-
Клеточный цикл и апоптоз

Содержание
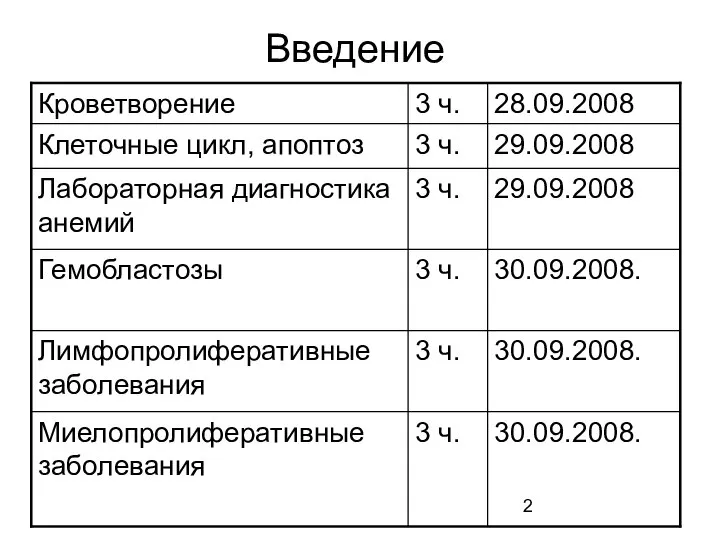
- 2. Введение
- 3. I Прибайкальская межрегиональная научно-практическая конференция (2006 год)
- 4. Кроветворение Кроветворение (синоним гематопоэз) — процесс, заключающийся в серии клеточных дифференцировок, в результате которых образуются зрелые
- 5. Сигнальные системы клетки
- 6. Биология сигнальных систем клетки Кроветворные клетки имеют рецепторы к экзогенным сигнальным молекулам: факторам роста, цитокинам, гормонам(стероидным
- 7. Киназы и фосфатазы Фосфорилирование и дефосфорилирование - основные механизмы внутриклеточной передачи сигнала В этих профессах участвуют
- 8. Рецепторы факторов роста Рецепторы факторов роста обычно стимулируют клеточный рост, однако при определенных условиях они могут
- 9. Вторичные мессенджеры Циклический аденозинмонофосфат ( cAMP) протеинкиназа А Циклический аденозинмонофосфат (cGMP) Диацилглицерол ( DA) связывается с
- 10. Способы доставки сигнальных молекул к клеткам Эндокринный механизм Паракринный механизм – клетка секретирует сигнальные молекулы и
- 11. Рецепторы клеток Группа 1 Семейство липофильных рецепторов: Стероиды: ГК, МК, половые стероиды Тиреоидные гормоны, тироксин Ретиноиды,
- 12. Fabio Candotti, Luigi Notarangelo, 2002 Экспрессия сигнальных молекул Т и В лимфоцитов
- 13. Fluorescence Distribution – NF-kB Translocation Assay
- 14. Связывание с bcr/abl-тирозинкиназой: сравнение иматиниба и нилотиниба
- 15. Внутриклеточные сигнальные пути Кроме активации ras-путиКроме активации ras-пути , в клетках существуют еще по крайней мере
- 16. Свойства факторов роста Факторами роста называют группу белковых молекул, индуцирующих синтез ДНКФакторами роста называют группу белковых
- 17. К Л Е Т О Ч Ы Й Ц И К Л Основные термины: Интерфаза Митотический
- 18. Трубочки, колечки и висюльки Первым, как отмечают историки, клетку увидел Роберт Гук в 1663 году. Но
- 19. Митотический цикл Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению,
- 20. Жизненный цикл клетки Профаза Метафаза Анафаза Телофаза Деление клетки Интерфаза
- 22. Митотический цикл Митотический цикл = фаза М – это цикл деления, который длится обычно 30-60 минут
- 24. G 0 Наступает за фазой G1 Клетка экспрессирует белки, которые используются не для клеточного деления Гены,
- 25. Когда нормальные клетки прекращают делиться? Гены, включающие пролиферацию выключаются, если клетка получает сигнал, что все точки
- 26. Интегрины Интегрины-поверхностные рецепторы клетки, переносящие сигналы от внеклеточного матрикса к цитоскелету. «включают» рост, метастазирование, опухолевых клеток,
- 27. Адгезионные молекулы c-kit Клетки стромы Эндотелиальные клетки ГСК Гиалуроновая кислота CD44 Фибронектин VLA-4 VCAM-1 LFA-1 ICAM-1
- 28. Хоминг Костный мозг Сосуд Остеобласты SDF Остеоциты Стромальные клетки Эндотелиальные клетки ГСК CXCR4 Главные агенты: SDF-1/CXCR4
- 29. Figure 1. G-CSF HSC mobilization. a: There are adhesive interactions between HSC and matrix components in
- 31. ОСНОВНЫЕ ПОВЕРХНОСТНЫЕ МАРКЕРЫ ЭК CD 62 E (E-селектин) CD 62 P (Р-селектин) CD 106 (VCAM-1) CD
- 32. Роль циклинов Деление клетки запускается MPF В клетках млекопитающих существует семейство циклинзависимых киназ (cyclin-dependent kinases –Cdk).
- 33. Циклин В Циклин В – контролирует вхождение клеток в митоз. Состоит из 2 субьединиц: - киназного
- 34. MPF (циклин В) Разрушение MPF циклином Д вызывает выход из митоза. MPF участвует в начальных этапах
- 35. Митоз Сопровождается: Компактизацией, конденсацией хромосом, в 10 000 раз в нуклеосомы в середине G2 фазы Растворением
- 36. Cell Cycle – Representative Image Hoechst dye and 3 antibodies enable identification of Cells in G0,
- 37. ДНК- цитометрия а д т Д- диплоидные клетки А-клетки в апоптозе Т- тетраплоидные клетки анеуплоидия
- 38. Белок р 53 Точная репликация и распределение генетического материала – это важнейшее условие выживания клетки. В
- 39. Делеция 17p перед началом терапии Прогрессия на фоне лечения или рецидив в течение 12 месяцев от
- 40. N Ключевая роль p53 в резистентности 17p- До лечения
- 41. Делеция 17p Прогрессия на фоне лечения или рецидив в течение 12 месяцев от начала терапии Преимущественное
- 42. Протоонкогены – гены, чьи продукты участвуют в регуляции клеточной пролиферации. Мутации этих генов приводят к неконтролируемой
- 43. ЛКМЗ В-клеточная опухоль, характеризующаяся транслокацией или амплификацией локуса 11q13, приводящими к гиперэкспрессии ядерного белка циклина D1
- 44. 1-2 – толстые ригидные складки; 3-5 – лимфатические фолликулы подслизистого слоя; 6-8 – лимфоматозный полипоз; 9-12
- 45. ЛКМЗ Ядерная экспрессия циклина Д1 клетками костномозгового субстрата лимфомы из клеток мантии. Иммуногистохимия, ув. 250
- 46. Регуляция клеточного цикла в тканях При разработке метода выращивания клеток в культуре, было отмечено, что клетки
- 47. Факторы роста Факторы роста Семейство тромбоцитарных факторов роста (PDGF) : PDGF A, PDGF B, VEGF, PLGF,CSF-1
- 48. Старение - феномен Хейфлика. Клетки запрограммированы на определенное число делений, а затем прекращают делиться Репликативное старение
- 49. Теломеры Теломеры-это концевые участки хромосом, которые содержат повторяющиеся последовательности ДНК и укорачиваются с каждой репликацией. В
- 50. Апоптоз – программируемая клеточная гибель. Регуляция процессов клеточного деления необходима для выживания организма. Размножение клеток зависит
- 51. Апоптоз – программируемая клеточная гибель. француз Бонне различал следующие четыре типа ядерной дегенерации: Кариорексис - хроматин
- 52. Этапы апоптоза Каскад апоптотической гибели включает: Конденсацию хроматина, разрезание нитей хроматина эндонуклеазой на равные отрезки около
- 53. Механизмы апоптоза 2 фазы: Формирование и проведение апоптотического сигнала –фаза принятия решения Демонтаж клеточных структур –эффекторная
- 54. Каспазы Семейство протеаз, участвующих в апоптозе Существует 14 видов каспаз Активаторы цитокинов (каспазы 1,4, 5, 13)
- 55. Сигнальные пути апоптоза Повреждение ДНК, радиация, токсические воздействия, глюкокортикоиды, укорочение теломер приводит к активации каспазы 9
- 56. Изменения мембран апоптотических клеток Клеточная мембрана апоптозной клетки теряет сиаловую кислоту на гликопротеинах и гликолипидах, входящих
- 57. слияние в результате слияния получаются гибридомы СХЕМА ПОЛУЧЕНИЯ ГИБРИДОМ для производства моноклональных антител бессмертная клеточная линия
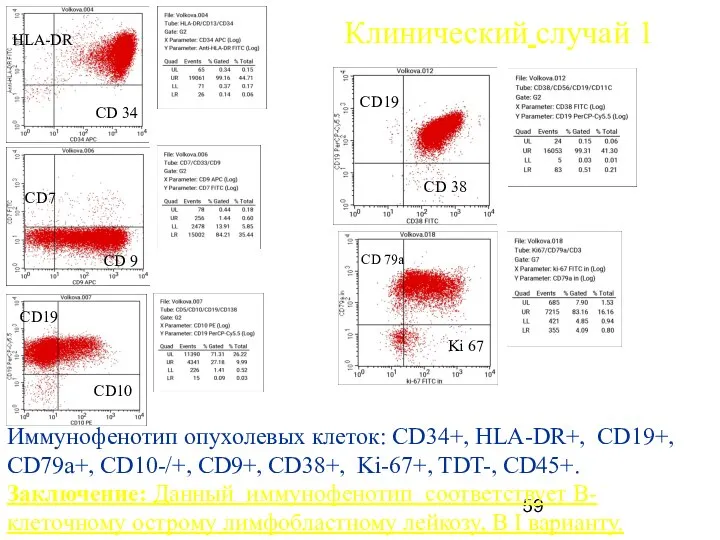
- 58. Клинический случай 1 Б-ая В.66 лет , поступила в отделение гематологии 21.05.2007 , ОАК: L-8.66*10 9/л,
- 59. HLA-DR CD 34 CD7 CD 9 Клинический случай 1 Иммунофенотип опухолевых клеток: СD34+, HLA-DR+, СD19+, СD79а+,
- 60. Клинический случай 1
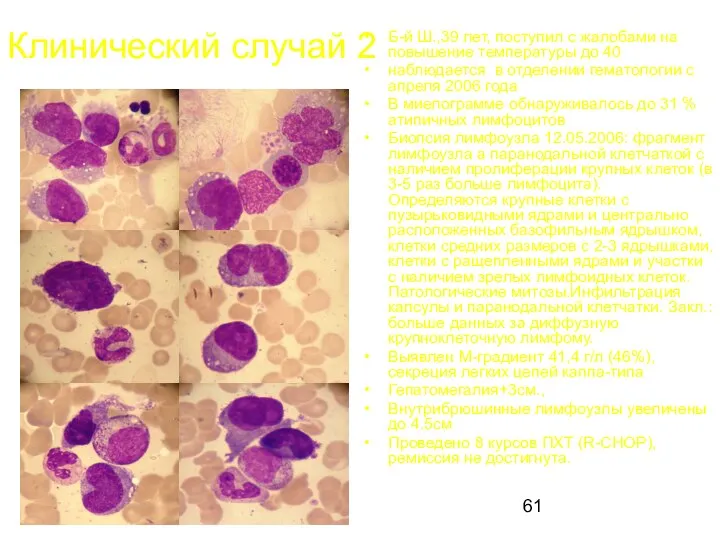
- 61. Клинический случай 2 Б-й Ш.,39 лет, поступил с жалобами на повышение температуры до 40 наблюдается в
- 62. Клинический случай 2 Заключение: Данный иммунофенотип в наибольшей степени соответствует плазмоклеточной пролиферации. Иммунофенотип опухолевых клеток: СD138+,
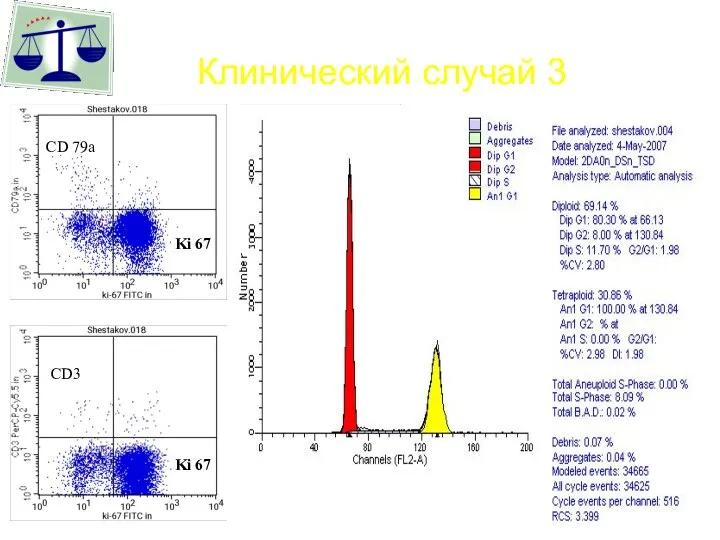
- 63. Клинический случай 3 Ki 67 Ki 67 CD 79a CD3
- 64. Комбинация флудара + ритуксимаб Функциональный синергизм CD55, CD46, CD59 CD 55 мешает действию ритуксимаба Флударабин ингибирует
- 65. Совместное российское исследование
- 66. Спасибо, что не вздремнули!
- 67. ЛИТЕРАТУРА Кассирский И.А. и Алексеев Г.А. Клиническая гематология. М., 1970; Максимов А.А. Основы гистологии, ч. 1—2,
- 69. Скачать презентацию
Введение
Введение

I Прибайкальская межрегиональная
научно-практическая конференция (2006 год)
I Прибайкальская межрегиональная
научно-практическая конференция (2006 год)

Кроветворение
Кроветворение (синоним гематопоэз) — процесс, заключающийся в серии клеточных дифференцировок, в результате
Кроветворение
Кроветворение (синоним гематопоэз) — процесс, заключающийся в серии клеточных дифференцировок, в результате
Сигнальные системы клетки
Сигнальные системы клетки

Биология сигнальных систем клетки
Кроветворные клетки имеют рецепторы к экзогенным сигнальным молекулам:
Биология сигнальных систем клетки
Кроветворные клетки имеют рецепторы к экзогенным сигнальным молекулам:

Киназы и фосфатазы
Фосфорилирование и дефосфорилирование - основные механизмы внутриклеточной передачи сигнала
В
Киназы и фосфатазы
Фосфорилирование и дефосфорилирование - основные механизмы внутриклеточной передачи сигнала
В

Рецепторы факторов роста
Рецепторы факторов роста обычно стимулируют клеточный рост, однако при
Рецепторы факторов роста
Рецепторы факторов роста обычно стимулируют клеточный рост, однако при
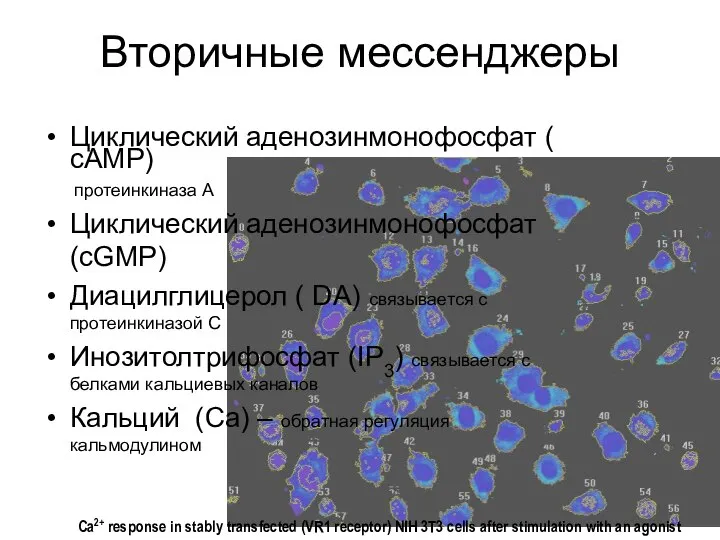
Вторичные мессенджеры
Циклический аденозинмонофосфат ( cAMP)
протеинкиназа А
Циклический аденозинмонофосфат (cGMP)
Диацилглицерол ( DA)
Вторичные мессенджеры
Циклический аденозинмонофосфат ( cAMP)
протеинкиназа А
Циклический аденозинмонофосфат (cGMP)
Диацилглицерол ( DA)

Способы доставки сигнальных молекул к клеткам
Эндокринный механизм
Паракринный механизм – клетка секретирует
Способы доставки сигнальных молекул к клеткам
Эндокринный механизм
Паракринный механизм – клетка секретирует

Рецепторы клеток
Группа 1 Семейство липофильных рецепторов:
Стероиды: ГК, МК, половые стероиды
Тиреоидные гормоны,
Рецепторы клеток
Группа 1 Семейство липофильных рецепторов:
Стероиды: ГК, МК, половые стероиды
Тиреоидные гормоны,
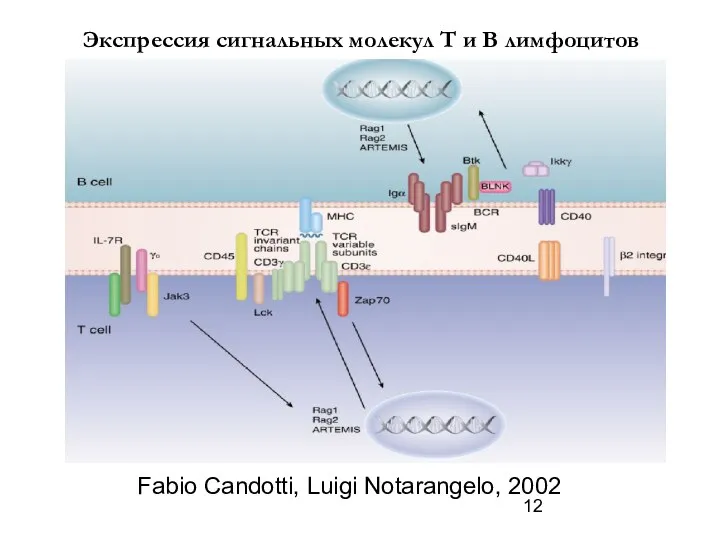
Fabio Candotti, Luigi Notarangelo, 2002
Экспрессия сигнальных молекул Т и В лимфоцитов
Fabio Candotti, Luigi Notarangelo, 2002
Экспрессия сигнальных молекул Т и В лимфоцитов
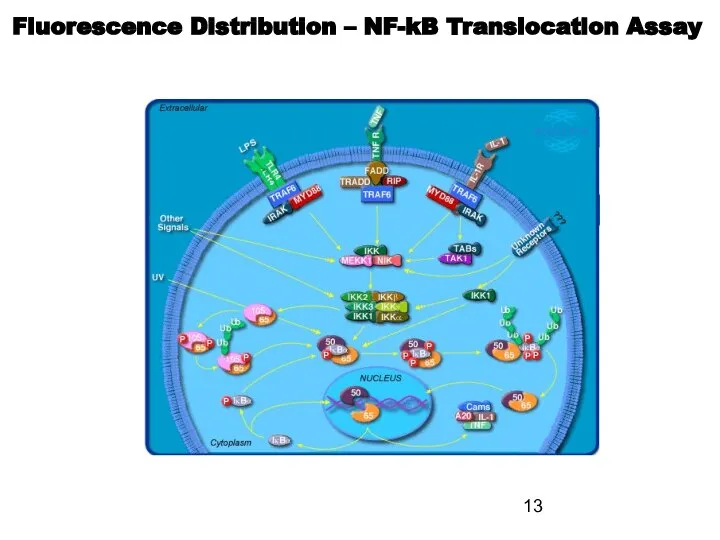
Fluorescence Distribution – NF-kB Translocation Assay
Fluorescence Distribution – NF-kB Translocation Assay
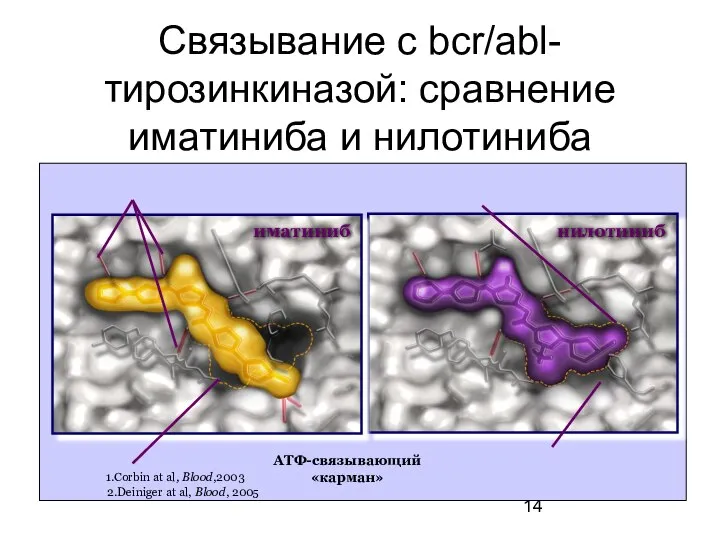
Связывание с bcr/abl-тирозинкиназой: сравнение иматиниба и нилотиниба
Связывание с bcr/abl-тирозинкиназой: сравнение иматиниба и нилотиниба

Внутриклеточные сигнальные пути
Кроме активации ras-путиКроме активации ras-пути , в клетках существуют
Внутриклеточные сигнальные пути
Кроме активации ras-путиКроме активации ras-пути , в клетках существуют

Свойства факторов роста
Факторами роста называют группу белковых молекул, индуцирующих синтез ДНКФакторами
Свойства факторов роста
Факторами роста называют группу белковых молекул, индуцирующих синтез ДНКФакторами
К Л Е Т О Ч Ы Й Ц И К
К Л Е Т О Ч Ы Й Ц И К

Трубочки, колечки и висюльки
Первым, как отмечают историки, клетку увидел Роберт Гук в 1663 году.
Трубочки, колечки и висюльки
Первым, как отмечают историки, клетку увидел Роберт Гук в 1663 году.

Митотический цикл
Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период
Митотический цикл
Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период
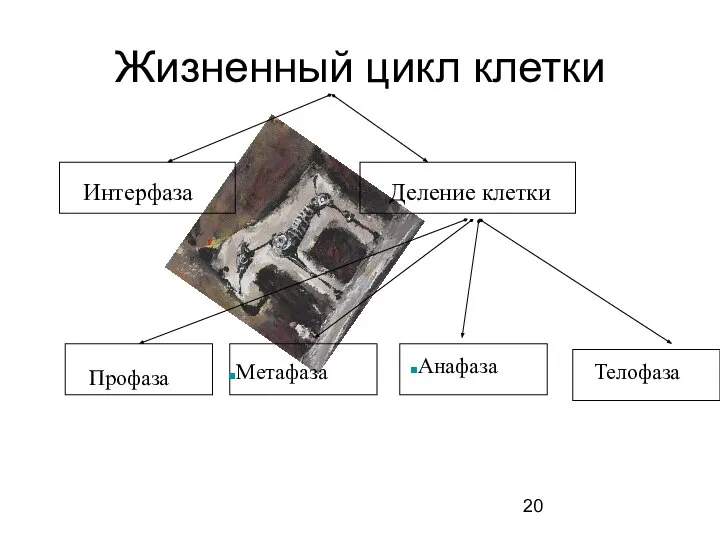
Жизненный цикл клетки
Профаза
Метафаза
Анафаза
Телофаза
Деление клетки
Интерфаза
Жизненный цикл клетки
Профаза
Метафаза
Анафаза
Телофаза
Деление клетки
Интерфаза


Митотический цикл
Митотический цикл = фаза М – это цикл деления,
Митотический цикл
Митотический цикл = фаза М – это цикл деления,


G 0
Наступает за фазой G1
Клетка экспрессирует белки, которые используются не для
G 0
Наступает за фазой G1
Клетка экспрессирует белки, которые используются не для

Когда нормальные клетки прекращают делиться?
Гены, включающие пролиферацию выключаются, если клетка получает
Когда нормальные клетки прекращают делиться?
Гены, включающие пролиферацию выключаются, если клетка получает

Интегрины
Интегрины-поверхностные рецепторы клетки, переносящие сигналы от внеклеточного матрикса к цитоскелету.
«включают» рост,
Интегрины
Интегрины-поверхностные рецепторы клетки, переносящие сигналы от внеклеточного матрикса к цитоскелету.
«включают» рост,
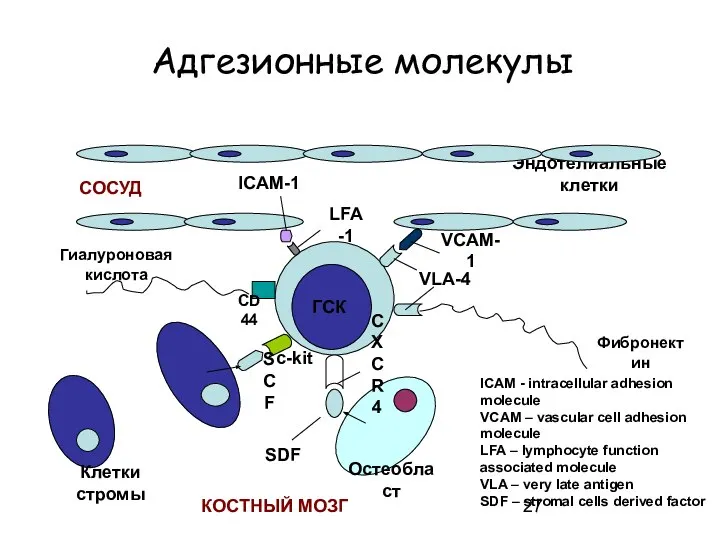
Адгезионные молекулы
c-kit
Клетки стромы
Эндотелиальные клетки
ГСК
Гиалуроновая кислота
CD44
Фибронектин
VLA-4
VCAM-1
LFA-1
ICAM-1
ICAM - intracellular adhesion
molecule
VCAM – vascular
Адгезионные молекулы
c-kit
Клетки стромы
Эндотелиальные клетки
ГСК
Гиалуроновая кислота
CD44
Фибронектин
VLA-4
VCAM-1
LFA-1
ICAM-1
ICAM - intracellular adhesion
molecule
VCAM – vascular
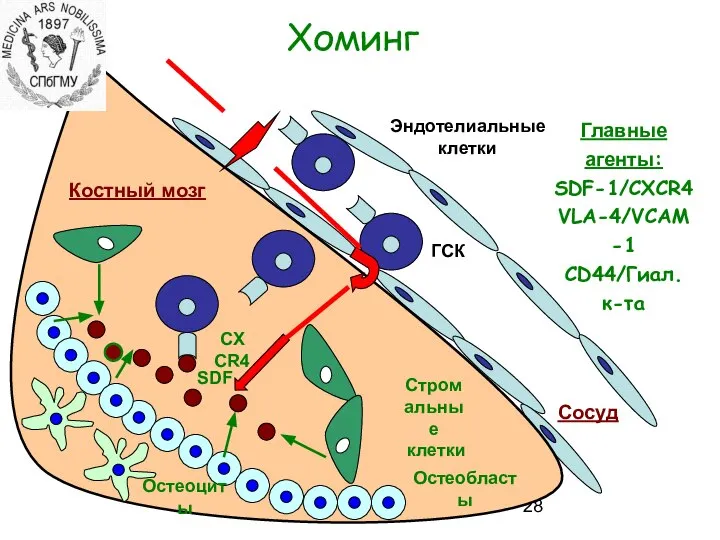
Хоминг
Костный мозг
Сосуд
Остеобласты
SDF
Остеоциты
Стромальные
клетки
Эндотелиальные
клетки
ГСК
CXCR4
Главные агенты:
SDF-1/CXCR4
VLA-4/VCAM-1
CD44/Гиал.к-та
Хоминг
Костный мозг
Сосуд
Остеобласты
SDF
Остеоциты
Стромальные
клетки
Эндотелиальные
клетки
ГСК
CXCR4
Главные агенты:
SDF-1/CXCR4
VLA-4/VCAM-1
CD44/Гиал.к-та
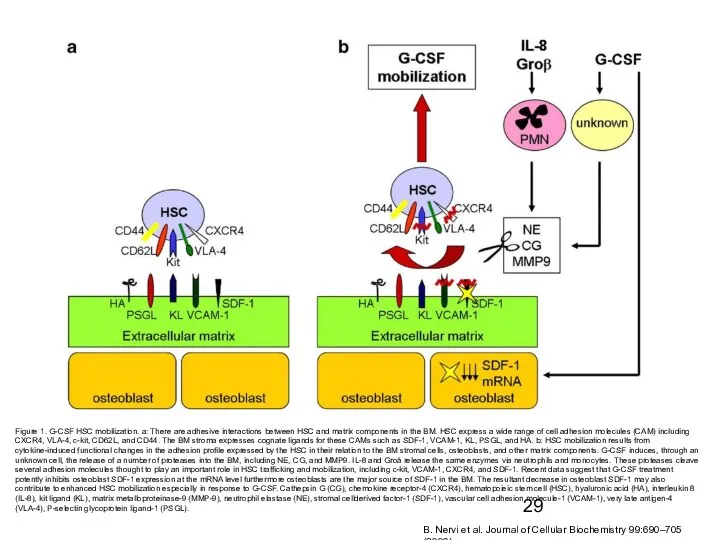
Figure 1. G-CSF HSC mobilization. a: There are adhesive interactions between
Figure 1. G-CSF HSC mobilization. a: There are adhesive interactions between

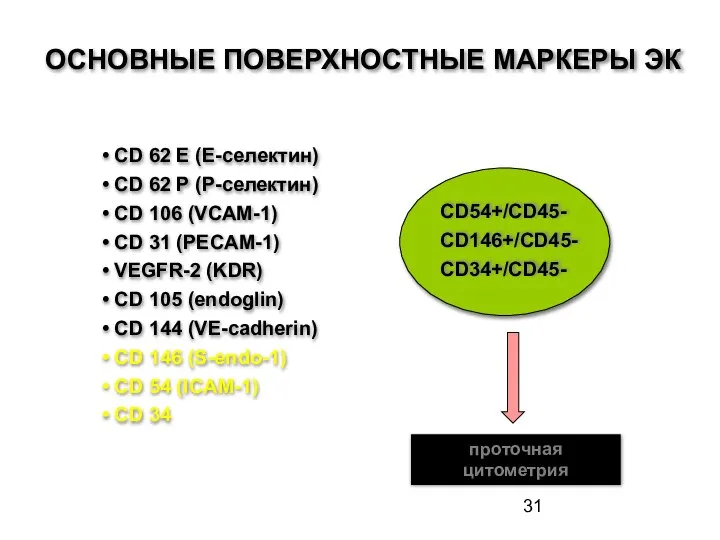
ОСНОВНЫЕ ПОВЕРХНОСТНЫЕ МАРКЕРЫ ЭК
CD 62 E (E-селектин)
CD 62 P
ОСНОВНЫЕ ПОВЕРХНОСТНЫЕ МАРКЕРЫ ЭК
CD 62 E (E-селектин)
CD 62 P
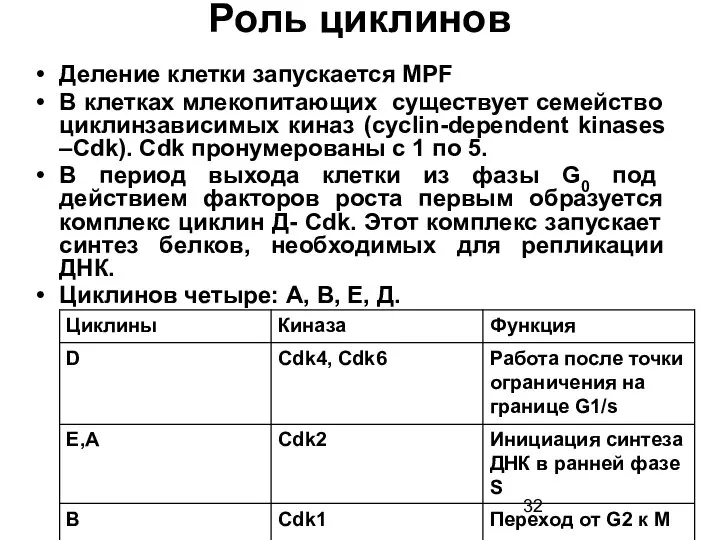
Роль циклинов
Деление клетки запускается MPF
В клетках млекопитающих существует семейство циклинзависимых
Роль циклинов
Деление клетки запускается MPF
В клетках млекопитающих существует семейство циклинзависимых

Циклин В
Циклин В – контролирует вхождение клеток в митоз.
Состоит из
Циклин В
Циклин В – контролирует вхождение клеток в митоз.
Состоит из

MPF (циклин В)
Разрушение MPF циклином Д вызывает выход из митоза.
MPF
MPF (циклин В)
Разрушение MPF циклином Д вызывает выход из митоза.
MPF

Митоз
Сопровождается:
Компактизацией, конденсацией хромосом, в
10 000 раз в нуклеосомы в
Митоз
Сопровождается:
Компактизацией, конденсацией хромосом, в
10 000 раз в нуклеосомы в
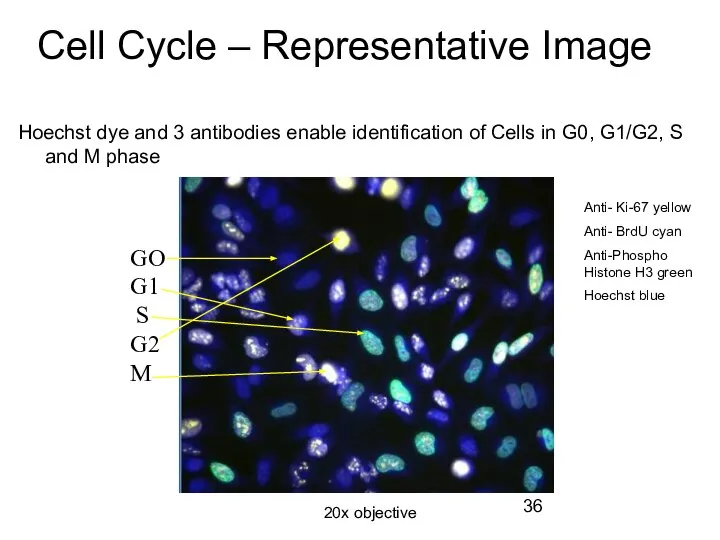
Cell Cycle – Representative Image
Hoechst dye and 3 antibodies enable identification
Cell Cycle – Representative Image
Hoechst dye and 3 antibodies enable identification
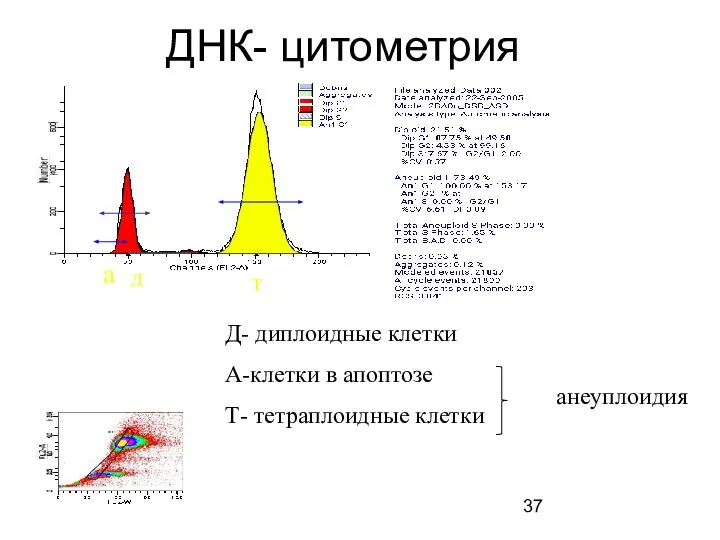
ДНК- цитометрия
а
д
т
Д- диплоидные клетки
А-клетки в апоптозе
Т- тетраплоидные клетки
анеуплоидия
ДНК- цитометрия
а
д
т
Д- диплоидные клетки
А-клетки в апоптозе
Т- тетраплоидные клетки
анеуплоидия
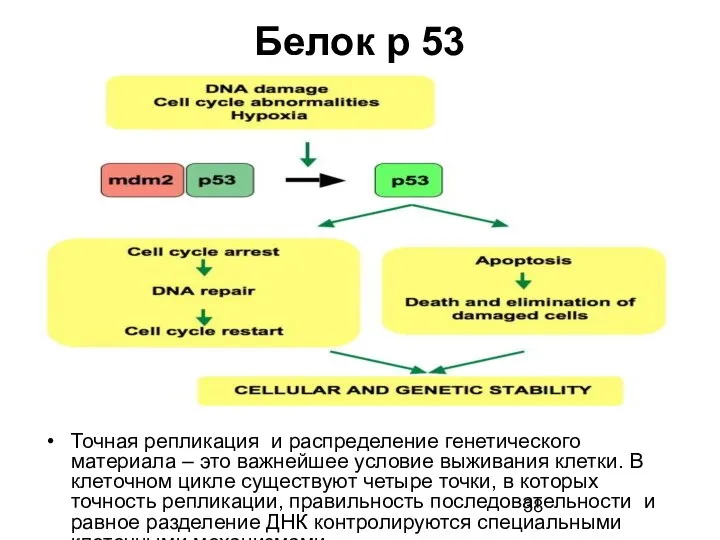
Белок р 53
Точная репликация и распределение генетического материала – это
Белок р 53
Точная репликация и распределение генетического материала – это

Делеция 17p перед началом терапии
Прогрессия на фоне лечения или рецидив в
Делеция 17p перед началом терапии
Прогрессия на фоне лечения или рецидив в
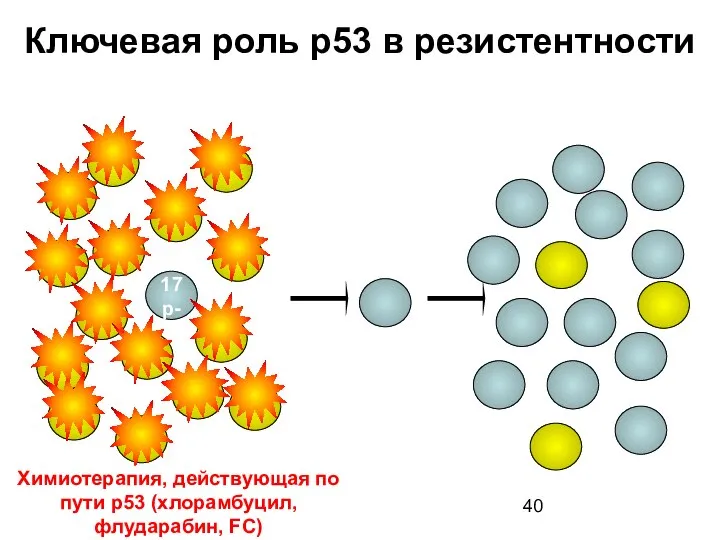
N
Ключевая роль p53 в резистентности
17p-
До лечения
N
Ключевая роль p53 в резистентности
17p-
До лечения

Делеция 17p
Прогрессия на фоне лечения или рецидив в течение 12 месяцев
Делеция 17p
Прогрессия на фоне лечения или рецидив в течение 12 месяцев
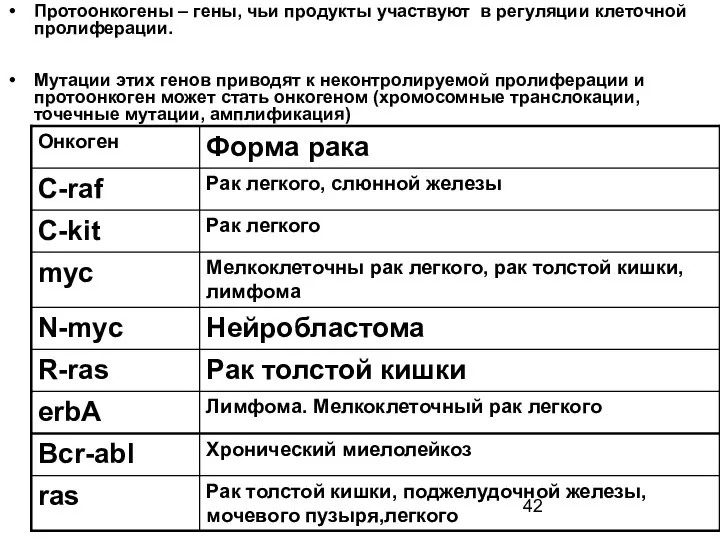
Протоонкогены – гены, чьи продукты участвуют в регуляции клеточной пролиферации.
Мутации этих
Протоонкогены – гены, чьи продукты участвуют в регуляции клеточной пролиферации.
Мутации этих

ЛКМЗ
В-клеточная опухоль, характеризующаяся транслокацией или амплификацией локуса 11q13, приводящими к
ЛКМЗ
В-клеточная опухоль, характеризующаяся транслокацией или амплификацией локуса 11q13, приводящими к
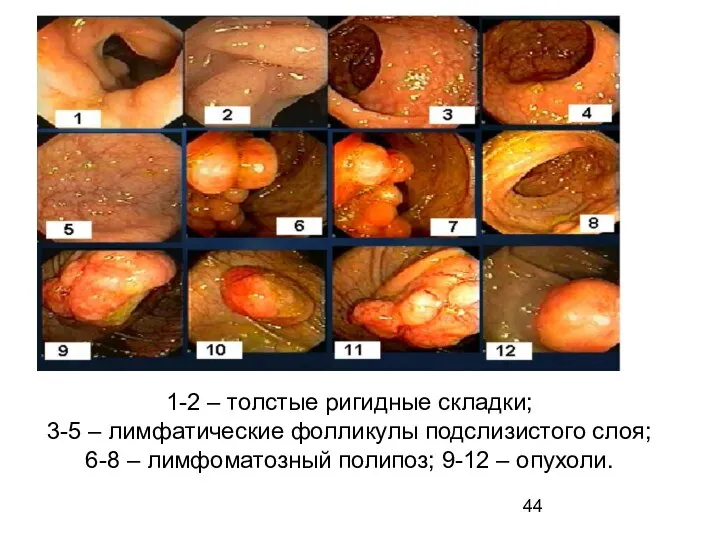
1-2 – толстые ригидные складки;
3-5 – лимфатические фолликулы подслизистого слоя;
1-2 – толстые ригидные складки; 3-5 – лимфатические фолликулы подслизистого слоя;
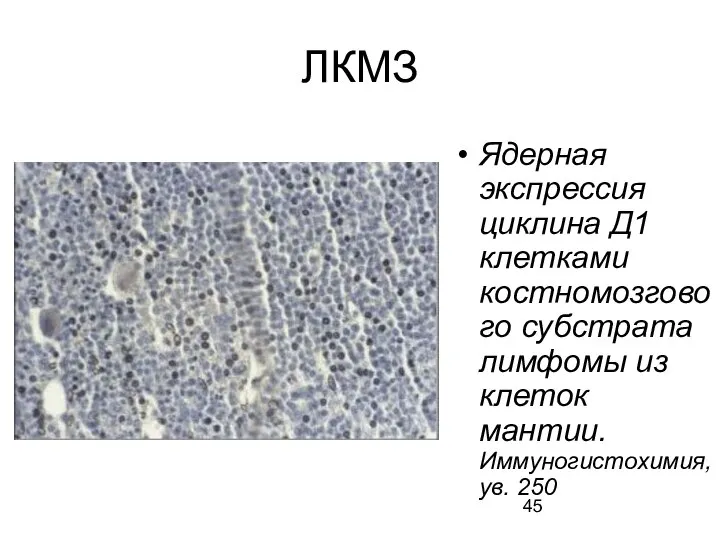
ЛКМЗ
Ядерная экспрессия циклина Д1 клетками костномозгового субстрата лимфомы из клеток мантии.
ЛКМЗ
Ядерная экспрессия циклина Д1 клетками костномозгового субстрата лимфомы из клеток мантии.
Регуляция клеточного цикла в тканях
При разработке метода выращивания клеток в культуре,
Регуляция клеточного цикла в тканях
При разработке метода выращивания клеток в культуре,

Факторы роста
Факторы роста
Семейство тромбоцитарных факторов роста (PDGF) : PDGF A, PDGF
Факторы роста
Факторы роста
Семейство тромбоцитарных факторов роста (PDGF) : PDGF A, PDGF

Старение - феномен Хейфлика.
Клетки запрограммированы на определенное число делений, а затем
Старение - феномен Хейфлика.
Клетки запрограммированы на определенное число делений, а затем

Теломеры
Теломеры-это концевые участки хромосом, которые содержат повторяющиеся последовательности ДНК и укорачиваются
Теломеры
Теломеры-это концевые участки хромосом, которые содержат повторяющиеся последовательности ДНК и укорачиваются

Апоптоз – программируемая клеточная гибель.
Регуляция процессов клеточного деления необходима для выживания
Апоптоз – программируемая клеточная гибель.
Регуляция процессов клеточного деления необходима для выживания

Апоптоз – программируемая клеточная гибель.
француз Бонне различал следующие четыре типа ядерной
Апоптоз – программируемая клеточная гибель.
француз Бонне различал следующие четыре типа ядерной
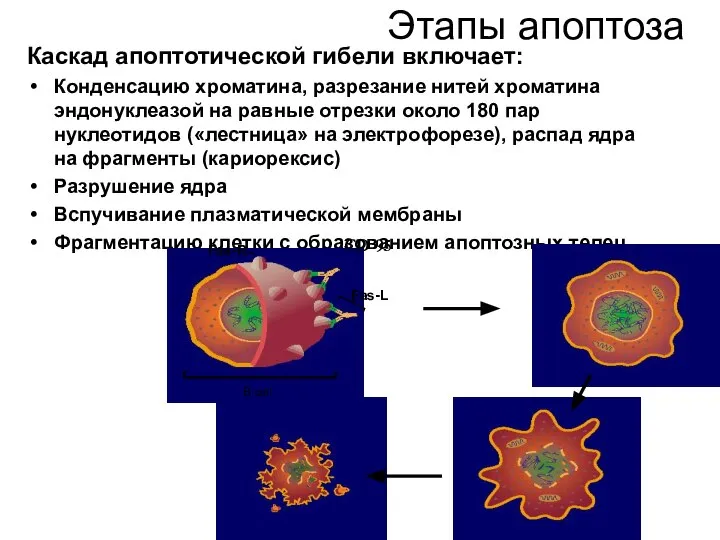
Этапы апоптоза
Каскад апоптотической гибели включает:
Конденсацию хроматина, разрезание нитей хроматина эндонуклеазой на
Этапы апоптоза
Каскад апоптотической гибели включает:
Конденсацию хроматина, разрезание нитей хроматина эндонуклеазой на

Механизмы апоптоза
2 фазы:
Формирование и проведение апоптотического сигнала –фаза принятия решения
Демонтаж клеточных
Механизмы апоптоза
2 фазы:
Формирование и проведение апоптотического сигнала –фаза принятия решения
Демонтаж клеточных

Каспазы
Семейство протеаз, участвующих в апоптозе
Существует 14 видов каспаз
Активаторы цитокинов (каспазы 1,4,
Каспазы
Семейство протеаз, участвующих в апоптозе
Существует 14 видов каспаз
Активаторы цитокинов (каспазы 1,4,

Сигнальные пути апоптоза
Повреждение ДНК, радиация, токсические воздействия, глюкокортикоиды, укорочение теломер приводит
Сигнальные пути апоптоза
Повреждение ДНК, радиация, токсические воздействия, глюкокортикоиды, укорочение теломер приводит

Изменения мембран апоптотических клеток
Клеточная мембрана апоптозной клетки теряет сиаловую кислоту на
Изменения мембран апоптотических клеток
Клеточная мембрана апоптозной клетки теряет сиаловую кислоту на
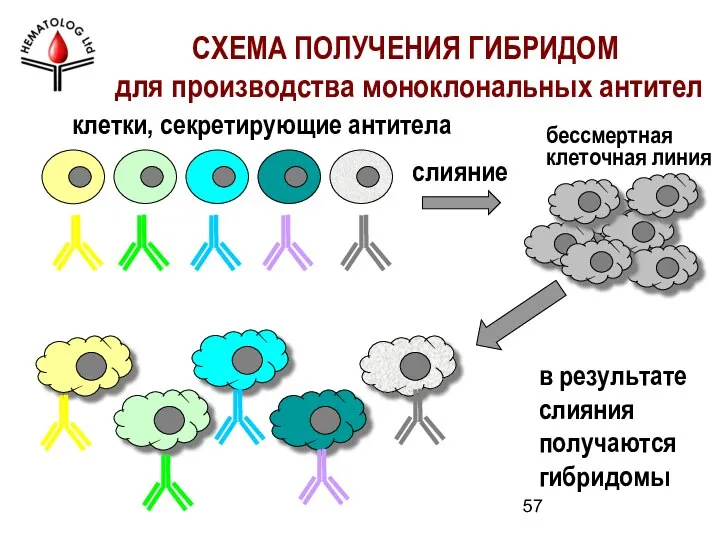
слияние
в результате
слияния
получаются
гибридомы
СХЕМА ПОЛУЧЕНИЯ ГИБРИДОМ
для производства моноклональных антител
бессмертная
клеточная линия
клетки, секретирующие
слияние
в результате
слияния
получаются
гибридомы
СХЕМА ПОЛУЧЕНИЯ ГИБРИДОМ
для производства моноклональных антител
бессмертная
клеточная линия
клетки, секретирующие
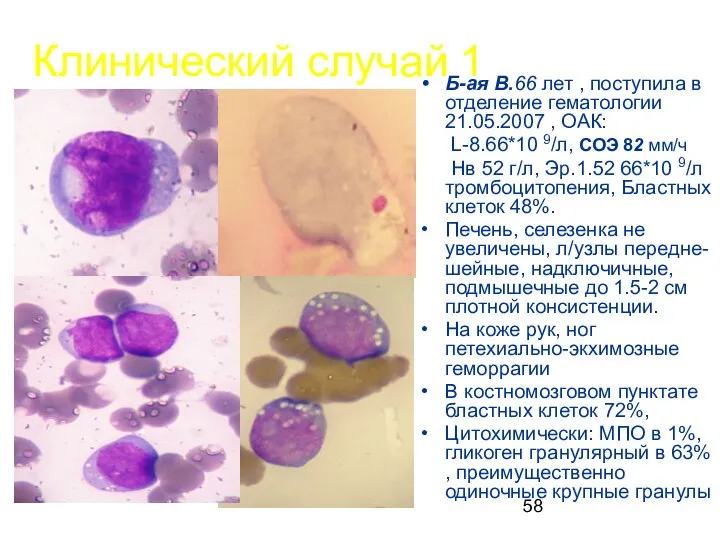
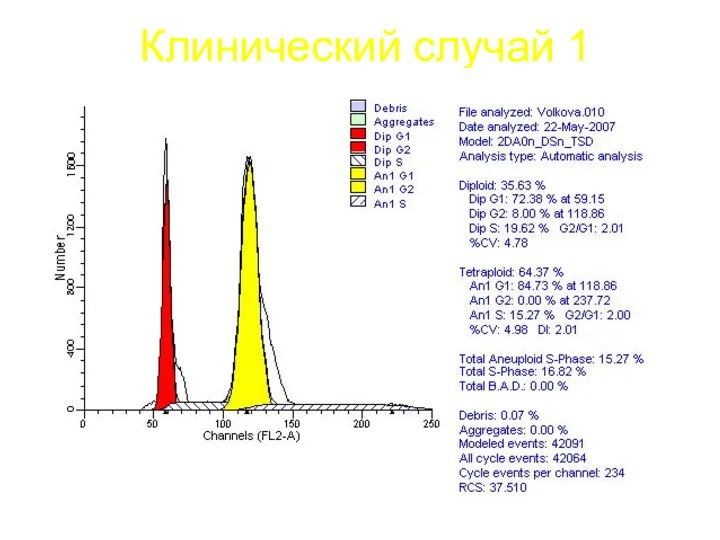
Клинический случай 1
Б-ая В.66 лет , поступила в отделение гематологии 21.05.2007 ,
Клинический случай 1
Б-ая В.66 лет , поступила в отделение гематологии 21.05.2007 ,
HLA-DR
CD 34
CD7
CD 9
Клинический случай 1
Иммунофенотип опухолевых клеток: СD34+, HLA-DR+, СD19+, СD79а+,
HLA-DR
CD 34
CD7
CD 9
Клинический случай 1
Иммунофенотип опухолевых клеток: СD34+, HLA-DR+, СD19+, СD79а+,
Клинический случай 1
Клинический случай 1
Клинический случай 2
Б-й Ш.,39 лет, поступил с жалобами на повышение температуры
Клинический случай 2
Б-й Ш.,39 лет, поступил с жалобами на повышение температуры

Клинический случай 2
Заключение: Данный иммунофенотип в наибольшей степени соответствует плазмоклеточной
Клинический случай 2
Заключение: Данный иммунофенотип в наибольшей степени соответствует плазмоклеточной
Клинический случай 3
Ki 67
Ki 67
CD 79a
CD3
Клинический случай 3
Ki 67
Ki 67
CD 79a
CD3
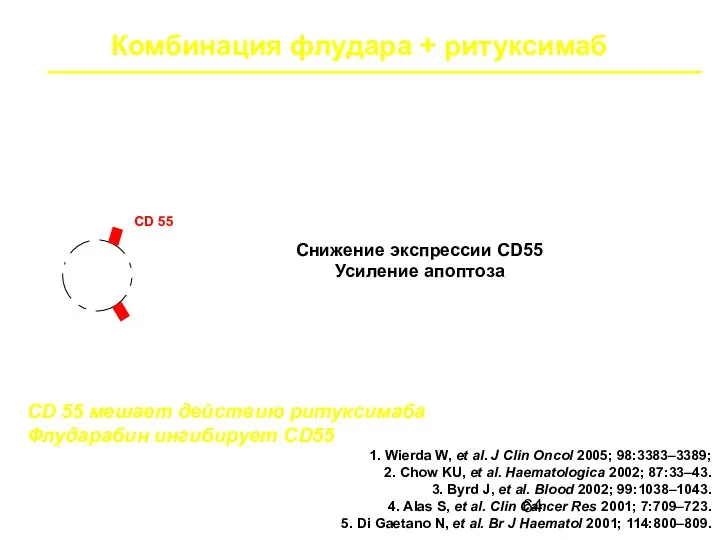
Комбинация флудара + ритуксимаб
Функциональный синергизм
CD55, CD46, CD59
CD
Комбинация флудара + ритуксимаб
Функциональный синергизм
CD55, CD46, CD59
CD

Совместное российское исследование
Совместное российское исследование

Спасибо, что не вздремнули!
Спасибо, что не вздремнули!

ЛИТЕРАТУРА
Кассирский И.А. и Алексеев Г.А. Клиническая гематология. М., 1970; Максимов А.А. Основы гистологии, ч.
ЛИТЕРАТУРА
Кассирский И.А. и Алексеев Г.А. Клиническая гематология. М., 1970; Максимов А.А. Основы гистологии, ч.
 Основные сведения о насосах
Основные сведения о насосах 01.10 Недостающая фигура
01.10 Недостающая фигура Презентация на тему "Боль в неврологии" - скачать презентации по Медицине
Презентация на тему "Боль в неврологии" - скачать презентации по Медицине Презентация Керамика
Презентация Керамика Оборотные засоби
Оборотные засоби Римская республика во 2 веке до н.э.
Римская республика во 2 веке до н.э. Викторина "Урок (час) футбола" - 2
Викторина "Урок (час) футбола" - 2 Абсолютные и относительные величины в статистике.
Абсолютные и относительные величины в статистике. Невербальные средства коммуникации
Невербальные средства коммуникации Особенности проектирования зданий для сейсмических районов
Особенности проектирования зданий для сейсмических районов . Лечение микозов
. Лечение микозов По ту сторону холста… Автор: Петрова Юлия 10 «а» класс МОУ СОШ №17
По ту сторону холста… Автор: Петрова Юлия 10 «а» класс МОУ СОШ №17  Тепловые двигатели
Тепловые двигатели Учебно-тренировочный сбор «Твоё здоровье». Правила
Учебно-тренировочный сбор «Твоё здоровье». Правила Комплексы с 5-ю 6-ю электронными лигандами
Комплексы с 5-ю 6-ю электронными лигандами  Презентация Понятие и характеристика единой системы таможенных органов РФ
Презентация Понятие и характеристика единой системы таможенных органов РФ Подготовила: Шестёро Алина
Подготовила: Шестёро Алина  Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2)
Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2) Технология обучения детей 9-10 лет плаванию в условиях оздоровительной группы
Технология обучения детей 9-10 лет плаванию в условиях оздоровительной группы Технологии цифрового информационного моделирования на этапе территориального планирования
Технологии цифрового информационного моделирования на этапе территориального планирования Моє хобі - футбол
Моє хобі - футбол Соціальна політика Італії
Соціальна політика Італії Решение задания В8 (ЕГЭ-2014) (анализ численного алгоритма)
Решение задания В8 (ЕГЭ-2014) (анализ численного алгоритма) Диаграмма состояний. Диаграмма, которая представляет конечный автомат
Диаграмма состояний. Диаграмма, которая представляет конечный автомат Устав Муниципального образовательного учреждения «Средняя общеобразовательная школа № 52»
Устав Муниципального образовательного учреждения «Средняя общеобразовательная школа № 52» определение расстояний до звезд
определение расстояний до звезд Moderní cukrovar HROCHŮV TÝNEC
Moderní cukrovar HROCHŮV TÝNEC babushkina_oet_316
babushkina_oet_316