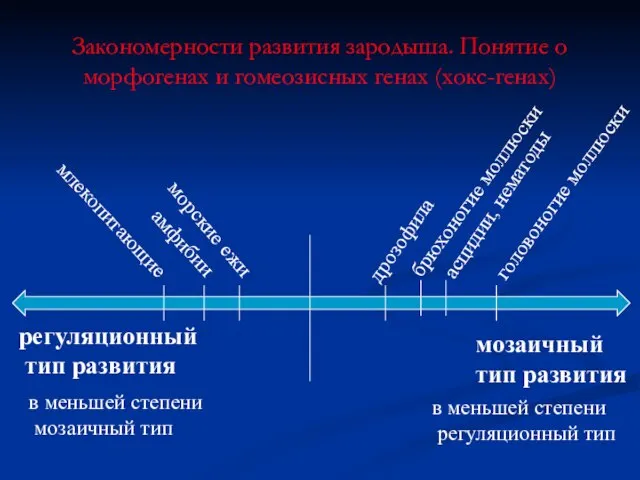
Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах)
мозаичный
тип развития
регуляционный
Закономерности развития зародыша. Понятие о морфогенах и гомеозисных генах (хокс-генах)
мозаичный
тип развития
регуляционный

План лекции
1. Периоды постнатального развития.
2. «О любви не говорят, о
План лекции
1. Периоды постнатального развития.
2. «О любви не говорят, о

Проблемы старения организма.
Максимальную продолжительность жизни нельзя увеличить, т.к. это признак вида
Проблемы старения организма.
Максимальную продолжительность жизни нельзя увеличить, т.к. это признак вида

Проблемы старения организма.
Генетический контроль за продолжительностью жизни очевиден (мутация 1 гена,
Проблемы старения организма.
Генетический контроль за продолжительностью жизни очевиден (мутация 1 гена,

Теории старения (механизмы)
Свободно-радикальная теория ( кол-ва свободных радикалов с возрастом)
Укорачивание
Теории старения (механизмы)
Свободно-радикальная теория ( кол-ва свободных радикалов с возрастом)
Укорачивание

План лекции
1. Терапевтическое клонирование
2. Вопросы трансплантации.
План лекции
1. Терапевтическое клонирование
2. Вопросы трансплантации.
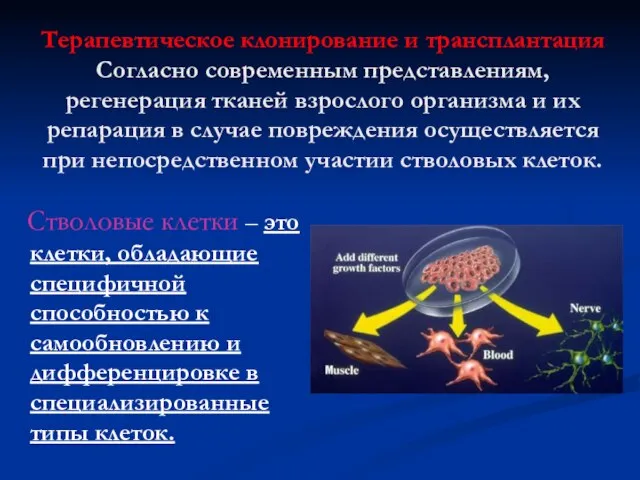
Терапевтическое клонирование и трансплантация
Согласно современным представлениям, регенерация тканей взрослого организма
Терапевтическое клонирование и трансплантация Согласно современным представлениям, регенерация тканей взрослого организма

Терапевтическое клонирование и трансплантация
Источники стволовых клеток:
Эмбриональные стволовые клетки (бластоциста)
Фетальные стволовые
Терапевтическое клонирование и трансплантация
Источники стволовых клеток:
Эмбриональные стволовые клетки (бластоциста)
Фетальные стволовые
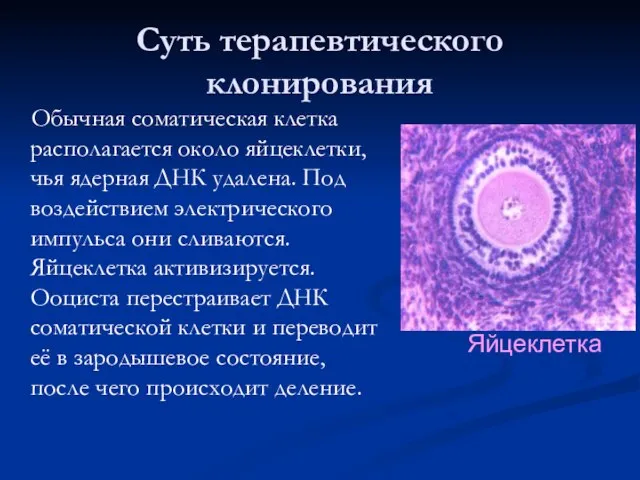
Суть терапевтического клонирования
Обычная соматическая клетка располагается около яйцеклетки, чья ядерная
Суть терапевтического клонирования
Обычная соматическая клетка располагается около яйцеклетки, чья ядерная

ТРАНСПЛАНТОЛОГИЯ – (греч. Trans – пере-, через; plantare – сажать, выращивать;
ТРАНСПЛАНТОЛОГИЯ – (греч. Trans – пере-, через; plantare – сажать, выращивать;

Трансплантология
Органная трансплантация.
Аутотрансплантация – трансплантат того же пациента.
Реплантация – пересадка органов
Трансплантология
Органная трансплантация.
Аутотрансплантация – трансплантат того же пациента.
Реплантация – пересадка органов

Литература к лекциям I семестра
1) Мэтт Ридли «Геном»,М.Эксмо,2008
2)С.Г. Инге-Вечтомов «Генетика
Литература к лекциям I семестра
1) Мэтт Ридли «Геном»,М.Эксмо,2008
2)С.Г. Инге-Вечтомов «Генетика

Вопросы иммунитета
2 лекции
Доц. Косенкова Н.С..
Вопросы иммунитета
2 лекции
Доц. Косенкова Н.С..

1798 — Э. Дженнер (Великобритания). Вакцинация.
1881 - Луи Пастер (Франция).
1798 — Э. Дженнер (Великобритания). Вакцинация.
1881 - Луи Пастер (Франция).

Система защиты генетической целостности и качественного постоянства организма.
Первая линия обороны.
Физико-химические барьеры.
Система защиты генетической целостности и качественного постоянства организма.
Первая линия обороны.
Физико-химические барьеры.
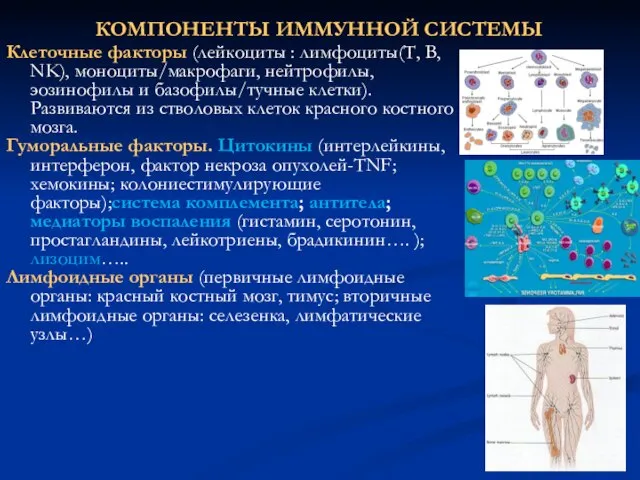
КОМПОНЕНТЫ ИММУННОЙ СИСТЕМЫ
Клеточные факторы (лейкоциты : лимфоциты(Т, В, NK), моноциты/макрофаги, нейтрофилы,
КОМПОНЕНТЫ ИММУННОЙ СИСТЕМЫ
Клеточные факторы (лейкоциты : лимфоциты(Т, В, NK), моноциты/макрофаги, нейтрофилы,

Врожденный (Неспецифический иммунитет)
Способность отличать «чужое» с помощью TLR и др. рецепторов
Клеточные
Врожденный (Неспецифический иммунитет)
Способность отличать «чужое» с помощью TLR и др. рецепторов
Клеточные
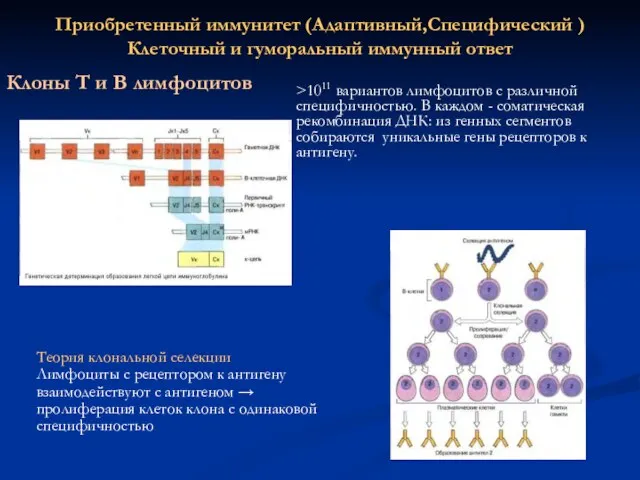
Приобретенный иммунитет (Адаптивный,Специфический )
Клеточный и гуморальный иммунный ответ
Клоны Т и В
Приобретенный иммунитет (Адаптивный,Специфический )
Клеточный и гуморальный иммунный ответ
Клоны Т и В

T лимфоциты. Клеточный специфический иммунитет.
Т-клеточные рецепторы (TCR) распознают фрагменты антигенных молекул
T лимфоциты. Клеточный специфический иммунитет.
Т-клеточные рецепторы (TCR) распознают фрагменты антигенных молекул

В лимфоциты. Гуморальный иммунный ответ
B лимфоциты распознают нерасщепленные антигены.
Наивные
В лимфоциты. Гуморальный иммунный ответ
B лимфоциты распознают нерасщепленные антигены.
Наивные

Антитела (иммуноглобулины)
Структурная единица AT — мономер
из двух идентичных тяжёлых Н-цепей
Антитела (иммуноглобулины)
Структурная единица AT — мономер
из двух идентичных тяжёлых Н-цепей

Основные функции антител. Эффекторные механизмы гуморального иммунитета
преципитация
агглютинация
Нейтрализация
опсонизация
активация системы комплемента
активация дегрануляции
Основные функции антител. Эффекторные механизмы гуморального иммунитета
преципитация
агглютинация
Нейтрализация
опсонизация
активация системы комплемента
активация дегрануляции
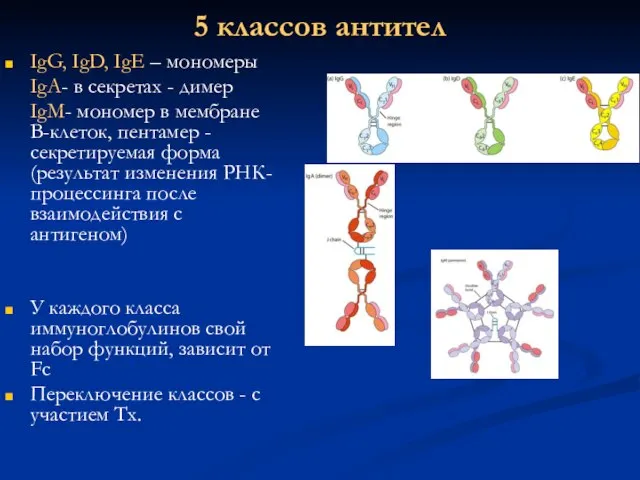
5 классов антител
IgG, IgD, IgE – мономеры
IgA- в секретах - димер
IgM-
5 классов антител
IgG, IgD, IgE – мономеры
IgA- в секретах - димер
IgM-

Моноклональные антитела.
Способ создания моноклональных антител Георгом Кёлером и Цезарем Мильштейном (Нобелевская
Моноклональные антитела.
Способ создания моноклональных антител Георгом Кёлером и Цезарем Мильштейном (Нобелевская

Геномика и
биоинформатика: роль в медицине
асс. каф. медицинской биологии Старунова
Геномика и
биоинформатика: роль в медицине
асс. каф. медицинской биологии Старунова

Влияние геномики на различные области современной медицины
Биотехнология: ДНК-последовательности как инструменты диагностики,
Влияние геномики на различные области современной медицины
Биотехнология: ДНК-последовательности как инструменты диагностики,

Получение терапевтических белков
С помощью клонирующих векторов можно также
экспрессировать клонированный ген
Получение терапевтических белков
С помощью клонирующих векторов можно также
экспрессировать клонированный ген

Главные направления научных
исследований, связанных
с проектом «Геном человека»:
аннотирование (описание) генома
Главные направления научных
исследований, связанных
с проектом «Геном человека»:
аннотирование (описание) генома
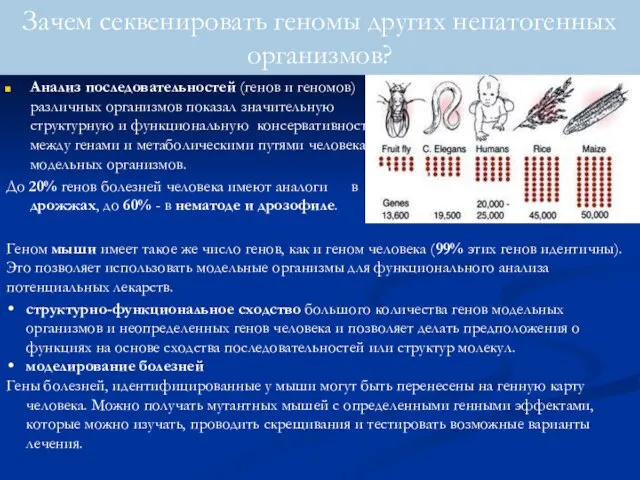
Анализ последовательностей (генов и геномов) различных организмов показал значительную структурную и
Анализ последовательностей (генов и геномов) различных организмов показал значительную структурную и
 Введение в биологию
Введение в биологию Тип Mollusca. Строение моллюсков классов Gastropoda и Cephalopoda
Тип Mollusca. Строение моллюсков классов Gastropoda и Cephalopoda Животные Африки
Животные Африки Плесневые грибы и дрожжи Питание Размножение Значение в природе
Плесневые грибы и дрожжи Питание Размножение Значение в природе  Панголиные. Классификация
Панголиные. Классификация Мухи и меры борьбы с ними
Мухи и меры борьбы с ними Презентация на тему "Хвостатые" - скачать презентации по Биологии
Презентация на тему "Хвостатые" - скачать презентации по Биологии I закон Менделя Закон доминирования: «При скрещивании двух гомозиготных организмов, отличающихся по альтернативным вариантам
I закон Менделя Закон доминирования: «При скрещивании двух гомозиготных организмов, отличающихся по альтернативным вариантам РЕФЛЕКСИВНО – ДЕЛОВАЯ ИГРА РЕФЛЕКСИВНО – ДЕЛОВАЯ ИГРА ПО БИОЛОГИИ «ЦИТОЛОГИЯ. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ» Подго
РЕФЛЕКСИВНО – ДЕЛОВАЯ ИГРА РЕФЛЕКСИВНО – ДЕЛОВАЯ ИГРА ПО БИОЛОГИИ «ЦИТОЛОГИЯ. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ» Подго Понятие биогеоценоза
Понятие биогеоценоза Тип членистоногие. Классы ракообразных, паукообразных, насекомых
Тип членистоногие. Классы ракообразных, паукообразных, насекомых Метаболизм углеводов
Метаболизм углеводов БИОЛОГИЯ 7 КЛАСС раздел «Животные» темы «Птицы» и «Млекопитающие» - по 5 уроков Разработка 10 уроков с использованием компьютера Л
БИОЛОГИЯ 7 КЛАСС раздел «Животные» темы «Птицы» и «Млекопитающие» - по 5 уроков Разработка 10 уроков с использованием компьютера Л Возникновение жизни на Земле
Возникновение жизни на Земле Класс Аскомицеты или сумчатые грибы
Класс Аскомицеты или сумчатые грибы Віруси, пріони
Віруси, пріони Животный мир. Антарктида
Животный мир. Антарктида Родина Складноцвіті
Родина Складноцвіті Биогенные d-элементы
Биогенные d-элементы Пластический обмен. Синтез белка
Пластический обмен. Синтез белка :Балықтар дүниесінің ерекшеліктерін, маңызын, қызметін, құрылысын түсіндіру
:Балықтар дүниесінің ерекшеліктерін, маңызын, қызметін, құрылысын түсіндіру Пурины. Строение пурина. (Лекция 9)
Пурины. Строение пурина. (Лекция 9) СВЕТ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
СВЕТ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР  Клітина – елементарна одиниця живого. Біомембрани
Клітина – елементарна одиниця живого. Біомембрани Пищеварительная система человека
Пищеварительная система человека Концепции появления живого на земле. (Лекция 10)
Концепции появления живого на земле. (Лекция 10) Мышцы туловища
Мышцы туловища Введение в курс спортивной физиологии. Физиологическая характеристика различных видов мышечной деятельности
Введение в курс спортивной физиологии. Физиологическая характеристика различных видов мышечной деятельности