- Нейронные сети. Принципы взаимодействия нейронов в нейронных сетях.

Содержание
- 2. Нейронные сети мозга – совокупность синаптически связанных нейронов, участвующих в получении, передаче, хранении и воспроизведении информации.
- 3. Нейрон имеет множественные синаптические контакты с другими нейронами Три принципа взаимодействия нейронов: 1. Принцип дивергенции. 2.
- 4. Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или
- 5. Нервные центры имеют ряд общих свойств, определяемых наличием синаптических образований и структурой нейронных сетей, образующих эти
- 6. Закономерности проведения возбуждения по рефлекторной дуге Одностороннее проведение; Замедленное проведение; Суммация подпороговых раздражений; Трансформация ритма возбуждения;
- 7. Одностороннее проведение. Обусловлено особенностями проведения возбуждения по химическому синапсу. Медиаторы, к которым рецепторы находятся в постсинаптической
- 8. Суммация подпороговых раздражений. Два типа: Суммация временная; П/п раздражение наносится на одну и ту же точку
- 9. Трансформация ритма возбуждения. При ритмическом возбуждении нервный центр перестраивает ритм как понижая, так и повышая частоту
- 10. Облегчение или потенциация - усиление рефлекторного ответа во время частотного раздражения. Посттетаническая потенциация – это усиление
- 12. Скачать презентацию
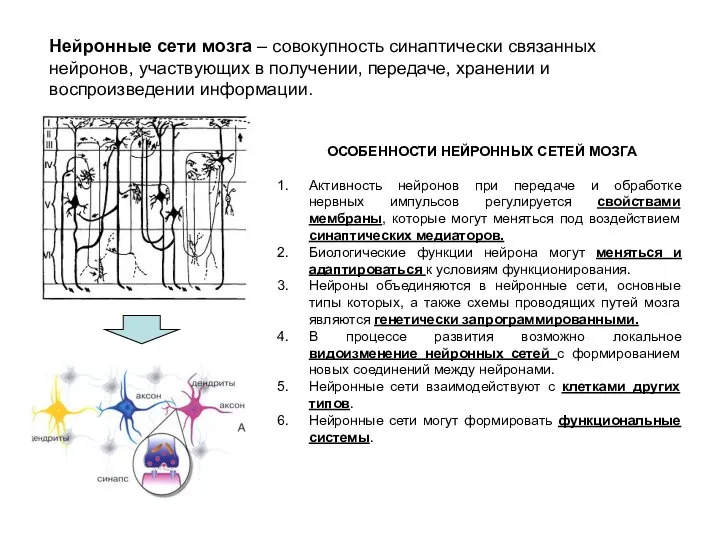
Нейронные сети мозга – совокупность синаптически связанных нейронов, участвующих в получении,
Нейронные сети мозга – совокупность синаптически связанных нейронов, участвующих в получении,
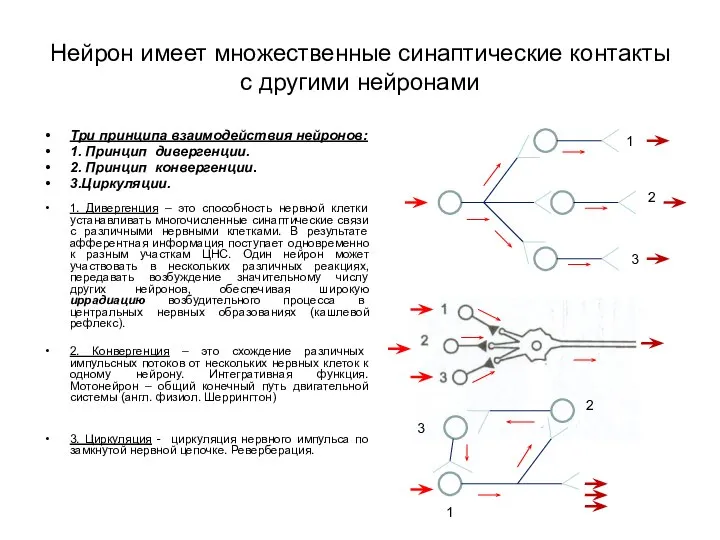
Нейрон имеет множественные синаптические контакты с другими нейронами
Три принципа взаимодействия нейронов:
1.
Нейрон имеет множественные синаптические контакты с другими нейронами
Три принципа взаимодействия нейронов:
1.

Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает
Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает

Нервные центры имеют ряд общих свойств, определяемых наличием синаптических образований и
Нервные центры имеют ряд общих свойств, определяемых наличием синаптических образований и

Закономерности проведения возбуждения по рефлекторной дуге
Одностороннее проведение;
Замедленное проведение;
Суммация подпороговых раздражений;
Трансформация ритма
Закономерности проведения возбуждения по рефлекторной дуге
Одностороннее проведение;
Замедленное проведение;
Суммация подпороговых раздражений;
Трансформация ритма
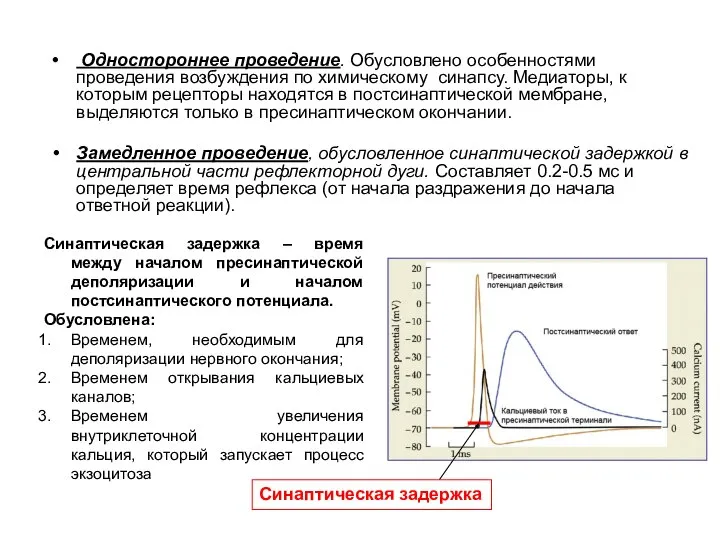
Одностороннее проведение. Обусловлено особенностями проведения возбуждения по химическому синапсу. Медиаторы, к
Одностороннее проведение. Обусловлено особенностями проведения возбуждения по химическому синапсу. Медиаторы, к

Суммация подпороговых раздражений.
Два типа:
Суммация временная;
П/п раздражение наносится на одну и ту
Суммация подпороговых раздражений.
Два типа:
Суммация временная;
П/п раздражение наносится на одну и ту

Трансформация ритма возбуждения.
При ритмическом возбуждении нервный центр перестраивает ритм как понижая,
Трансформация ритма возбуждения.
При ритмическом возбуждении нервный центр перестраивает ритм как понижая,
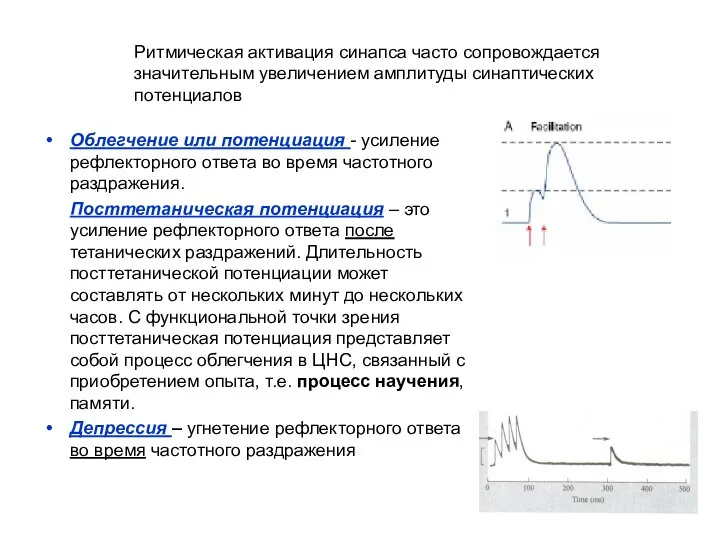
Облегчение или потенциация - усиление рефлекторного ответа во время частотного раздражения.
Облегчение или потенциация - усиление рефлекторного ответа во время частотного раздражения.
 Грех и его трактовка в разных религиях
Грех и его трактовка в разных религиях Что такое история? исторические источники
Что такое история? исторические источники  Множества и операции над ними - презентация по Алгебре
Множества и операции над ними - презентация по Алгебре Система отраслевого права (подготовка к ЕГЭ)
Система отраслевого права (подготовка к ЕГЭ) Компьютер. Устройства компьютера
Компьютер. Устройства компьютера Perfekt mit haben oder sein? Ordnen im Perfekt
Perfekt mit haben oder sein? Ordnen im Perfekt 16 грипп лекц.ppt
16 грипп лекц.ppt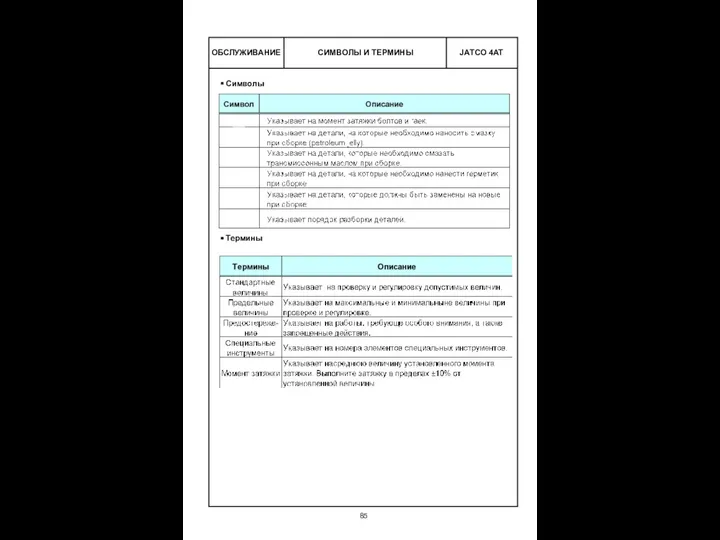 Коробка передач (4AT,JATCO). Составные части
Коробка передач (4AT,JATCO). Составные части Финансовая политика предприятия
Финансовая политика предприятия Презентация "Дом - жилище человека" - скачать презентации по МХК
Презентация "Дом - жилище человека" - скачать презентации по МХК Ремонтный процесс Standox Nissan KAB Ultimate Metal Silver. Тренинговая информация
Ремонтный процесс Standox Nissan KAB Ultimate Metal Silver. Тренинговая информация Монтаж и эксплуатация бурового оборудования. Основные определения и классификация. Лекция 1
Монтаж и эксплуатация бурового оборудования. Основные определения и классификация. Лекция 1 Презентация на тему "Валеологическая работа с детьми с туберкулезной интоксикацией" - скачать презентации по Медицине
Презентация на тему "Валеологическая работа с детьми с туберкулезной интоксикацией" - скачать презентации по Медицине Строительные конструкции. Еврокоды в строительной отрасли Казахстана. (Лекция 1)
Строительные конструкции. Еврокоды в строительной отрасли Казахстана. (Лекция 1) Одежда донских казаков Идем добывать зипуны… Казачья пословица
Одежда донских казаков Идем добывать зипуны… Казачья пословица  Презентация "Акира Куросава: Жизнь и творчество" - скачать презентации по МХК
Презентация "Акира Куросава: Жизнь и творчество" - скачать презентации по МХК Концепции космологии
Концепции космологии  Бунин Иван Алексеевич (1870 – 1953) Биография
Бунин Иван Алексеевич (1870 – 1953) Биография Политика. Представления о политике. Политика как подсистема общества
Политика. Представления о политике. Политика как подсистема общества Жанерке слайд
Жанерке слайд Электронные ресурсы в доступе СПбГУ
Электронные ресурсы в доступе СПбГУ Урок изобразительного искусства в 5 классе
Урок изобразительного искусства в 5 классе Simmetrical dsl technologies. Lecture 5
Simmetrical dsl technologies. Lecture 5 Расписание дня школьника Рыбаков Никита 5«Б» Класс
Расписание дня школьника Рыбаков Никита 5«Б» Класс  Профилактика травм опорно-двигательного аппарата в процессе учебно-тренировочных занятий у юных гандболистов 10-12 лет
Профилактика травм опорно-двигательного аппарата в процессе учебно-тренировочных занятий у юных гандболистов 10-12 лет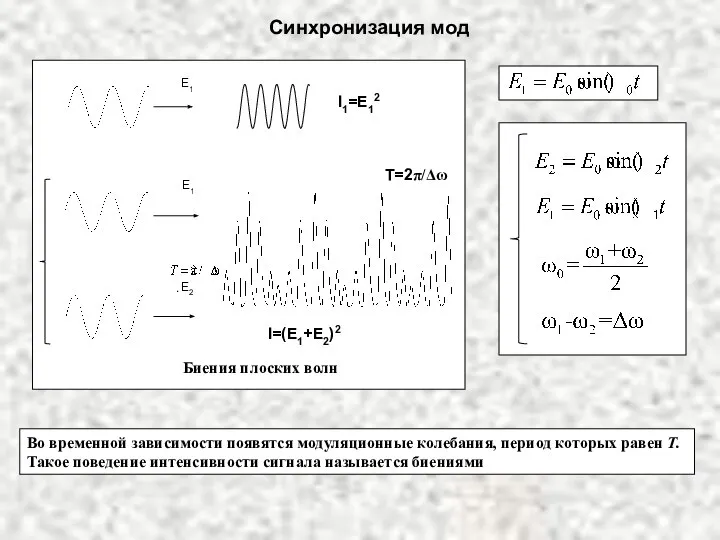 Синхронизация мод
Синхронизация мод  Ход реализации проекта «Оптимизация СХОД» Август 2010 г.
Ход реализации проекта «Оптимизация СХОД» Август 2010 г. Колонны. Расчет центрально-сжатых колонн по материалам
Колонны. Расчет центрально-сжатых колонн по материалам