- Адаптация, как результат эволюции

Содержание
- 2. Появление определенного генотипа на эволюционной арене и размножение его в определенных условиях – элементарное адаптационное явление
- 3. Адаптации – возникновение и развитие конкретных морфофизиологических свойств, значение которых для организма однозначно связано с теми
- 4. Классификация адаптаций по уровню организации биологических систем (Шкорбатов, 1971): онтогенетических систем - организмов, филогенетических систем -
- 5. С.С.Шварц - процесс видообразования, как процесс смены уровня адаптаций: развитие популяций в своеобразной среде → развитие
- 6. Классификация адаптаций (по Н.В.Тимофееву-Ресовскому)
- 7. Преадаптации (по Cuenot, 1914) Преадаптации – бесполезные или полуполезные особенности, способные превращаться в явные приспособления при
- 8. Чтобы иметь возможность успешно проникнуть в новую нишу или адаптивную зону, вид должен быть преадаптирован к
- 9. Комбинативные адаптации взаимодействие новых мутаций друг с другом и с генотипом. комплементация – усиление выраженности, эпистаз
- 10. they are considered the same species: Theridion grallator Популяционный полиморфизм и его роль в процессах адаптациогенеза
- 11. Термин “полиморфизм” используется только для обозначения прерывистой генетической изменчивости (Майр, 1974). Полиморфизм - сосуществование в более
- 12. Феногеография барсука (Meles meles) как пример, иллюстрирующий одновременно два крайних типа формообразования внутри вида: образование количественных
- 13. Гетерозиготный полиморфизм в лабораторной популяции Drosophila melanogaster по мутации ebony, устанавливающийся в результате некоторого повышения жизнеспособности
- 14. Сезонный полиморфизм
- 15. Сезонный полиморфизм
- 16. «Индустриальный меланизм»
- 19. Кора березы, покрытая лишайником Кора березы, покрытая копотью
- 20. Различная смертность особей березовой пяденицы (Bision betularia), выпущенных в разных лесистых местностях (Кеттлуэлл, 1961)
- 21. С.М.Гершензон Микроэволюция, полиморфизм и доминантные мутации // Природа. 1985. № 4. Гершензон С. М. «Мобилизационный резерв»
- 22. Характеристика мутантных генов, присутствующих в природных популяциях дрозофил (Гершензон, 1985)
- 23. Скорее всего, рецессивные мутации участвуют в эволюции тогда, когда адаптивную ценность приобретает редукция некоторых структур и
- 24. Первый этап: многократное возникновение доминантных, полудоминантных и кодоминантных мутаций в разных точках популяций, занимающих ареал вида
- 25. Генетический полиморфизм природных популяций — универсальное явление, составляющее основу генетической пластичности вида (Гершензон, 1983).
- 27. R.temporaria R.amurensis P.ridibundus R.arvalis
- 28. Frost et al., 2006
- 29. Pelophylax ridibundus Rana amurensis Rana temporaria Rana arvalis
- 30. Количество выделяемых морф
- 31. Встречаемость морфы striata у R.arvalis
- 32. Возрастные изменения во встречаемости морфы striata у R.arvalis
- 33. Встречаемость морфы striata у P.ridibundus
- 34. Возрастные изменения во встречаемости морфы striata у P.ridibundus
- 35. Средний возраст половозрелых R.arvalis
- 36. Натриевая проницаемость кожи R.arvalis
- 37. Уровень β-активности у R.arvalis c ВУРСа
- 38. ПОРОГ ВОЗБУДИМОСТИ НЕРВНОЙ ТКАНИ (R.arvalis)
- 39. КОНСТАНТА АККОМОДАЦИИ МЫШЕЧНОЙ ТКАНИ R.arvalis
- 40. Зависимость “длина – сила” (F=6,6; p=0,01)
- 41. Зависимость величины максимального изометрического напряжения миокарда R.arvalis от степени урбанизации (F=5,8; p=0,017).
- 42. Доля общих нейтрофилов P.ridibundus R.arvalis R.temporaria (F=5.1, p=0.03) (F=7.995, p=0.005)
- 43. Связь между количеством эритроцитов и индексом печени у R.temporaria
- 44. Связь между долей эритроидных предшественников и индексом печени у P.ridibundus
- 45. Видовые различия бесхвостых амфибий по содержанию общих лимфоцитов (%) у разных морф
- 46. Видовые различия бесхвостых амфибий по содержанию эозинофилов (%) у разных морф
- 47. Видовые различия бесхвостых амфибий по содержанию гранулоцитов (%) у разных морф
- 48. Видовые различия бесхвостых амфибий по содержанию эритроцитов (%) у разных морф
- 49. Видовые различия бесхвостых амфибий по содержанию гемоглобина у разных морф
- 50. Дистанцированность бесхвостых амфибий по изученным параметрам без учета морфы
- 51. Дистанцированность бесхвостых амфибий по изученным параметрам с учетом морфы
- 52. • Адаптивный потенциал во многом определяется наследственно обусловленными особенностями физиологии, спецификой структуры полиморфизма видов и популяций.
- 53. Широкое распространение генетических механизмов, которые обусловливают и поддерживают полиморфизм, непосредственно определяется отбором и само по себе
- 54. полиморфизм популяций - это универсальная стратегия, обеспечивающая сохранение целостности вида на основе постоянного взаимодействия наследственной изменчивости,
- 56. Скачать презентацию

Появление определенного генотипа на эволюционной арене и размножение его в определенных
Появление определенного генотипа на эволюционной арене и размножение его в определенных

Адаптации – возникновение и развитие конкретных морфофизиологических свойств, значение которых для
Адаптации – возникновение и развитие конкретных морфофизиологических свойств, значение которых для

Классификация адаптаций по уровню организации биологических систем (Шкорбатов, 1971):
онтогенетических систем
Классификация адаптаций по уровню организации биологических систем (Шкорбатов, 1971):
онтогенетических систем

С.С.Шварц - процесс видообразования, как процесс смены уровня адаптаций:
развитие популяций
С.С.Шварц - процесс видообразования, как процесс смены уровня адаптаций: развитие популяций

Классификация адаптаций
(по Н.В.Тимофееву-Ресовскому)
Классификация адаптаций
(по Н.В.Тимофееву-Ресовскому)

Преадаптации
(по Cuenot, 1914)
Преадаптации – бесполезные или полуполезные особенности, способные превращаться
Преадаптации
(по Cuenot, 1914)
Преадаптации – бесполезные или полуполезные особенности, способные превращаться

Чтобы иметь возможность успешно проникнуть в новую нишу или адаптивную
Чтобы иметь возможность успешно проникнуть в новую нишу или адаптивную

Комбинативные адаптации
взаимодействие новых мутаций друг с другом и с генотипом.
комплементация
Комбинативные адаптации
взаимодействие новых мутаций друг с другом и с генотипом.
комплементация

they are considered the same species: Theridion grallator
Популяционный полиморфизм
и его
they are considered the same species: Theridion grallator
Популяционный полиморфизм и его

Термин “полиморфизм” используется только для обозначения прерывистой генетической изменчивости (Майр, 1974).
Полиморфизм
Термин “полиморфизм” используется только для обозначения прерывистой генетической изменчивости (Майр, 1974).
Полиморфизм
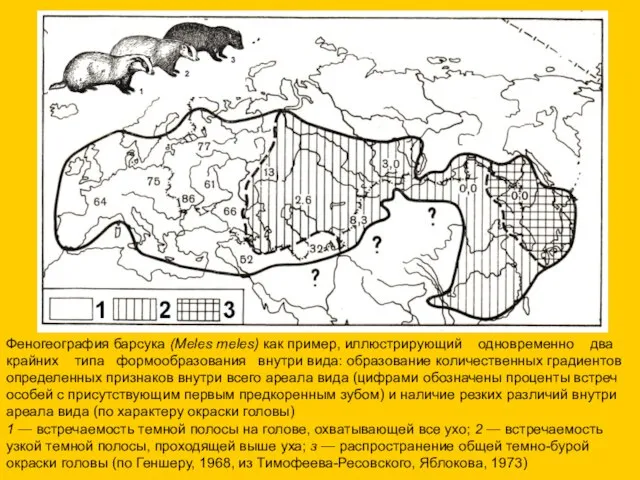
Феногеография барсука (Meles meles) как пример, иллюстрирующий одновременно два крайних типа
Феногеография барсука (Meles meles) как пример, иллюстрирующий одновременно два крайних типа
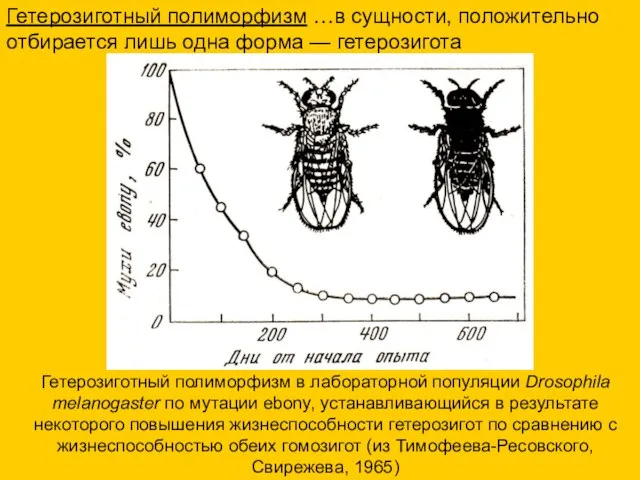
Гетерозиготный полиморфизм в лабораторной популяции Drosophila melanogaster по мутации ebony, устанавливающийся
Гетерозиготный полиморфизм в лабораторной популяции Drosophila melanogaster по мутации ebony, устанавливающийся
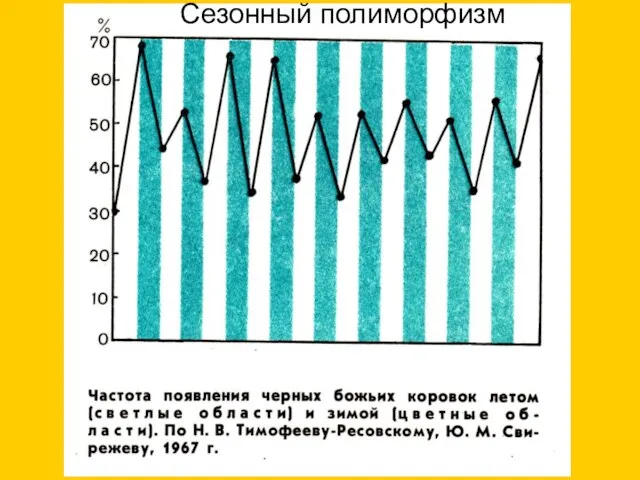
Сезонный полиморфизм
Сезонный полиморфизм
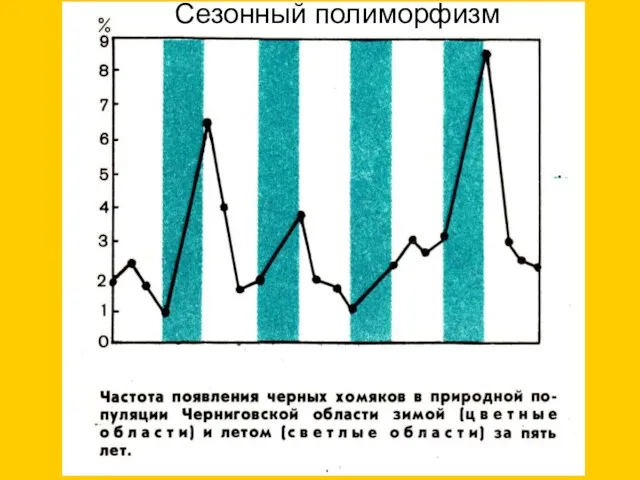
Сезонный полиморфизм
Сезонный полиморфизм
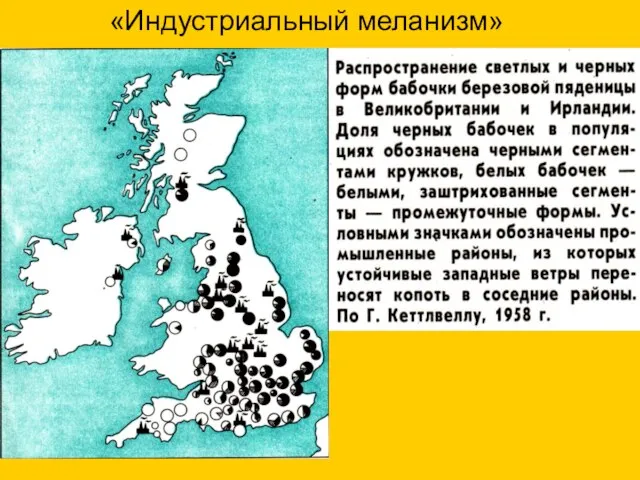
«Индустриальный меланизм»
«Индустриальный меланизм»



Кора березы, покрытая лишайником
Кора березы, покрытая копотью
Кора березы, покрытая лишайником
Кора березы, покрытая копотью
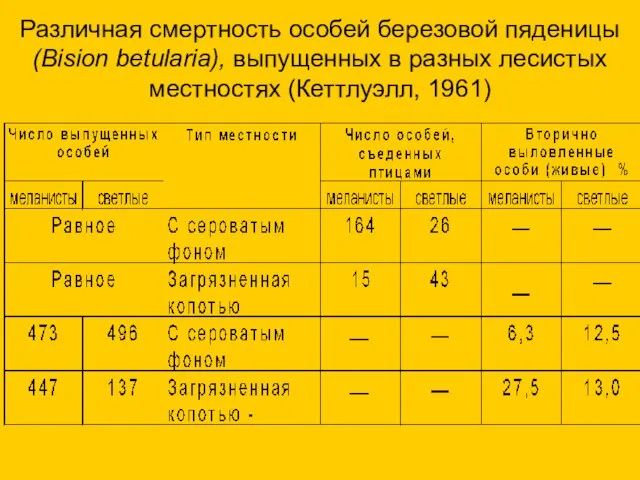
Различная смертность особей березовой пяденицы (Bision betularia), выпущенных в разных лесистых
Различная смертность особей березовой пяденицы (Bision betularia), выпущенных в разных лесистых

С.М.Гершензон Микроэволюция, полиморфизм и доминантные мутации // Природа. 1985. № 4.
Гершензон
С.М.Гершензон Микроэволюция, полиморфизм и доминантные мутации // Природа. 1985. № 4. Гершензон

Характеристика мутантных генов, присутствующих в природных популяциях дрозофил (Гершензон, 1985)
Характеристика мутантных генов, присутствующих в природных популяциях дрозофил (Гершензон, 1985)

Скорее всего, рецессивные мутации участвуют в эволюции тогда, когда адаптивную
Скорее всего, рецессивные мутации участвуют в эволюции тогда, когда адаптивную

Первый этап: многократное возникновение доминантных, полудоминантных и кодоминантных мутаций в
Первый этап: многократное возникновение доминантных, полудоминантных и кодоминантных мутаций в

Генетический полиморфизм природных популяций — универсальное явление, составляющее основу генетической пластичности
Генетический полиморфизм природных популяций — универсальное явление, составляющее основу генетической пластичности

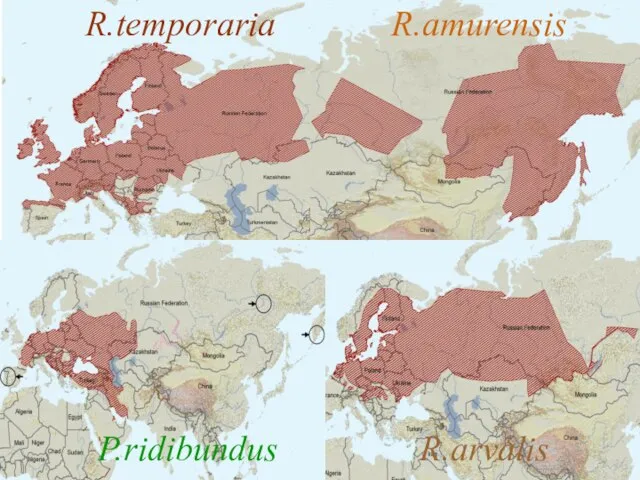
R.temporaria R.amurensis
P.ridibundus R.arvalis
R.temporaria R.amurensis
P.ridibundus R.arvalis
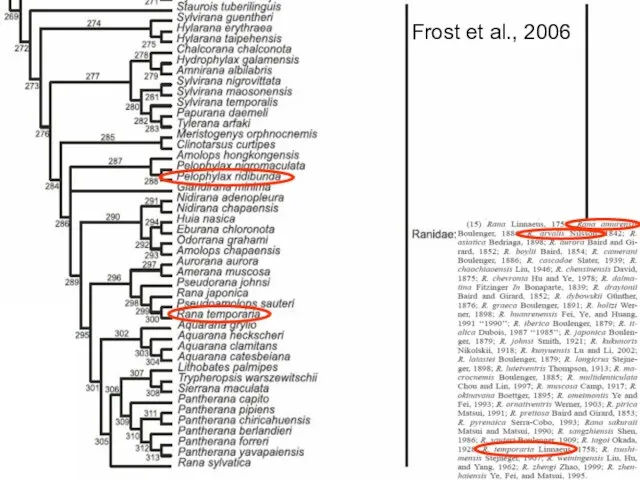
Frost et al., 2006
Frost et al., 2006
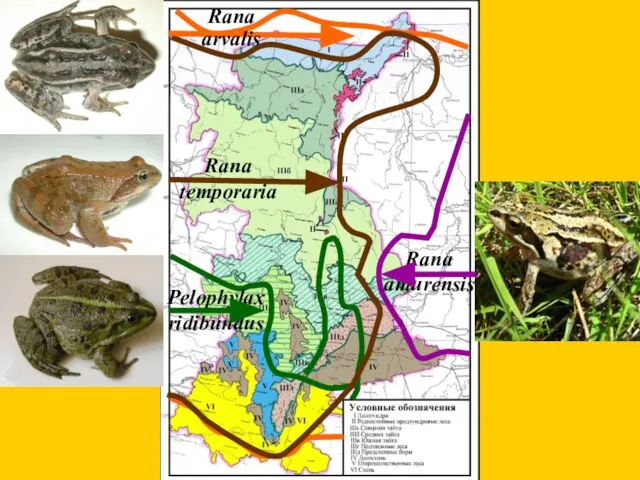
Pelophylax ridibundus
Rana amurensis
Rana temporaria
Rana arvalis
Pelophylax ridibundus
Rana amurensis
Rana temporaria
Rana arvalis
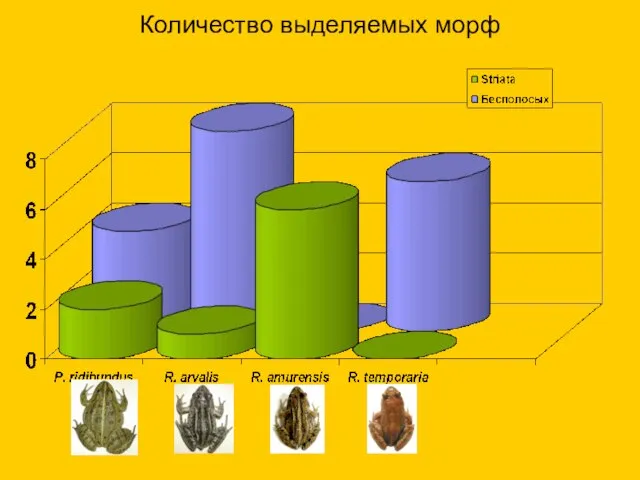
Количество выделяемых морф
Количество выделяемых морф
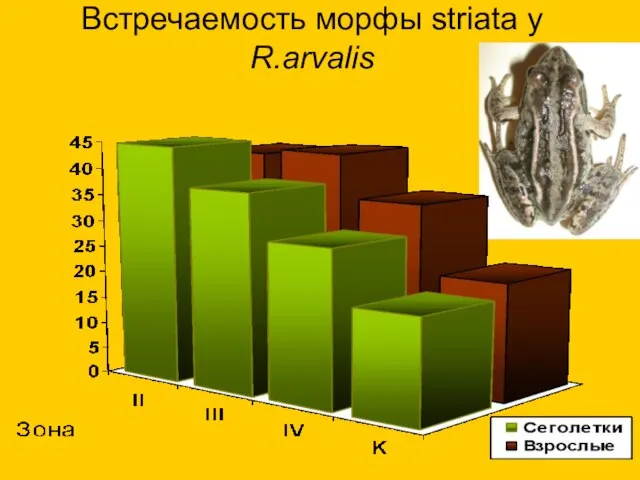
Встречаемость морфы striata у R.arvalis
Встречаемость морфы striata у R.arvalis
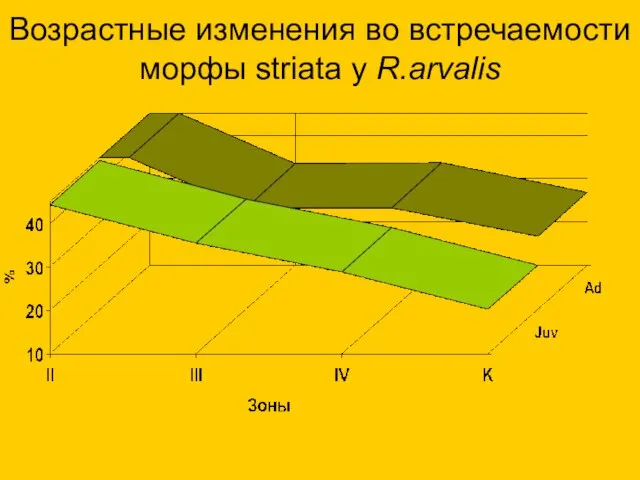
Возрастные изменения во встречаемости морфы striata у R.arvalis
Возрастные изменения во встречаемости морфы striata у R.arvalis

Встречаемость морфы striata у P.ridibundus
Встречаемость морфы striata у P.ridibundus
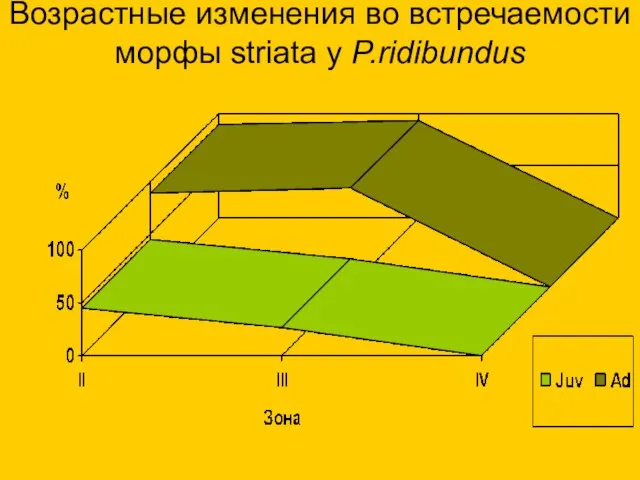
Возрастные изменения во встречаемости морфы striata у P.ridibundus
Возрастные изменения во встречаемости морфы striata у P.ridibundus

Средний возраст половозрелых R.arvalis
Средний возраст половозрелых R.arvalis

Натриевая проницаемость кожи R.arvalis
Натриевая проницаемость кожи R.arvalis

Уровень β-активности у R.arvalis c ВУРСа
Уровень β-активности у R.arvalis c ВУРСа

ПОРОГ ВОЗБУДИМОСТИ НЕРВНОЙ ТКАНИ (R.arvalis)
ПОРОГ ВОЗБУДИМОСТИ НЕРВНОЙ ТКАНИ (R.arvalis)

КОНСТАНТА АККОМОДАЦИИ МЫШЕЧНОЙ ТКАНИ R.arvalis
КОНСТАНТА АККОМОДАЦИИ МЫШЕЧНОЙ ТКАНИ R.arvalis
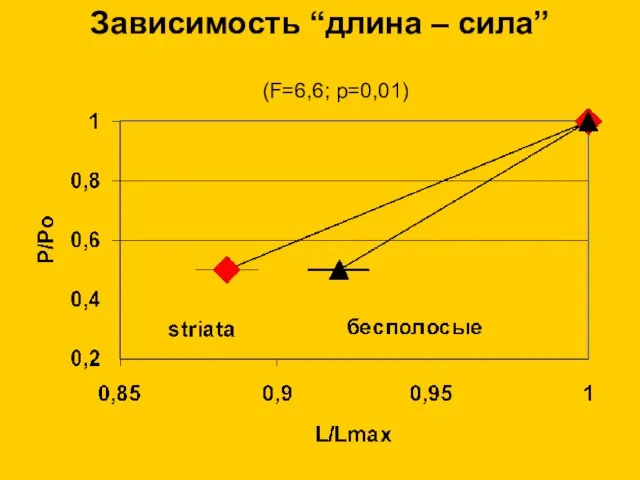
Зависимость “длина – сила”
(F=6,6; p=0,01)
Зависимость “длина – сила”
(F=6,6; p=0,01)
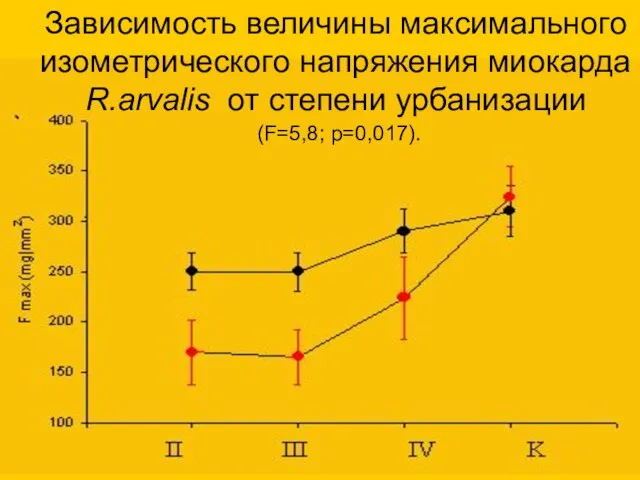
Зависимость величины максимального изометрического напряжения миокарда R.arvalis от степени урбанизации
(F=5,8; p=0,017).
Зависимость величины максимального изометрического напряжения миокарда R.arvalis от степени урбанизации
(F=5,8; p=0,017).

Доля общих нейтрофилов
P.ridibundus R.arvalis R.temporaria
(F=5.1, p=0.03)
(F=7.995, p=0.005)
Доля общих нейтрофилов
P.ridibundus R.arvalis R.temporaria
(F=5.1, p=0.03)
(F=7.995, p=0.005)
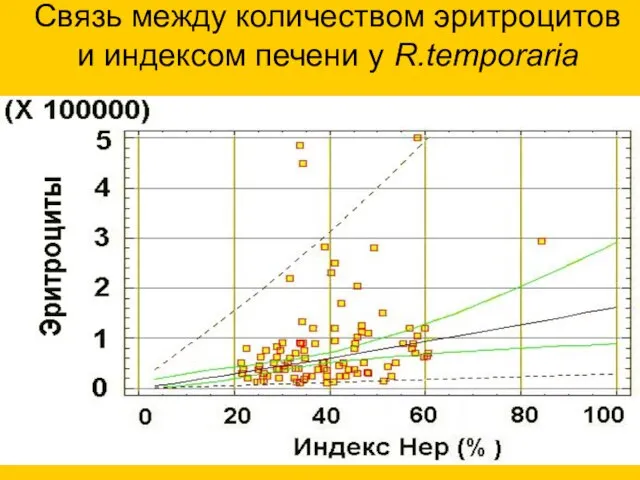
Связь между количеством эритроцитов
и индексом печени у R.temporaria
Связь между количеством эритроцитов
и индексом печени у R.temporaria
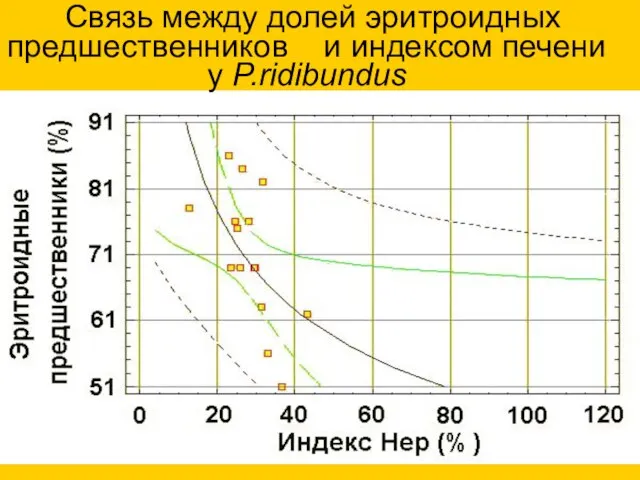
Связь между долей эритроидных предшественников и индексом печени
у P.ridibundus
Связь между долей эритроидных предшественников и индексом печени
у P.ridibundus

Видовые различия бесхвостых амфибий по содержанию общих лимфоцитов (%) у разных
Видовые различия бесхвостых амфибий по содержанию общих лимфоцитов (%) у разных

Видовые различия бесхвостых амфибий по содержанию эозинофилов (%) у разных морф
Видовые различия бесхвостых амфибий по содержанию эозинофилов (%) у разных морф

Видовые различия бесхвостых амфибий по содержанию гранулоцитов (%) у разных морф
Видовые различия бесхвостых амфибий по содержанию гранулоцитов (%) у разных морф

Видовые различия бесхвостых амфибий по содержанию эритроцитов (%) у разных морф
Видовые различия бесхвостых амфибий по содержанию эритроцитов (%) у разных морф

Видовые различия бесхвостых амфибий по содержанию гемоглобина у разных морф
Видовые различия бесхвостых амфибий по содержанию гемоглобина у разных морф

Дистанцированность бесхвостых амфибий по изученным параметрам без учета морфы
Дистанцированность бесхвостых амфибий по изученным параметрам без учета морфы

Дистанцированность бесхвостых амфибий по изученным параметрам с учетом морфы
Дистанцированность бесхвостых амфибий по изученным параметрам с учетом морфы

• Адаптивный потенциал во многом определяется наследственно обусловленными особенностями физиологии, спецификой
• Адаптивный потенциал во многом определяется наследственно обусловленными особенностями физиологии, спецификой

Широкое распространение генетических механизмов, которые обусловливают и поддерживают полиморфизм, непосредственно определяется
Широкое распространение генетических механизмов, которые обусловливают и поддерживают полиморфизм, непосредственно определяется

полиморфизм популяций - это универсальная стратегия, обеспечивающая сохранение целостности вида на
полиморфизм популяций - это универсальная стратегия, обеспечивающая сохранение целостности вида на
 Генетика популяций
Генетика популяций Презентация на тему "Корень.Строение и функции.ВУидоизменения корня." - скачать презентации по Биологии
Презентация на тему "Корень.Строение и функции.ВУидоизменения корня." - скачать презентации по Биологии Дикие животные ранней весной
Дикие животные ранней весной Учение о тканях. Виды тканей
Учение о тканях. Виды тканей Теория Кибби
Теория Кибби Вирусы. Дмитрий Иосифович Ивановский
Вирусы. Дмитрий Иосифович Ивановский Презентация на тему "Молочнокислые бактерии" - скачать бесплатно презентации по Биологии
Презентация на тему "Молочнокислые бактерии" - скачать бесплатно презентации по Биологии Черви. Классификация
Черви. Классификация Строение организма человека
Строение организма человека Эксперимент в космосе Исследование фотосинтеза водорослей в условиях невесомости. Выполнила ученица МОУ 8б «Бахтемирская СО
Эксперимент в космосе Исследование фотосинтеза водорослей в условиях невесомости. Выполнила ученица МОУ 8б «Бахтемирская СО Сердце и кровообращение
Сердце и кровообращение Вегетативная (автономная) нервная система. Глава II
Вегетативная (автономная) нервная система. Глава II Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз
Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз Путешествие в царство грибов
Путешествие в царство грибов Презентация на тему "Тип Членистоногие. Класс Паукообразные" - скачать презентации по Биологии
Презентация на тему "Тип Членистоногие. Класс Паукообразные" - скачать презентации по Биологии Презентация на тему "Белый медведь - полярный странник" - скачать презентации по Биологии
Презентация на тему "Белый медведь - полярный странник" - скачать презентации по Биологии Современная зондовая микроскопия
Современная зондовая микроскопия Презентация на тему Трофическая структура БГЦ
Презентация на тему Трофическая структура БГЦ  Регуляция пищеварения
Регуляция пищеварения Презентация на тему Свойства экосистем. Смена экосистем
Презентация на тему Свойства экосистем. Смена экосистем  Цитоплазма бактерий
Цитоплазма бактерий Презентация на тему "Морской конек" - скачать презентации по Биологии
Презентация на тему "Морской конек" - скачать презентации по Биологии Сити-фермер, космобиолог
Сити-фермер, космобиолог Опорно-двигательная система, филогенез
Опорно-двигательная система, филогенез Презентация на тему "Семейство маревые" - скачать презентации по Биологии
Презентация на тему "Семейство маревые" - скачать презентации по Биологии Четвертичный период
Четвертичный период Нервная система. Клеточное строение
Нервная система. Клеточное строение М’язові тканини
М’язові тканини