- Механизмы центрального торможения

Содержание
- 2. Торможение – самостоятельный нервный процесс, который вызывается возбуждением и проявляется в подавлении другого возбуждения.
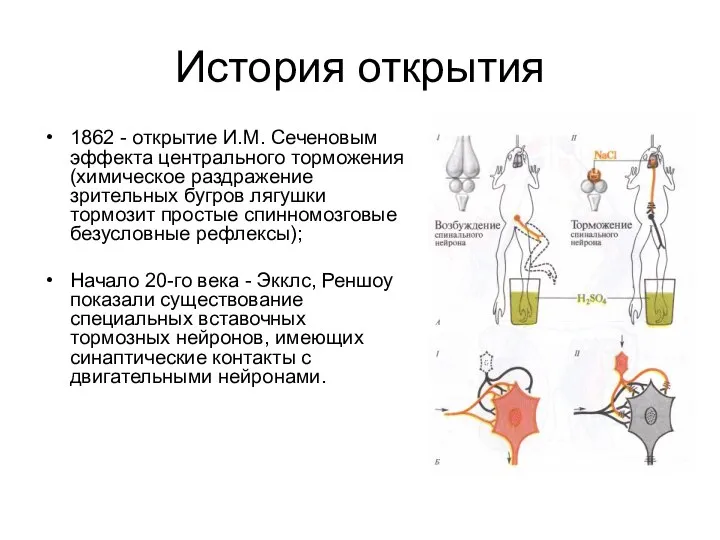
- 3. История открытия 1862 - открытие И.М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров лягушки тормозит
- 4. Механизмы центрального торможения В зависимости от нейронного механизма, различают первичное торможение, осуществляемое с помощью тормозных нейронов
- 5. Постсинаптическое торможение - основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендрических синапсов под влиянием
- 6. Пресинаптическое торможение Деполяризация постсинаптической области вызывает уменьшение амплитуды ПД, приходящего в пресинаптическое окончание возбуждающего нейрона (механизм
- 7. Пессимальное торможение Представляет собой вид торможения центральных нейронов. Наступает при высокой частоте раздражения. Не требует специальных
- 8. В зависимости от строения нейронных сетей различают три вида торможения: Возвратное; Реципрокное (сопряженное); Латеральное.
- 9. Возвратное торможение Угнетение активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки с участием тормозного вставочного нейрона.
- 10. Реципрокное (сопряженное) торможение Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений между нервными центрами благодаря
- 11. Латеральное торможение При латеральном торможении возбуждение, передаваемое через коллатерали аксона возбужденной нервной клетки, активирует вставочные тормозные
- 12. Значение торможения Координация рефлекторных актов. Направляет возбуждение к определенным нервным центрам или по определенному пути, выключая
- 13. Координация В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности отдельных нервных клеток и
- 14. Основные принципы координационного взаимодействия Сопряженное (реципрокное) торможение. Обратная связь. Положительная – сигналы, поступающие на вход системы
- 16. Скачать презентацию

Торможение – самостоятельный нервный процесс, который вызывается возбуждением и проявляется в
Торможение – самостоятельный нервный процесс, который вызывается возбуждением и проявляется в
История открытия
1862 - открытие И.М. Сеченовым эффекта центрального торможения (химическое раздражение
История открытия
1862 - открытие И.М. Сеченовым эффекта центрального торможения (химическое раздражение

Механизмы центрального торможения
В зависимости от нейронного механизма, различают первичное торможение, осуществляемое
Механизмы центрального торможения
В зависимости от нейронного механизма, различают первичное торможение, осуществляемое
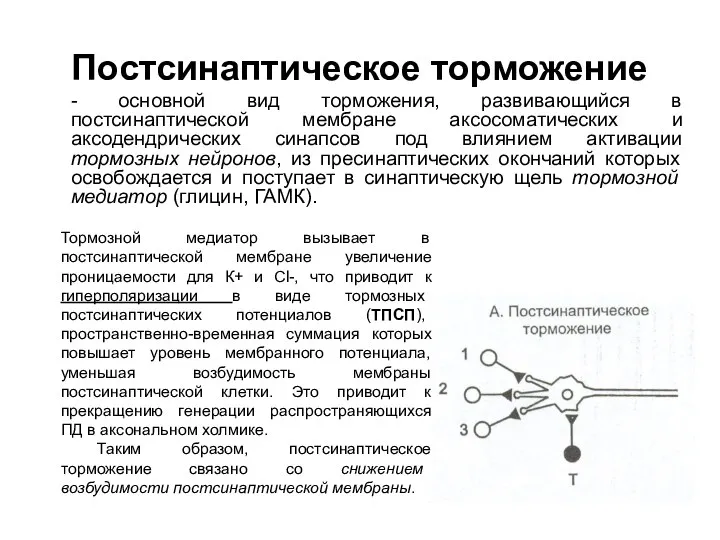
Постсинаптическое торможение
- основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и
Постсинаптическое торможение
- основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и

Пресинаптическое торможение
Деполяризация постсинаптической области вызывает уменьшение амплитуды ПД, приходящего в пресинаптическое
Пресинаптическое торможение
Деполяризация постсинаптической области вызывает уменьшение амплитуды ПД, приходящего в пресинаптическое

Пессимальное торможение
Представляет собой вид торможения центральных нейронов.
Наступает при высокой частоте
Пессимальное торможение
Представляет собой вид торможения центральных нейронов.
Наступает при высокой частоте

В зависимости от строения нейронных сетей различают три вида торможения:
Возвратное;
Реципрокное (сопряженное);
Латеральное.
В зависимости от строения нейронных сетей различают три вида торможения:
Возвратное;
Реципрокное (сопряженное);
Латеральное.
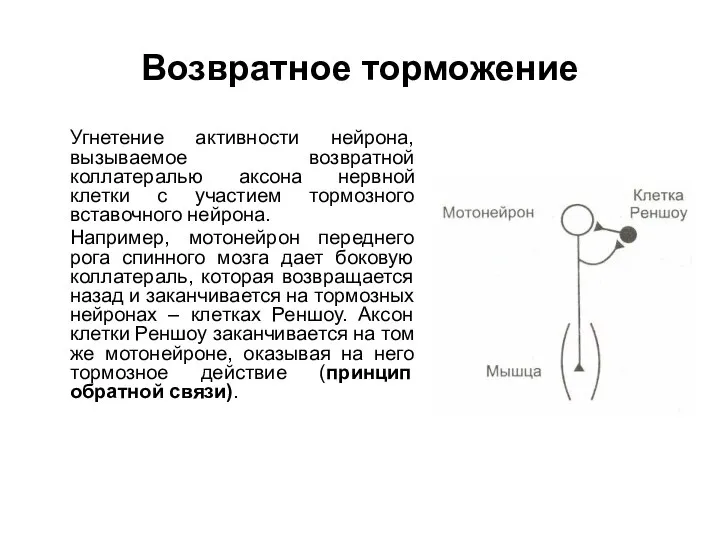
Возвратное торможение
Угнетение активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки с
Возвратное торможение
Угнетение активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки с
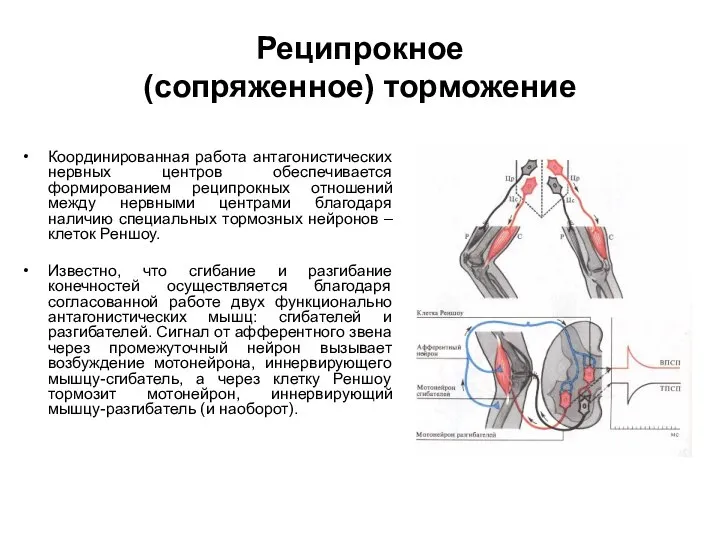
Реципрокное
(сопряженное) торможение
Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений
Реципрокное
(сопряженное) торможение
Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений

Латеральное торможение
При латеральном торможении возбуждение, передаваемое через коллатерали аксона возбужденной нервной
Латеральное торможение
При латеральном торможении возбуждение, передаваемое через коллатерали аксона возбужденной нервной

Значение торможения
Координация рефлекторных актов. Направляет возбуждение к определенным нервным центрам или
Значение торможения
Координация рефлекторных актов. Направляет возбуждение к определенным нервным центрам или

Координация
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности
Координация
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности

Основные принципы координационного взаимодействия
Сопряженное (реципрокное) торможение.
Обратная связь. Положительная – сигналы,
Основные принципы координационного взаимодействия
Сопряженное (реципрокное) торможение.
Обратная связь. Положительная – сигналы,
 Государственное бюджетное дошкольное образовательное учреждение детский сад №27 комбинированного вида Пушкинского района город
Государственное бюджетное дошкольное образовательное учреждение детский сад №27 комбинированного вида Пушкинского района город Прямоугольное проецирование. Расположение видов на чертеже
Прямоугольное проецирование. Расположение видов на чертеже Вестибулярный анализ
Вестибулярный анализ  ГОСУДАРСТВЕННЫЕ ЦЕННЫЕ БУМАГИ I. Общая характеристика II. Виды государственных ценных бумаг III. Ценные бумаги региональных ор
ГОСУДАРСТВЕННЫЕ ЦЕННЫЕ БУМАГИ I. Общая характеристика II. Виды государственных ценных бумаг III. Ценные бумаги региональных ор Общие технические свойства строительных материалов
Общие технические свойства строительных материалов Сумматор
Сумматор Преимущества ручного режима в фотоаппарате
Преимущества ручного режима в фотоаппарате Методологія та методи наукового дослідження
Методологія та методи наукового дослідження Маркетинг онлайн (тренды 2009) Евгений Шевченко Интернет-агентство UaMaster «THE MARKETING JAZZ FEST 2009»
Маркетинг онлайн (тренды 2009) Евгений Шевченко Интернет-агентство UaMaster «THE MARKETING JAZZ FEST 2009»  Средства по уходу «Актив-уход». Oriflame
Средства по уходу «Актив-уход». Oriflame Основные виды помех средствам активной радиолокации
Основные виды помех средствам активной радиолокации Презентация Административное право
Презентация Административное право  Площадь Академгородка. Реконструкция
Площадь Академгородка. Реконструкция Домены из первых рук. Нераскрытый потенциал для маркетолога Федор Смирнов зам. директора департамента PR fos@nic.ru. - презентация
Домены из первых рук. Нераскрытый потенциал для маркетолога Федор Смирнов зам. директора департамента PR fos@nic.ru. - презентация Аварии на объектах коммунального хозяйства Майстренко Денис Т-115
Аварии на объектах коммунального хозяйства Майстренко Денис Т-115 Анализ некоторых систем электронных денежных средств
Анализ некоторых систем электронных денежных средств Человек политический. Поведение и участие
Человек политический. Поведение и участие Истинный солдат своего Отечества. 2019 год - год Даниила Гранина
Истинный солдат своего Отечества. 2019 год - год Даниила Гранина Два лица Кореи
Два лица Кореи Развитие спортивно-оздоровительного туризма в России
Развитие спортивно-оздоровительного туризма в России Политические партии и движения
Политические партии и движения Личность Ивана 3 и его деятельность
Личность Ивана 3 и его деятельность Анализ характера конкуренции и рыночных действий конкурентов при сбыте товаров на зарубежный рынок
Анализ характера конкуренции и рыночных действий конкурентов при сбыте товаров на зарубежный рынок ПОЗНАВАТЕЛЬНАЯ АКТИВНОСТЬ МЛАДШИХ ШКОЛЬНИКОВ Уровни активности: Репродуктивно-подражательная активность Поисково-исполните
ПОЗНАВАТЕЛЬНАЯ АКТИВНОСТЬ МЛАДШИХ ШКОЛЬНИКОВ Уровни активности: Репродуктивно-подражательная активность Поисково-исполните Рекурсия
Рекурсия Информационное обеспечение финансового менеджмента
Информационное обеспечение финансового менеджмента  Сучасні вимоги до оформлення службових документів
Сучасні вимоги до оформлення службових документів Розробка бібліотеки для динамічного завантаження виконуваного коду в запущений процес
Розробка бібліотеки для динамічного завантаження виконуваного коду в запущений процес