- Физиология высшей нервной деятельности. Физиология сна и бодрствования. Активирующие системы мозга

Содержание
- 2. Суточные ритмы
- 3. Мориц Эшер. День и ночь
- 4. Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира.
- 5. Супрахиазматическое ядро Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной
- 6. Структура сна. Ритмы ЭЭГ при различных стадиях сна и при бодрствовании.
- 7. Выделяют две основные стадии сна: — медленноволновый сон и — парадоксальный сон, или сон с быстрыми
- 8. Феномен «быстрого» сна открыли в 1953 г. американские ученые А.Азеринский и К. Клейтман. Медленноволновой сон человека
- 9. На протяжении ночи глубина сна волнообразно увеличивается и уменьшается, а периоды БДГ-сна постепенно удлиняются. Переход между
- 10. График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального
- 11. ЭЭГ человека при бодрствовании и сне
- 12. В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне.
- 13. Основные ритмы ЭЭГ человека
- 14. ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды
- 15. Физиологические механизмы сна
- 16. Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки
- 17. Дж.Моруцци и Х.Мегун показали в 1949 г., что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон
- 18. В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря
- 19. Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна
- 20. Ацетилхолин, норадреналин, гистамин, серотонин, гистамин и дофамин – все эти медиаторы необходимы для поддержания бодрствующего и
- 21. В настоящее время очевидно, что бодрствование и два вида сна – качественно различные состояния, каждое из
- 22. Все центры, обеспечивающие состояние бодрствования, координируются из одного общего центра в гипоталамусе, который выделяет орексин. Функция
- 23. Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным).
- 24. Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор — гамма-аминомасляная
- 25. Нарушения сна
- 26. Группы нарушений сна человека: Инсомнии (бессонница) 2. Гиперсомнии (сонливость, нарколепсия) 3. Парасомнии - двигательные: сомнамбулизм (лунатизм),
- 32. Скачать презентацию

Суточные ритмы
Суточные ритмы

Мориц Эшер. День и ночь
Мориц Эшер. День и ночь
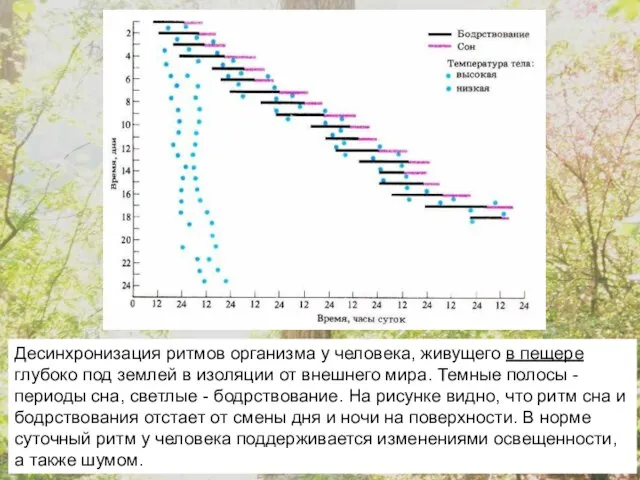
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей
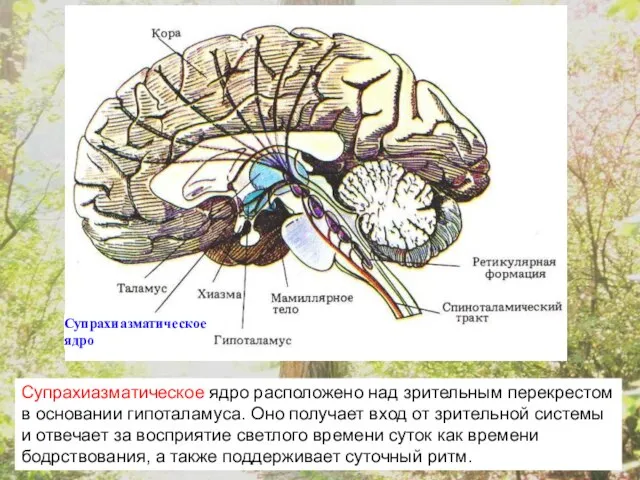
Супрахиазматическое ядро
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно
Супрахиазматическое ядро
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно

Структура сна. Ритмы ЭЭГ при различных стадиях сна и при бодрствовании.
Структура сна. Ритмы ЭЭГ при различных стадиях сна и при бодрствовании.

Выделяют две основные стадии сна:
— медленноволновый сон и
— парадоксальный сон,
Выделяют две основные стадии сна:
— медленноволновый сон и
— парадоксальный сон,

Феномен «быстрого» сна открыли в 1953 г. американские ученые А.Азеринский и
Феномен «быстрого» сна открыли в 1953 г. американские ученые А.Азеринский и
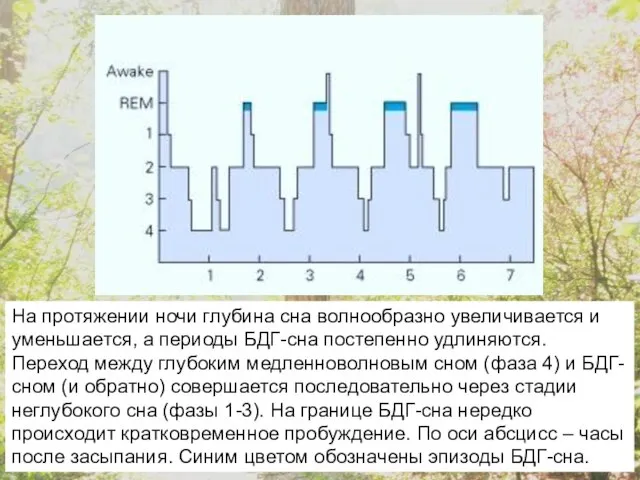
На протяжении ночи глубина сна волнообразно увеличивается и уменьшается, а периоды
На протяжении ночи глубина сна волнообразно увеличивается и уменьшается, а периоды
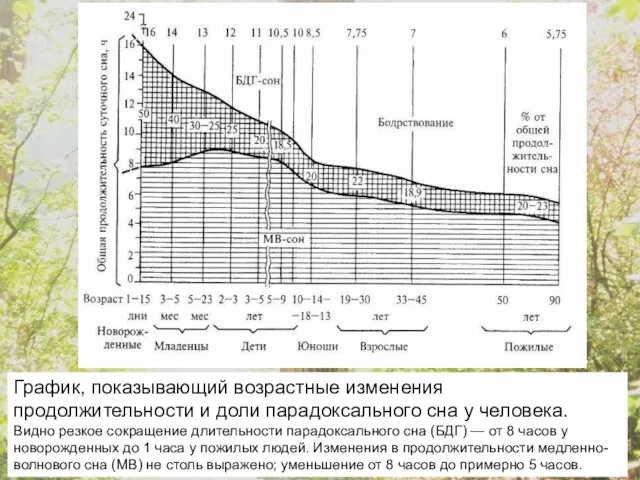
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека.
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека.
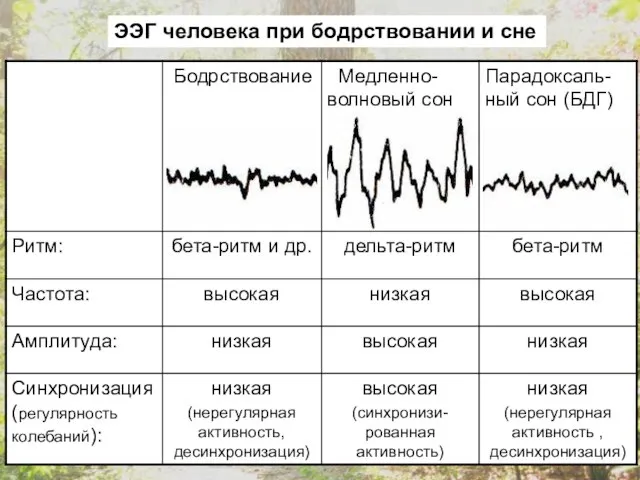
ЭЭГ человека при бодрствовании и сне
ЭЭГ человека при бодрствовании и сне

В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же
В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же
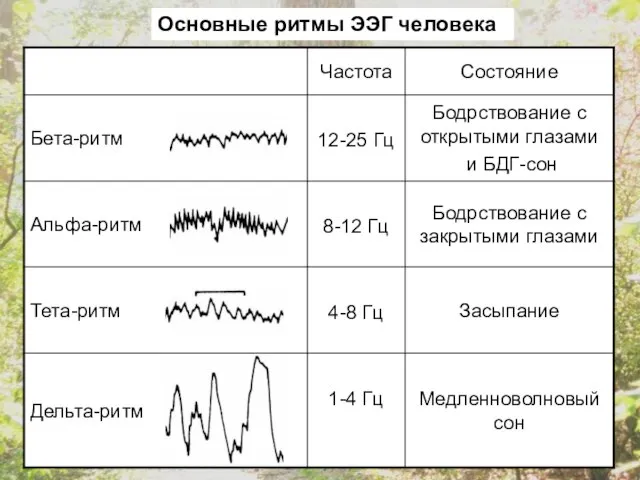
Основные ритмы ЭЭГ человека
Основные ритмы ЭЭГ человека
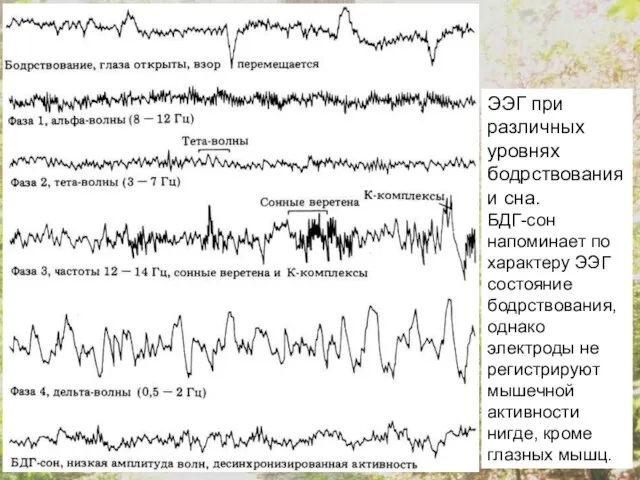
ЭЭГ при различных уровнях бодрствования и сна.
БДГ-сон напоминает по характеру
ЭЭГ при различных уровнях бодрствования и сна.
БДГ-сон напоминает по характеру

Физиологические механизмы сна
Физиологические механизмы сна
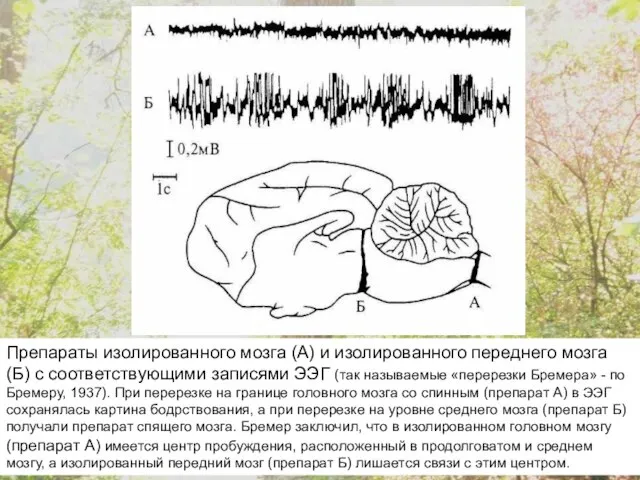
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими

Дж.Моруцци и Х.Мегун показали в 1949 г., что стимуляция ретикулярной формации
Дж.Моруцци и Х.Мегун показали в 1949 г., что стимуляция ретикулярной формации

В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации»
В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации»
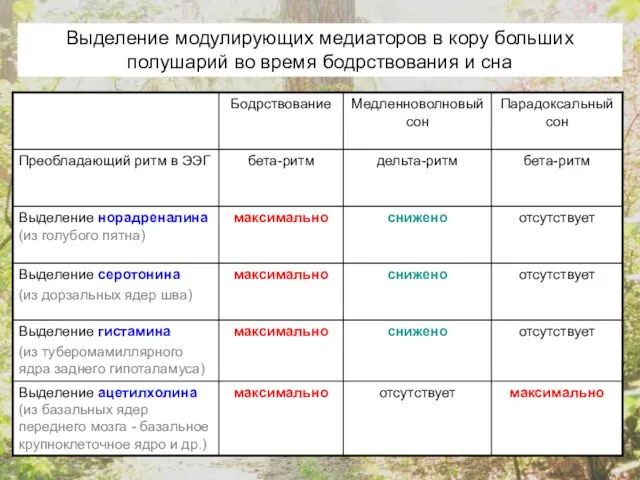
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и

Ацетилхолин, норадреналин, гистамин, серотонин, гистамин и дофамин – все эти медиаторы
Ацетилхолин, норадреналин, гистамин, серотонин, гистамин и дофамин – все эти медиаторы

В настоящее время очевидно, что бодрствование и два вида сна –
В настоящее время очевидно, что бодрствование и два вида сна –

Все центры, обеспечивающие состояние бодрствования, координируются из одного общего центра в
Все центры, обеспечивающие состояние бодрствования, координируются из одного общего центра в
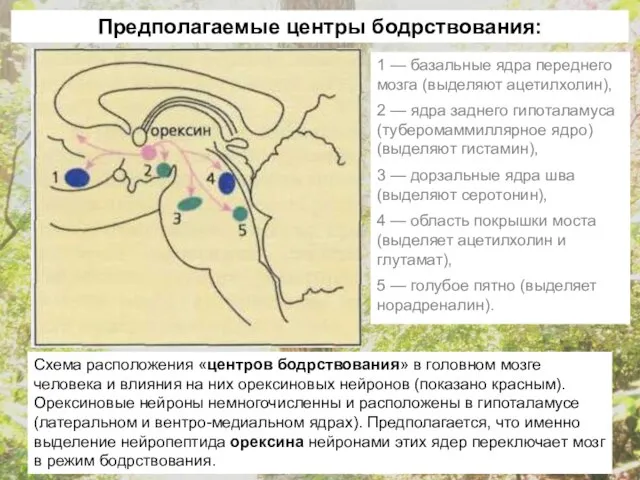
Схема расположения «центров бодрствования» в головном мозге человека и влияния на
Схема расположения «центров бодрствования» в головном мозге человека и влияния на

Предполагаемые центры сна:
Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом
Предполагаемые центры сна:
Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом

Нарушения сна
Нарушения сна

Группы нарушений сна человека:
Инсомнии (бессонница)
2. Гиперсомнии (сонливость, нарколепсия)
3. Парасомнии
- двигательные:
Группы нарушений сна человека:
Инсомнии (бессонница)
2. Гиперсомнии (сонливость, нарколепсия)
3. Парасомнии
- двигательные:




 Птицы. Выполнила ученица 7 класса: Пашко Екатерина. Учитель: Волчкова Л.К
Птицы. Выполнила ученица 7 класса: Пашко Екатерина. Учитель: Волчкова Л.К  Природная зональность
Природная зональность Ученицы 6 класса “Д” Атаянц Насти
Ученицы 6 класса “Д” Атаянц Насти Подцарство Настоящие бактерии, Архебактерии
Подцарство Настоящие бактерии, Архебактерии Презентация на тему Развитие жизни в палеозойской эре Ранний палеозой (кембрий, ордовик, силур)
Презентация на тему Развитие жизни в палеозойской эре Ранний палеозой (кембрий, ордовик, силур)  клетка 8 кл
клетка 8 кл Микропрепараты к итоговому занятию по разделу Ткани
Микропрепараты к итоговому занятию по разделу Ткани Сон. Фізіологія сну
Сон. Фізіологія сну Антагонизмдік қарым-қатынастар
Антагонизмдік қарым-қатынастар Проверка домашнего задания: Верно ли утверждение? Членистоногие –самый крупный по количеству видов тип животных. К членистон
Проверка домашнего задания: Верно ли утверждение? Членистоногие –самый крупный по количеству видов тип животных. К членистон Череп человека
Череп человека Третий этап энергетического обмена
Третий этап энергетического обмена Оригинальная технология эффективных микроорганизмов в России
Оригинальная технология эффективных микроорганизмов в России История открытия микроорганизмов
История открытия микроорганизмов Адаптации организмов к условиям окружающей среды
Адаптации организмов к условиям окружающей среды Презентация на тему Среды обитания организмов
Презентация на тему Среды обитания организмов  Органы кровеносной и лимфатической систем. Круги кровообращения
Органы кровеносной и лимфатической систем. Круги кровообращения КЛЕТКА -ОСНОВА СТРОЕНИЯ И РОСТА ЖИВЫХ ОРГАНИЗМОВ
КЛЕТКА -ОСНОВА СТРОЕНИЯ И РОСТА ЖИВЫХ ОРГАНИЗМОВ  Тип Иглокожие. Особенности строения и жизнедеятельности
Тип Иглокожие. Особенности строения и жизнедеятельности Презентация на тему Мезозойская эра
Презентация на тему Мезозойская эра Кто такие насекомые
Кто такие насекомые Использование микробиологии в промышленности, добыче минеральных ресурсов
Использование микробиологии в промышленности, добыче минеральных ресурсов Папоротники, хвощи, плауны
Папоротники, хвощи, плауны Ткани животных
Ткани животных Полезные и забавные растения
Полезные и забавные растения Сукцессия. Классификации сукцессий
Сукцессия. Классификации сукцессий Презентация на тему Современные методы селекции
Презентация на тему Современные методы селекции  Своя игра Занимательная биология
Своя игра Занимательная биология