- Механизмы сокращения скелетных мышц

Содержание
- 2. Сравнение динамики сокращения разных типов мышечной ткани Типы мышечной ткани Гладкая Сердечная Скелетная
- 3. Уровни организации скелетной мышцы Н.Б.Гусев Соросовский образовательный журнал 2000, том 6, №8, с.24-32 Миофибрилла Волокна (клетки)
- 4. Саркомер – единица строения и работы миофибриллы Z-линии разных миофибрилл расположены на одном уровне
- 5. Белки саркомера α-актинин Миозин Титин (тайтин) Актин Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
- 6. 1939 г.: В.А. Энгельгардт и М.Н.Любимова установили, что миозин обладает АТФ-азной активностью (работа «Ферментативные свойства миозина
- 7. Толстые филаменты состоят из миозина II типа Молекула миозина Хвост Легкие цепи: пара щелочных (17 кДа)
- 8. Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55 Связывание Са2+ с тропонином С
- 9. Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55 Связывание Са2+ с тропонином С
- 10. Теория скольжения нитей: во время сокращения мышцы длина толстых и тонких филаментов не изменяется, происходит их
- 11. Цикл работы головки миозина В отсуттствие АТФ развивается ригор («трупное окоченение»)
- 12. Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых и миозиновых филаментов)
- 13. Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых и миозиновых филаментов)
- 14. Основные пути Са2+-зависимой активации актомиозинового взаимодействия Актин = «рельсы» Миозиновый мотор КПД 50-80% (у двигателей внутреннего
- 15. Взаимодействие головок миозина с актином возможно лишь при повышении концентрации Са2+ в цитоплазме Зависимость между концентрацией
- 17. 1. Потенциал действия 3. Связывание Са2+ c тропонином 4. Сокращение 2. Повышение внутриклеточной концентрации Ca2+ Электромеханическое
- 18. Потенциал действия распространяется вглубь мышечного волокна по поперечным трубочкам (Т-трубочки) и запускает высвобождение Са2+ из саркоплазматического
- 19. Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
- 20. Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР) Три гипотезы о механизме передачи сигнала от Т-трубочек к
- 21. Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР: Chandler, Rakowski and Schneider (1976)
- 22. Соединительные ножки между мембранами Т-трубочки и СПР
- 23. «Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-управляемый кальциевый канал L-типа) А.М.Рубцов Соросовский образовательный журнал 2000, том
- 24. Классификация потенциал-зависимых Са2+-каналов
- 25. Ryania speciosa Трехмерная модель РиР, построенная по данным электронной микроскопии Вид сбоку Вид со стороны цитоплазмы
- 26. Основные этапы электромеханического сопряжения в скелетной мышце
- 27. А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24 Активация рианодиновых рецепторов Типы рианодиновых рецепторов: cкелетная
- 28. Разные мышцы человека существенно различаются по динамике одиночного сокращения
- 29. Типы мышечных волокон у позвоночных Не генерируют ПД Имеют полисинаптическую иннервацию Отвечают на нервное воздействие градуальным
- 30. Поперечные серийные срезы четырехглавой мышцы бедра человека, окрашенные антителами против медленных (слева) и быстрых (справа) изоформ
- 31. Toniolo et al. Am J Physiol. 2007 292(5):C1915-26. In vitro motility system
- 32. Prochniewicza et al. Exp.Gerontol 2007. V.42, p.931–938 Toniolo et al. Am J Physiol. 2007 292(5):C1915-26. In
- 33. Источники АТФ в скелетной мышце
- 34. Типы мышечных волокон у позвоночных Не генерируют ПД Имеют полисинаптическую иннервацию Отвечают на нервное воздействие градуальным
- 35. Гликолитические и оксидативные мышечные волокна различаются по размерам и активности митохондриальных ферментов Гистохимическое окрашивание капилляров Гистохимическое
- 36. Гликолитические и оксидативные волокна различаются по устойчивости к утомлению
- 37. Свойства различных типов фазических мышечных волокон
- 38. ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы (а не торможение сокращения) Нервно-мышечный синапс обладает
- 39. Двигательные (моторные) единицы Двигательная единица = мотонейрон + группа иннервируемых им мышечных волокон Все мышечные волокна
- 40. Размер и количество ДЕ в мышцах человека сильно различаются (в зависимости от функций этих мышц и
- 41. Два способа увеличения силы сокращения мышцы: Активация большего числа мотонейронов (рекрутирование ДЕ) Увеличение частоты разрядов мотонейронов:
- 42. Метаболический тип мышечного волокна определяется его иннервацией (характером разрядной активности мотонейрона) Изменение свойств мышц при перекрестной
- 43. Изменения мышц при естественной активности мотонейронов Тренировка силы (тяжелая атлетика) Тренировка выносливости (марафон) Как выглядят тела
- 44. В здоровом теле – здоровый дух!
- 45. Спасибо за внимание!
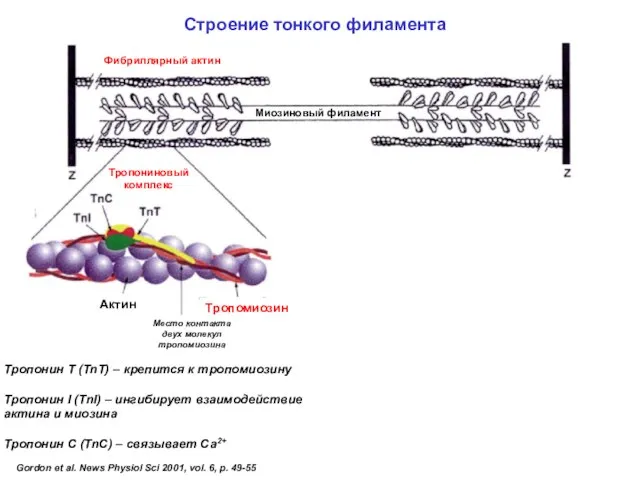
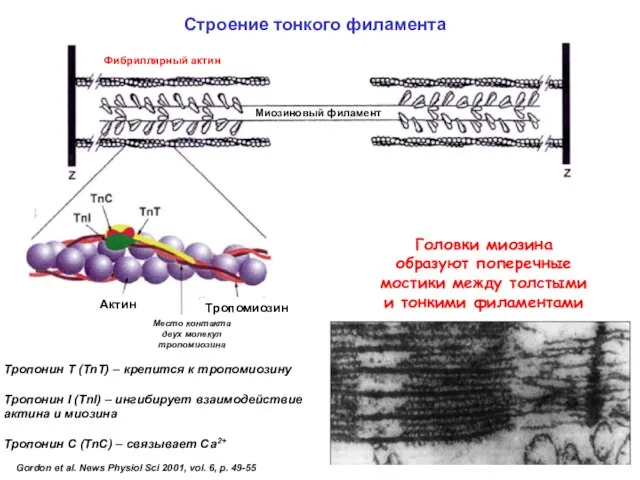
- 46. Строение тонкого филамента
- 47. Разные типы мышечных волокон в икроножной мышце крысы А и В – окрашивание антителами к разным
- 48. Быстрые и медленные мышечные волокна различаются по активности ферментов окислительного фосфорилирования и гликолиза А и В
- 49. Строение толстого филамента (состоит из миозина II типа)
- 51. Скачать презентацию
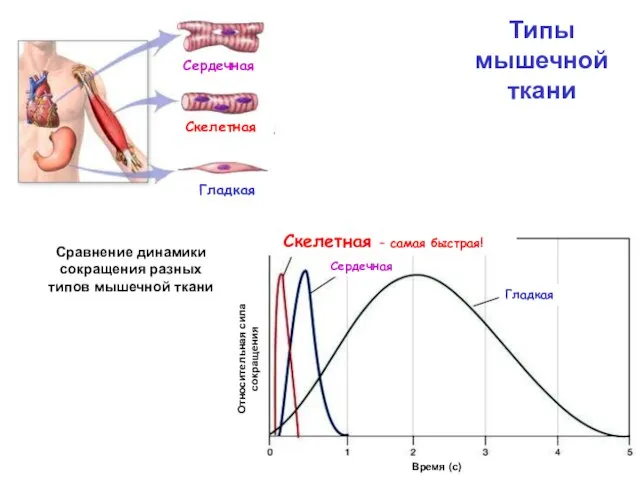
Сравнение динамики сокращения разных типов мышечной ткани
Типы мышечной ткани
Гладкая
Сравнение динамики сокращения разных типов мышечной ткани
Типы мышечной ткани
Гладкая
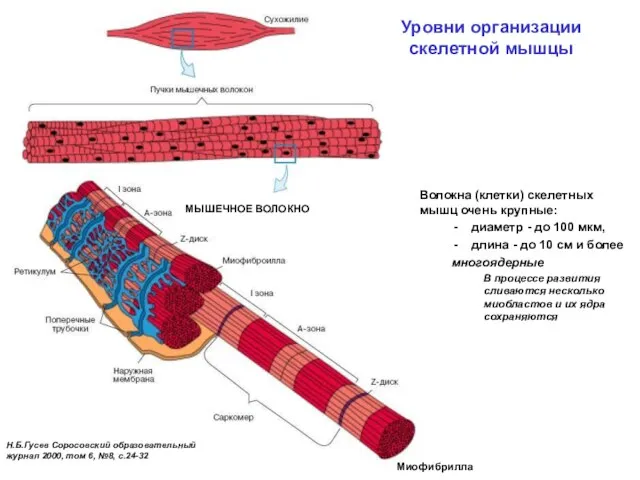
Уровни организации
скелетной мышцы
Н.Б.Гусев Соросовский образовательный
журнал 2000, том 6, №8,
Уровни организации
скелетной мышцы
Н.Б.Гусев Соросовский образовательный
журнал 2000, том 6, №8,
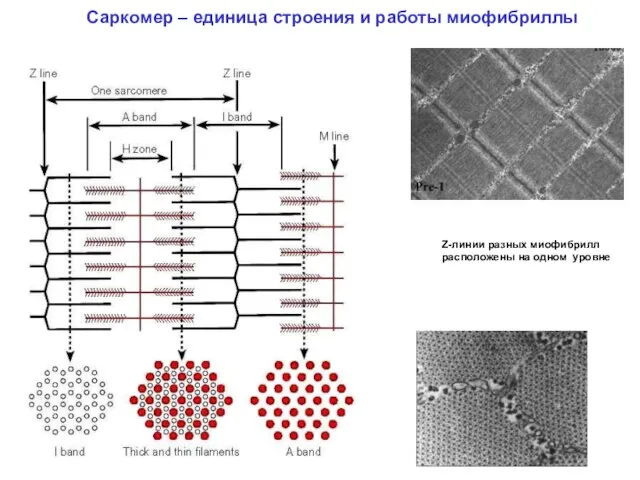
Саркомер – единица строения и работы миофибриллы
Z-линии разных миофибрилл расположены
Саркомер – единица строения и работы миофибриллы
Z-линии разных миофибрилл расположены
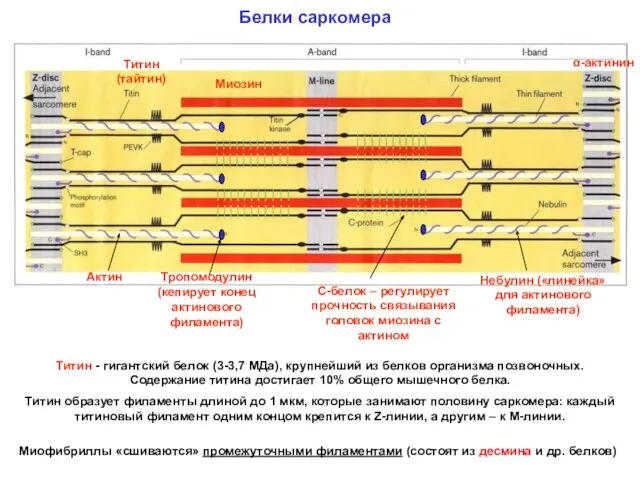
Белки саркомера
α-актинин
Миозин
Титин (тайтин)
Актин
Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
Белки саркомера
α-актинин
Миозин
Титин (тайтин)
Актин
Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
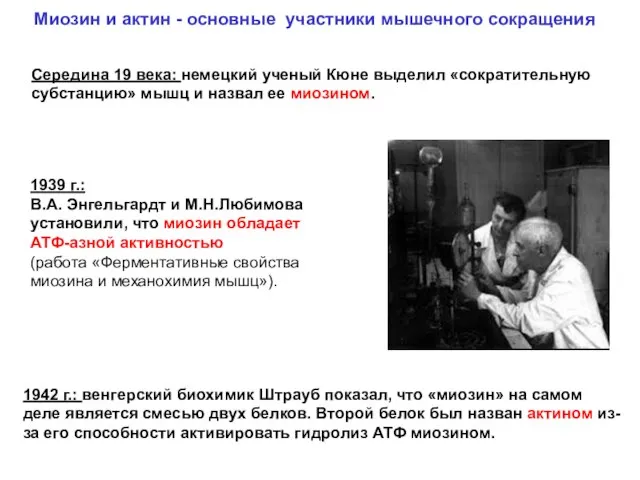
1939 г.:
В.А. Энгельгардт и М.Н.Любимова установили, что миозин обладает АТФ-азной
1939 г.:
В.А. Энгельгардт и М.Н.Любимова установили, что миозин обладает АТФ-азной
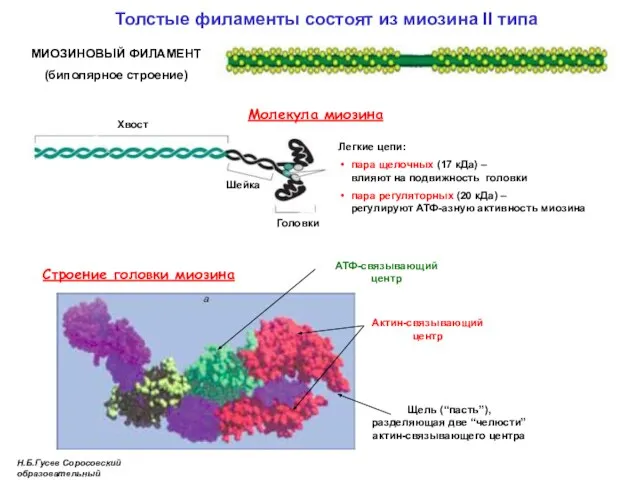
Толстые филаменты состоят из миозина II типа
Молекула миозина
Хвост
Легкие цепи:
пара щелочных
Толстые филаменты состоят из миозина II типа
Молекула миозина
Хвост
Легкие цепи:
пара щелочных
Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55
Связывание
Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55
Связывание
Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55
Связывание
Gordon et al. News Physiol Sci 2001, vol. 6, p. 49-55
Связывание

Теория скольжения нитей:
во время сокращения мышцы длина толстых и тонких
Теория скольжения нитей:
во время сокращения мышцы длина толстых и тонких
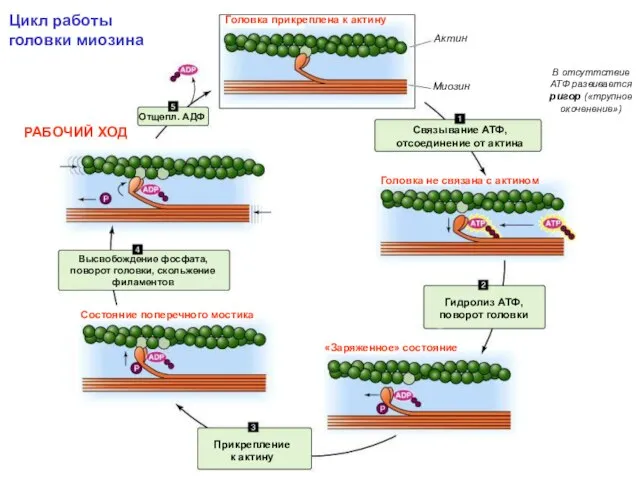
Цикл работы головки миозина
В отсуттствие АТФ развивается ригор («трупное окоченение»)
Цикл работы головки миозина
В отсуттствие АТФ развивается ригор («трупное окоченение»)
Сила сокращения скелетной мышцы зависит от длины саркомеров
(степени перекрывания актиновых
Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых
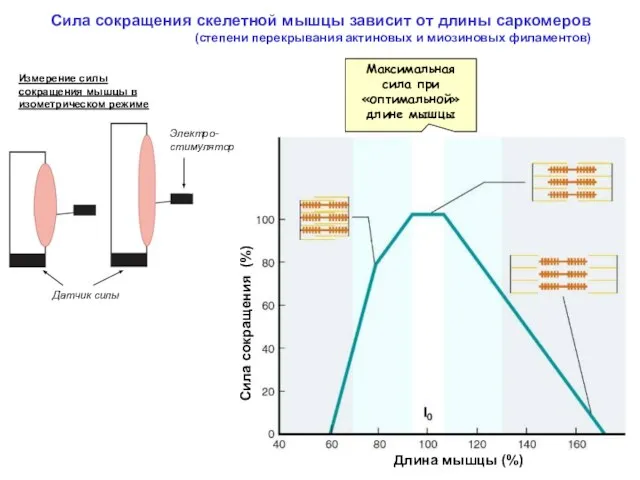
Сила сокращения скелетной мышцы зависит от длины саркомеров
(степени перекрывания актиновых
Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых

Основные пути Са2+-зависимой активации актомиозинового взаимодействия
Актин = «рельсы»
Миозиновый мотор
КПД 50-80%
(у
Основные пути Са2+-зависимой активации актомиозинового взаимодействия
Актин = «рельсы»
Миозиновый мотор
КПД 50-80%
(у
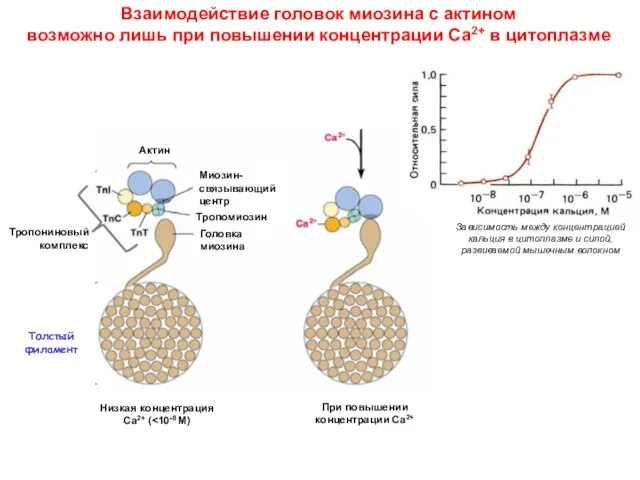
Взаимодействие головок миозина с актином
возможно лишь при повышении концентрации Са2+
Взаимодействие головок миозина с актином возможно лишь при повышении концентрации Са2+
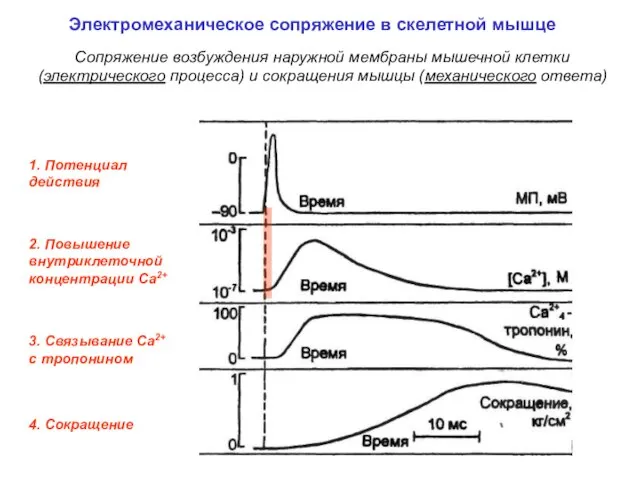
1. Потенциал действия
3. Связывание Са2+
c тропонином
4. Сокращение
2. Повышение внутриклеточной концентрации
1. Потенциал действия
3. Связывание Са2+
c тропонином
4. Сокращение
2. Повышение внутриклеточной концентрации
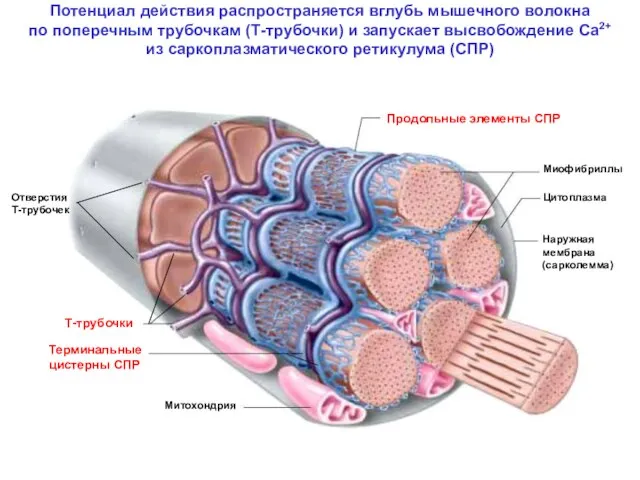
Потенциал действия распространяется вглубь мышечного волокна
по поперечным трубочкам (Т-трубочки) и
Потенциал действия распространяется вглубь мышечного волокна по поперечным трубочкам (Т-трубочки) и
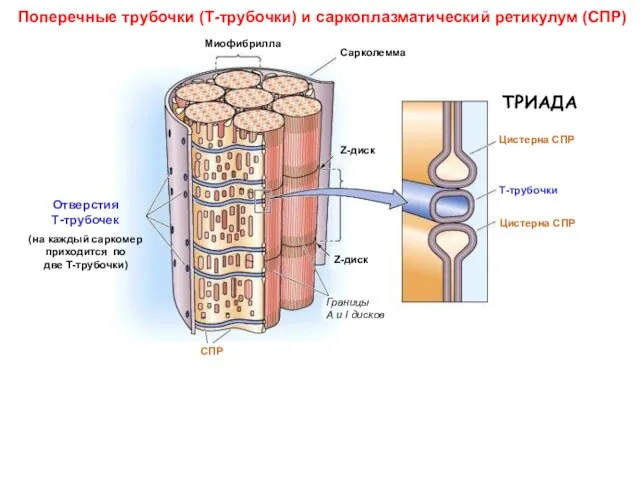
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
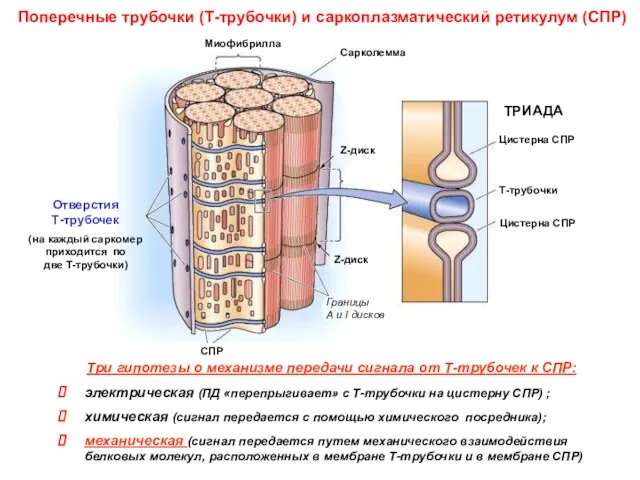
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Три гипотезы о механизме
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Три гипотезы о механизме
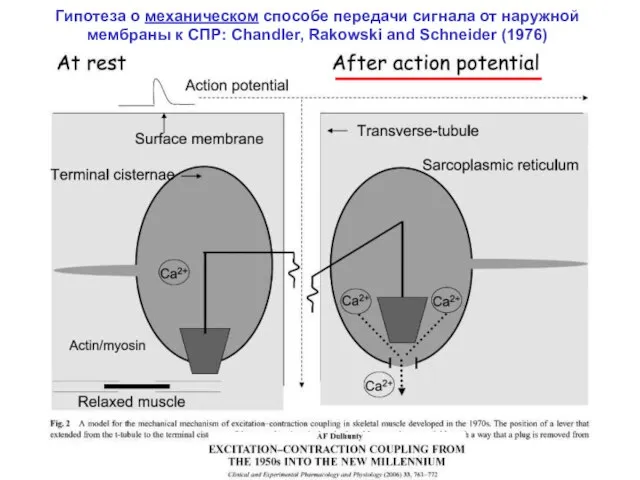
Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР:
Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР:
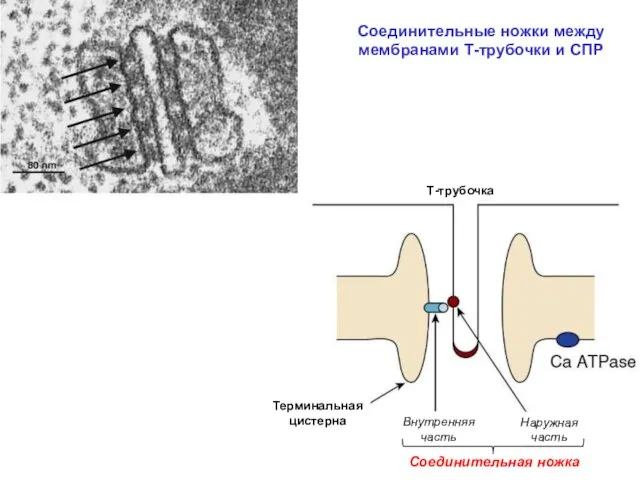
Соединительные ножки между мембранами Т-трубочки и СПР
Соединительные ножки между мембранами Т-трубочки и СПР
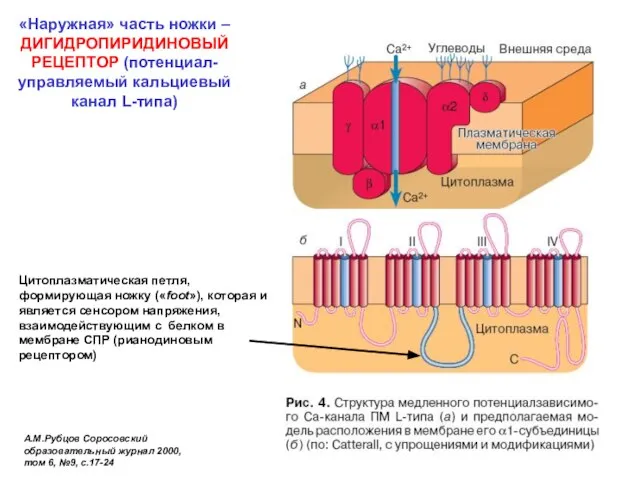
«Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-управляемый кальциевый канал L-типа)
А.М.Рубцов Соросовский
«Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-управляемый кальциевый канал L-типа)
А.М.Рубцов Соросовский
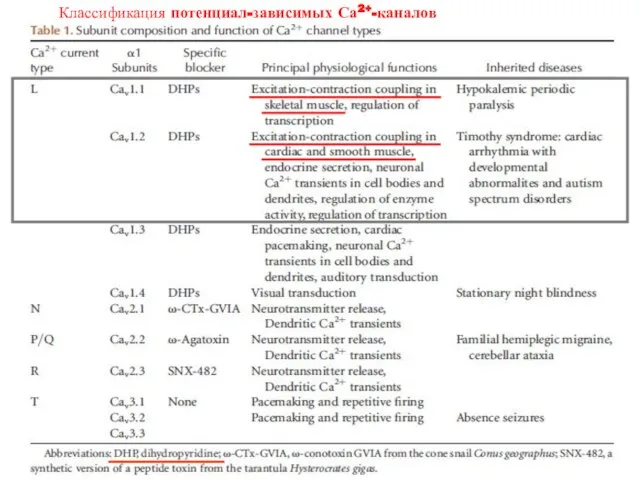
Классификация потенциал-зависимых Са2+-каналов
Классификация потенциал-зависимых Са2+-каналов
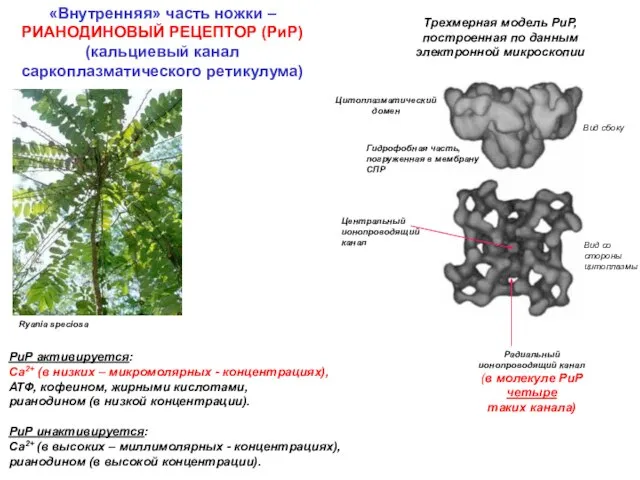
Ryania speciosa
Трехмерная модель РиР,
построенная по данным электронной микроскопии
Вид сбоку
Вид со
Ryania speciosa
Трехмерная модель РиР,
построенная по данным электронной микроскопии
Вид сбоку
Вид со
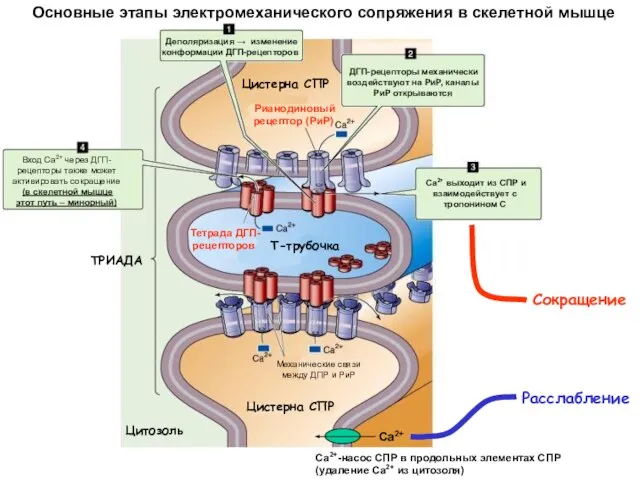
Основные этапы электромеханического сопряжения в скелетной мышце
Основные этапы электромеханического сопряжения в скелетной мышце
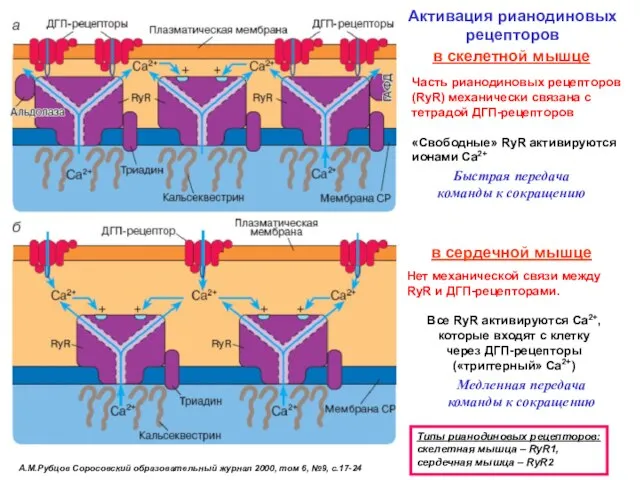
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
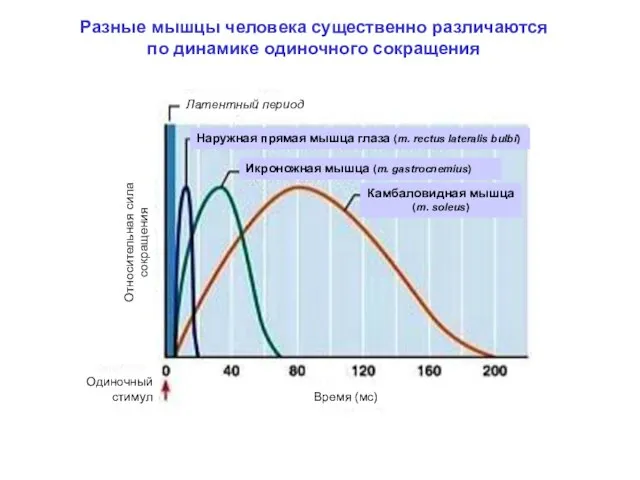
Разные мышцы человека существенно различаются
по динамике одиночного сокращения
Разные мышцы человека существенно различаются
по динамике одиночного сокращения
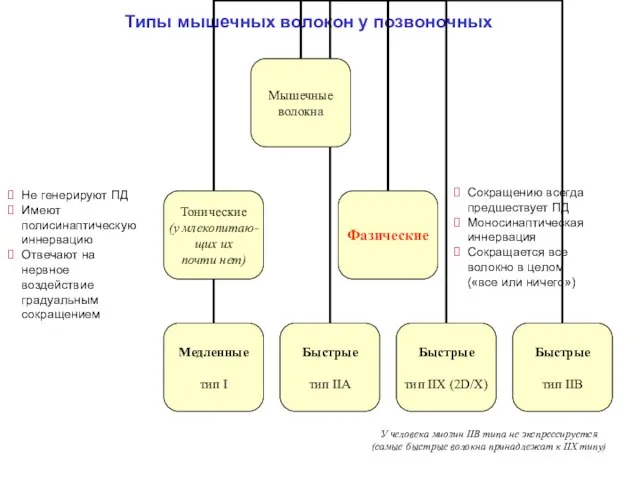
Типы мышечных волокон у позвоночных
Не генерируют ПД
Имеют полисинаптическую иннервацию
Отвечают на нервное
Типы мышечных волокон у позвоночных
Не генерируют ПД
Имеют полисинаптическую иннервацию
Отвечают на нервное

Поперечные серийные срезы четырехглавой мышцы бедра человека, окрашенные антителами против медленных
Поперечные серийные срезы четырехглавой мышцы бедра человека, окрашенные антителами против медленных
Toniolo et al. Am J Physiol. 2007 292(5):C1915-26.
In vitro motility
Toniolo et al. Am J Physiol. 2007 292(5):C1915-26.
In vitro motility
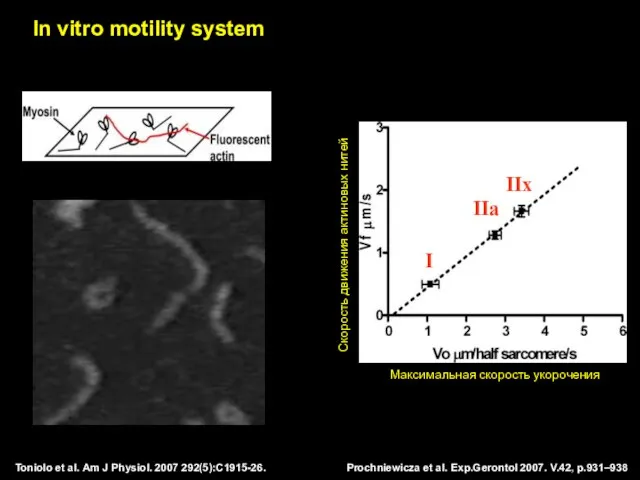
Prochniewicza et al. Exp.Gerontol 2007. V.42, p.931–938
Toniolo et al. Am J
Prochniewicza et al. Exp.Gerontol 2007. V.42, p.931–938
Toniolo et al. Am J

Источники АТФ в скелетной мышце
Источники АТФ в скелетной мышце
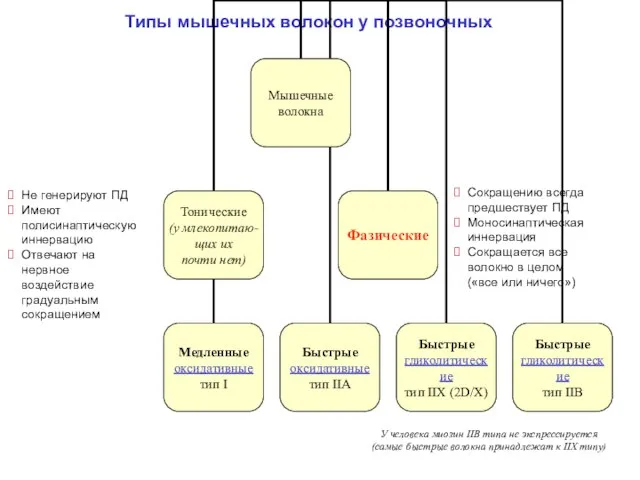
Типы мышечных волокон у позвоночных
Не генерируют ПД
Имеют полисинаптическую иннервацию
Отвечают на нервное
Типы мышечных волокон у позвоночных
Не генерируют ПД
Имеют полисинаптическую иннервацию
Отвечают на нервное
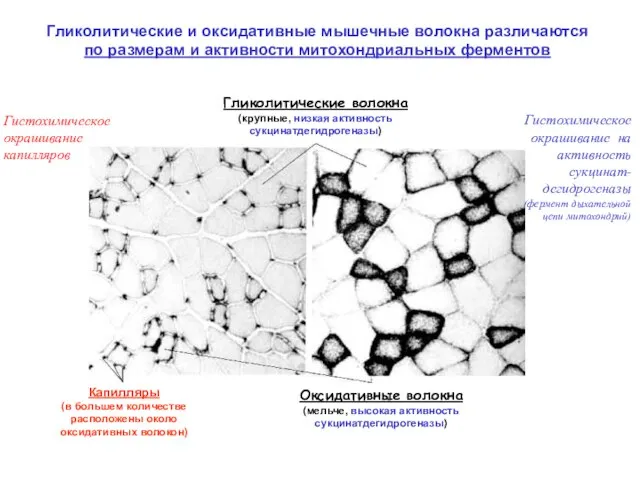
Гликолитические и оксидативные мышечные волокна различаются
по размерам и активности митохондриальных
Гликолитические и оксидативные мышечные волокна различаются по размерам и активности митохондриальных
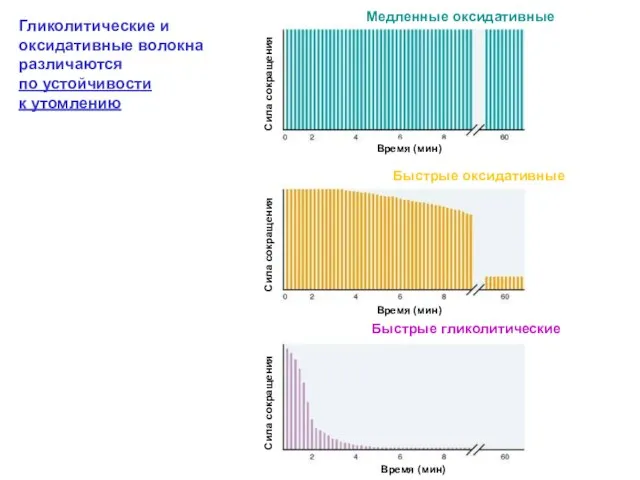
Гликолитические и оксидативные волокна различаются
по устойчивости
к утомлению
Гликолитические и оксидативные волокна различаются
по устойчивости
к утомлению
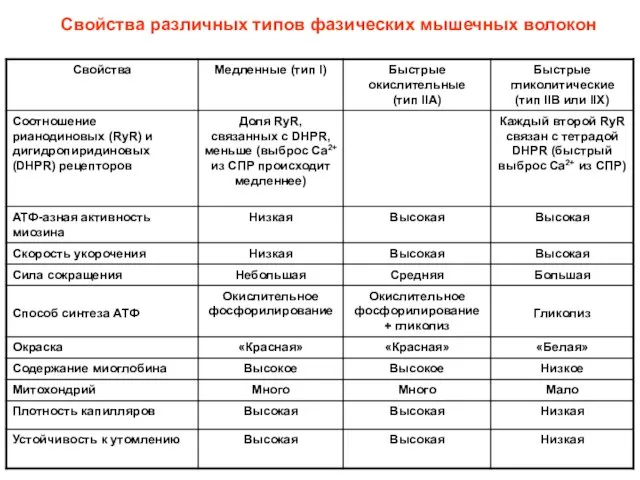
Свойства различных типов фазических мышечных волокон
Свойства различных типов фазических мышечных волокон
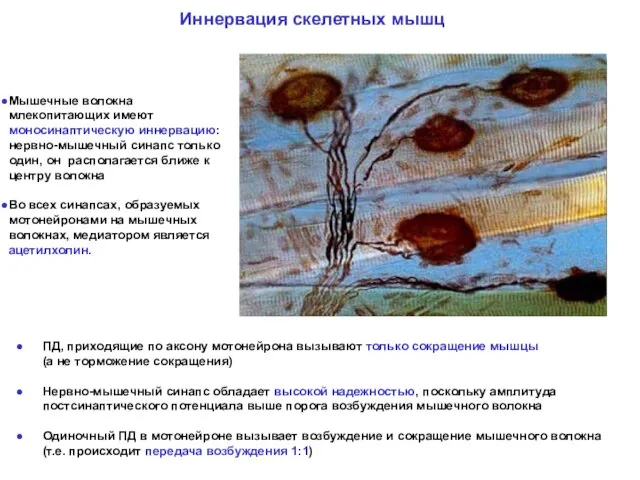
ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы
(а не
ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы (а не
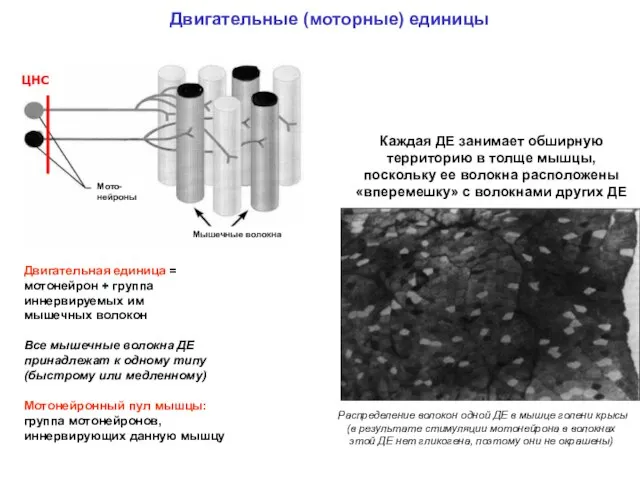
Двигательные (моторные) единицы
Двигательная единица = мотонейрон + группа иннервируемых им
мышечных
Двигательные (моторные) единицы
Двигательная единица = мотонейрон + группа иннервируемых им мышечных
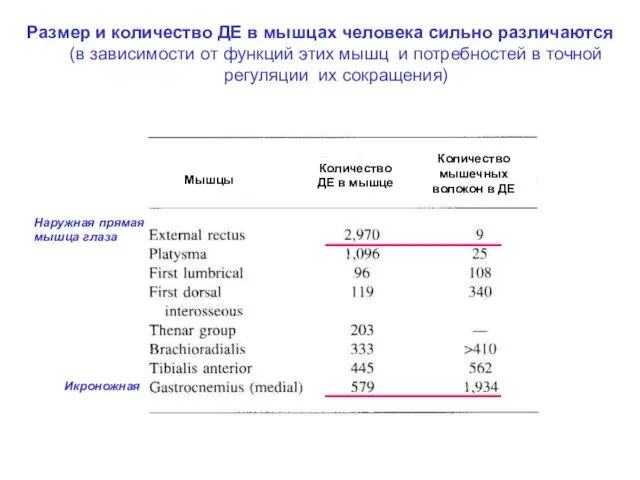
Размер и количество ДЕ в мышцах человека сильно различаются
(в зависимости
Размер и количество ДЕ в мышцах человека сильно различаются (в зависимости
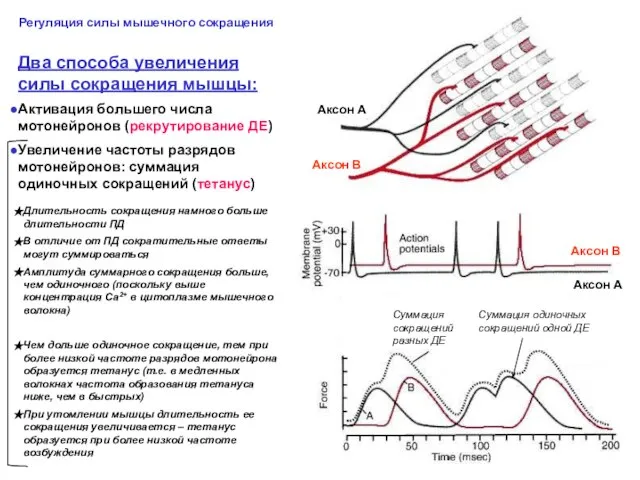
Два способа увеличения
силы сокращения мышцы:
Активация большего числа мотонейронов (рекрутирование ДЕ)
Увеличение
Два способа увеличения
силы сокращения мышцы:
Активация большего числа мотонейронов (рекрутирование ДЕ)
Увеличение
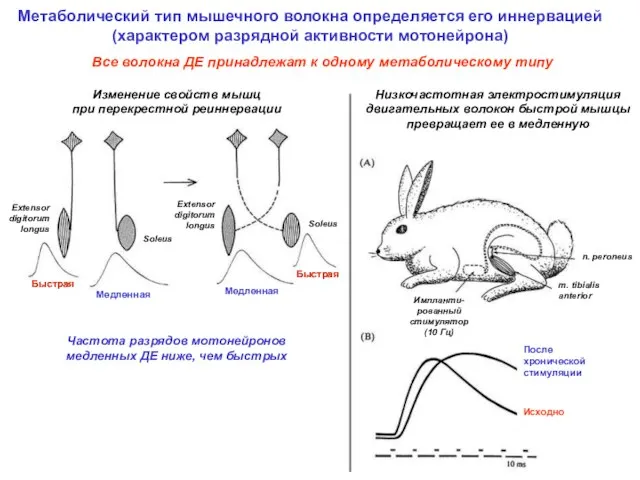
Метаболический тип мышечного волокна определяется его иннервацией
(характером разрядной активности мотонейрона)
Изменение
Метаболический тип мышечного волокна определяется его иннервацией
(характером разрядной активности мотонейрона)
Изменение

Изменения мышц при естественной активности мотонейронов
Тренировка силы
(тяжелая атлетика)
Тренировка выносливости
(марафон)
Как выглядят тела
Изменения мышц при естественной активности мотонейронов
Тренировка силы
(тяжелая атлетика)
Тренировка выносливости
(марафон)
Как выглядят тела
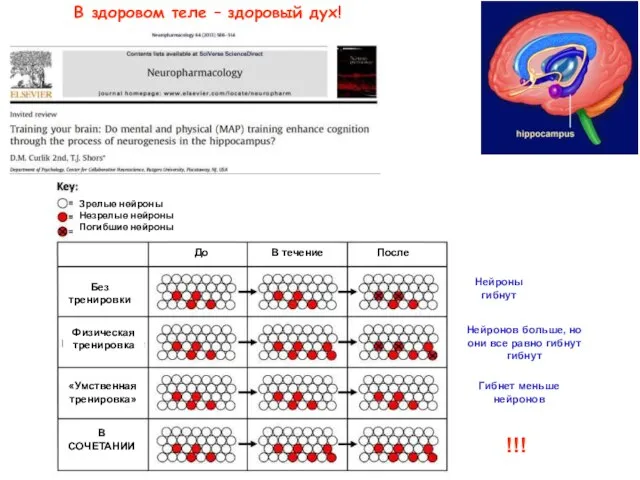
В здоровом теле – здоровый дух!
В здоровом теле – здоровый дух!

Спасибо за внимание!
Спасибо за внимание!
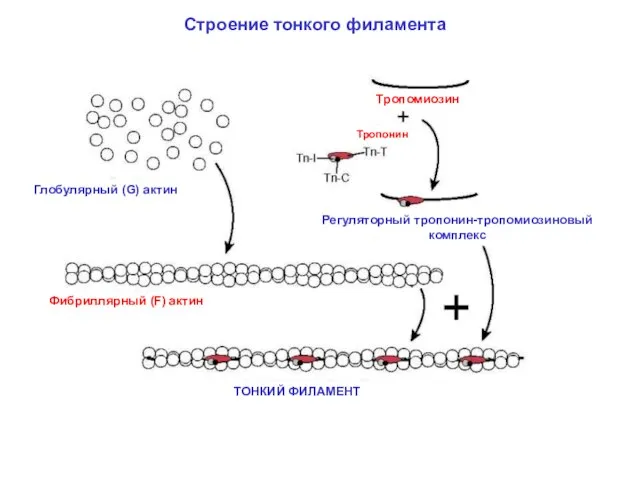
Строение тонкого филамента
Строение тонкого филамента
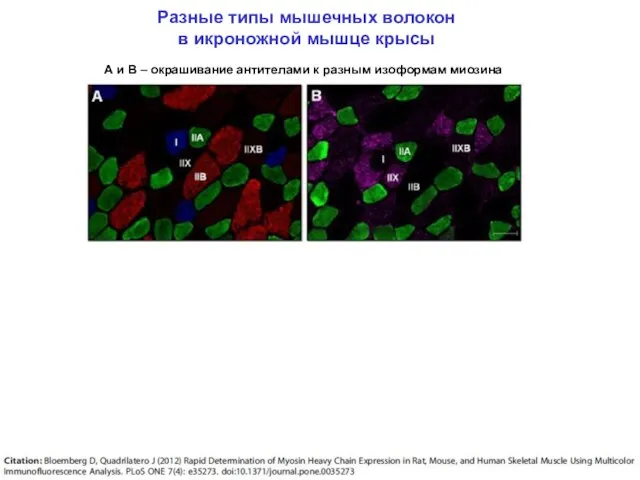
Разные типы мышечных волокон
в икроножной мышце крысы
А и В –
Разные типы мышечных волокон
в икроножной мышце крысы
А и В –
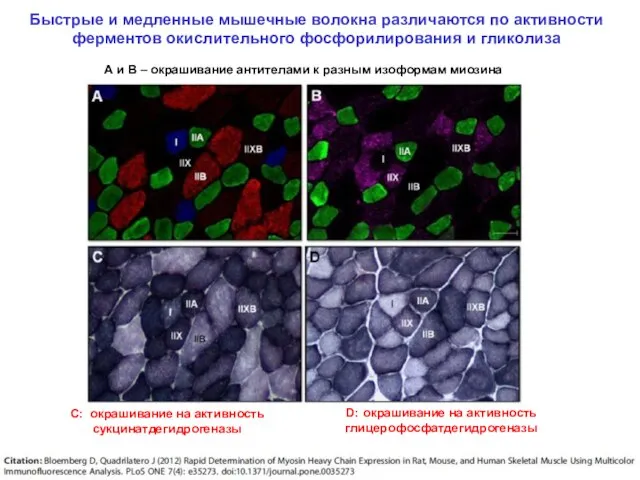
Быстрые и медленные мышечные волокна различаются по активности ферментов окислительного фосфорилирования
Быстрые и медленные мышечные волокна различаются по активности ферментов окислительного фосфорилирования
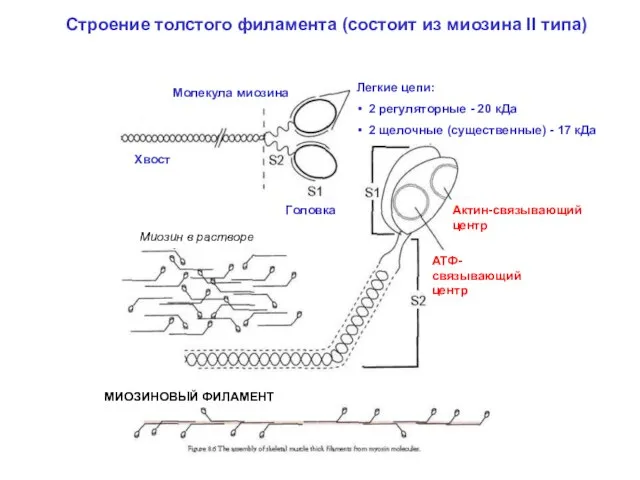
Строение толстого филамента (состоит из миозина II типа)
Строение толстого филамента (состоит из миозина II типа)
 Удивительный мир динозавров
Удивительный мир динозавров Презентация на тему "Гипотеза биохимической эволюции" - скачать презентации по Биологии
Презентация на тему "Гипотеза биохимической эволюции" - скачать презентации по Биологии Морфология органов женской половой системы
Морфология органов женской половой системы Функции живого вещества
Функции живого вещества ТЕСТ "Жизнедеятельность клетки"
ТЕСТ "Жизнедеятельность клетки" Презентация на тему Клещевой энцефалит ( таёжный энцефалит)
Презентация на тему Клещевой энцефалит ( таёжный энцефалит) Насекомые
Насекомые Эволюция иммуннитета
Эволюция иммуннитета Выращивание улиток архахатин в домашних условиях
Выращивание улиток архахатин в домашних условиях Презентация по биологии Грибы
Презентация по биологии Грибы  цитологические основы насл
цитологические основы насл Строение органа слуха. Гигиена органа слуха
Строение органа слуха. Гигиена органа слуха Моллюски www.skachat-prezentaciju-besplatno.ru
Моллюски www.skachat-prezentaciju-besplatno.ru  Пресмыкающиеся (рептилии.)
Пресмыкающиеся (рептилии.) Внутреннее строение лягушки
Внутреннее строение лягушки Институт генетики и цитологии Национальной академии наук Беларуси
Институт генетики и цитологии Национальной академии наук Беларуси Наши органы чувств.
Наши органы чувств. Выполнил: Мустафин Тимур МОУ СОШ №156 2а класс Екатеринбург, 2011 г
Выполнил: Мустафин Тимур МОУ СОШ №156 2а класс Екатеринбург, 2011 г Введение в биотехнологию
Введение в биотехнологию Растительные сообщества и взаимосвязи в них.
Растительные сообщества и взаимосвязи в них. Размножение животных и воспроизводство населения. Исторические типы воспроизводства населения
Размножение животных и воспроизводство населения. Исторические типы воспроизводства населения Типы клеток
Типы клеток Тіршіліктің пайда болуы
Тіршіліктің пайда болуы Высшая нервная деятельность Поведение
Высшая нервная деятельность Поведение Биология клетки №1
Биология клетки №1 Отряд Грызуны Суслики
Отряд Грызуны Суслики Мутационная изменчивость. Виды мутаций. Мутагены
Мутационная изменчивость. Виды мутаций. Мутагены  Презентация по биологии Органы пищеварения
Презентация по биологии Органы пищеварения