- Метаболизм углеводов (дополнение)

Содержание
- 2. Вопросы из билетов по теме Моносахариды, олигосахариды. Полисахариды. Важнейшие представители. Химическое строение. Биологическая роль. Моносахариды, олигосахариды,
- 3. Структура темы Углеводы с точки зрения химии: структура, классификация. Функции углеводов Моносахариды Дисахариды Полисахариды АТФ и
- 4. УГЛЕВОДЫ: ХИМИЯ И ФУНКЦИИ
- 5. По химической структуре: Многоатомные спирты, содержащие альдегидную (альдозы, альдосахара) или кетогруппу (кетозы, кетосахара) Углеводы также делятся
- 6. Пример: разные способы охарактеризовать глюкозу (с точки зрения ее химической структуры) Альдоза Гексоза Если объединить 1-й
- 7. НАИБОЛЕЕ ВАЖНЫЕ УГЛЕВОДЫ
- 8. Важные моносахариды: рибоза, дезоксирибоза Рибоза, дезоксирибоза: входит в состав нуклеозидов, нуклеотидов, нуклеиновых кислот (ДНК, РНК) и
- 9. Важные моносахариды: глюкоза Глюкоза: основной источник энергии предшественник всех других углеводов входит в состав гликопротеинов
- 10. Важные моносахариды: фруктоза Фруктоза источник энергии может превращаться в глюкозу
- 11. Важные моносахариды: галактоза Галактоза источник энергии входит в состав лактозы – молочного сахара входит в состав
- 12. Важные моносахариды: манноза Манноза входит в состав гликопротеинов
- 13. Важные моносахариды: ксилоза Ксилоза входит в состав гликопротеинов (гепарансульфат, хондроитинсульфат)
- 14. Важные дисахариды: мальтоза Мальтоза глюкоза + глюкоза образуется в результате расщепления крахмала в кишечнике гидролизуется ферментом
- 15. Важные дисахариды: лактоза Лактоза (молочный сахар) галактоза + глюкоза содержится в молоке гидролизуется ферментом лактазой только
- 16. Важные дисахариды: сахароза сахароза глюкоза + фруктоза содержится в растениях гидролизуется ферментом сахаразой только в кишечнике
- 17. Важные полисахариды: гликоген и крахмал Гликоген (крахмал у растений) состоит из глюкозы основная запасная форма углеводов
- 18. Переваривание углеводов Сахараза, мальтаза и лактаза – ферменты класса гидролаз, которые расщепляют соответствующие дисахариды до моносахаридов.
- 19. АТФ И ЭНЕРГИЯ
- 20. АТФ: источник энергии для биохимических процессов По химической структуре АТФ является нуклеотидом: аденин (азотистое основание) +
- 21. Распад и синтез АТФ Распад АТФ в процессах, требующих затраты энергии представляет собой отщепление фосфата от
- 22. Существует всего 2 способа синтеза АТФ (т.е. фосфорилирования АДФ) 1. Субстратное фосфорилирование* Сначала образуется органическая молекула,
- 23. ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ РЕАКЦИИ (ОВР)
- 24. Общая схема ОВР: восстановленный субстрат отдает электроны окисленному окислителю Восстановленный субстрат Окисленный продукт Окисленный окислитель (кофермент)
- 25. ОВР (окислительно-восстановительные реакции в биохимии) В качестве одного из участников реакции часто выступает кофермент, принимающий или
- 26. Примеры ОВР окисление восстановление Самый восстановленный Самый окисленный
- 27. Конкретный пример (из ЦТК) Сукцинат окисляется до фумарата ФАД восстанавливается до ФАДН2 В этом примере сукцинат
- 28. ГЛИКОЛИЗ
- 29. Схема: Функции энергетическая промежуточные метаболиты гликолиза используются в синтезе аминокислот и липидов Внутриклеточная локализация: цитозоль Тканевая
- 30. 1-ю реакцию могут катализировать 2 фермента, в зависимости от ткани: гексокиназа (все ткани) или глюкокиназа (только
- 31. Гликолитическая оксидоредукция, или почему есть аэробный и анаэробный гликолиз 6-я реакция окислительно-восстановительная Фермент: глицеральдегидфосфатдегидрогеназа Кофермент: НАД
- 32. ГЛЮКОНЕОГЕНЕЗ
- 33. Глюконеогенез - синтез глюкозы из неуглеводных предшественников («субстратов глюконеогенеза») Схема: Функции: поддержание концентрации глюкозы в крови
- 34. На схеме показана последовательность гликолиза (сверху вниз) и глюконеогенеза (снизу вверх) На схеме: Реакции 1 –
- 35. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
- 36. Схема Функции: энергетическая связь между гликолизом и ЦТК Внутриклеточная локализация: митохондрии Тканевая локализация: все ткани, в
- 37. В результате реакции от пирувата отщепляется карбоксильная группа, которая уходит в форме СО2. Оставшиеся два атома
- 38. ЦИКЛ КРЕБСА (ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ)
- 39. Схема: Функции: Энергетическая Связь метаболизма аминокислот, углеводов и липидов Внутриклеточная локализация: матрикс митохондрий Тканевая локализация: все
- 40. Метаболиты и ферменты ЦТК 2 атома С входят в цикл в форме ацетил-КоА (реакция 1) 2
- 41. ДЫХАТЕЛЬНАЯ ЦЕПЬ И ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
- 42. Схема: Функция: энергетическая (большая часть АТФ синтезируется этим путем) Внутриклеточная локализация: митохондрии Тканевая локализация: все ткани,
- 43. Общая схема синтеза АТФ (хемиосмотическая гипотеза Митчелла) НАДН ФАДН2 Дыхательная цепь Градиент протонов АТФсинтаза Внешняя мембрана
- 44. Цепь переноса электронов и АТФсинтаза В реальности дыхательная цепь состоит из нескольких белков, которые работают друг
- 45. Подробное описание работы дыхательной цепи Комплекс I окисляет НАДН и восстанавливается (и выкачивает протоны из матрикса)
- 46. ГЛИКОГЕН: СТРУКТУРА, СИНТЕЗ, РАСПАД
- 47. Схема: Функция: Запасание энергии (синтез гликогена) Получение энергии и поддержание постоянной концентрации глюкозы в крови (распад
- 48. Гликоген: структура и особенности синтеза и распада Молекула гликогена разветвленная, поэтому в его структуре есть два
- 49. В пространстве молекула гликогена похожа на сферу (гранулы гликогена) Ветви гликогена в гранулах уложены в 12
- 50. Этапы синтеза гликогена В клетке всегда существуют небольшие молекулы гликогена, имеющие несколько веточек, которые можно удлинять
- 51. Синтез гликогена 1. Синтез глюкозо-1-фосфата Внутри клетки вся глюкоза находится в фосфорилированной форме (глюкозо-6-фосфат = г-6-Ф).
- 52. Синтез гликогена 2. Активация глюкозы (синтез УДФ-глюкозы) Теперь нужно активировать молекулу глюкозы, чтобы она могла присоединиться
- 53. Синтез гликогена 3. Удлинение молекулы гликогена На рисунке урацил в составе УДФ-глюкозы не нарисован, а просто
- 54. Синтез гликогена 4.Ветвление молекулы гликогена Когда ветка гликогена достигает большой длины (примерно 11 глюкозных звеньев), происходит
- 55. Этапы распада гликогена Ферменты укорачивают отдельные ветки молекул гликогена или удаляют некоторые из этих веток полностью.
- 56. Распад гликогена 1. Укорачивание линейных участков (фосфоролиз = фосфоролитическое расщепление) с образованием глюкозо-1-фосфата Разрыв связи между
- 57. Распад гликогена 2. Превращение глюкозо-1-фосфата в глюкозо-6-фосфат и его дальнейшая судьба Образовавшийся глюкозо-1-фосфат может использоваться в
- 58. Распад гликогена 3. Удаление точек ветвления Красным показаны звенья (глюкозы), которые удаляются фосфорилазой (см. этап 1)
- 59. Нарушения метаболизма гликогена - гликогенозы Ферменты, участвующие в синтезе и распаде гликогена, разные в печени и
- 60. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
- 61. Схема: Функции Образование рибозы для синтеза нуклеотидов Образование НАДФН для синтеза липидов и защиты от свободных
- 62. НАДН vs. НАДФН Оба являются коферментами ОВР Оба содержат в составе витамин РР (никотинамид) НАДФН: используется
- 63. Две стадии: Окислительная: глюкозо-6-фосфат (С6) превращается в рибулозо-5-фосфат (С5), образуются 2 НАДФН Неокислительная: рибулозо-5-фосфат (С5) превращается
- 64. I. Окислительная стадия Целями первой стадии являются: Превращение 6-углеродной глюкозы в 5-углеродную рибозу получение НАДФН в
- 65. Целью неокислительной стадии является превращение нескольких 5-углеродных молекул в 6-углеродные, чтобы они снова могли вступить в
- 66. Полная схема пентозофосфатного пути Обратите внимание на метаболиты неокислительной стадии: среди них встречаются вещества, принимающие участие
- 67. Нарушения пентозофосфатного пути Наследственное заболевание, связанное с нарушением работы пентозофосфатного пути: лекарственная гемолитическая анемия Фермент, снижение
- 68. Участие глутатиона и пентозофосфатного пути в защите от свободных радикалов Глутатионпероксидаза при участии глутатиона инактивирует перекись
- 69. РЕГУЛЯЦИЯ МЕТАБОЛИЗМА УГЛЕВОДОВ
- 70. Регуляторные ферменты В любом метаболическом пути есть два типа ферментов (с точки зрения возможности изменения скорости
- 71. 1 2 3 4 5 6 7 8 9 10 11 Регуляторные ферменты в гликолизе
- 72. Общие принципы регуляции работы ферментов Изменение скорости работы ферментов происходит путем их: фосфорилирования киназами и/или присоединения/удаления
- 73. Регуляторные ферменты гликолиза Необратимые реакции (они же регуляторные = ключевые = скорость-лимитирующие = аллостерические): 1 –
- 74. Аллостерические регуляторы гликолиза Тормозят гликолиз: АТФ, цитрат, глюкозо-6-фосфат (мышцы), фруктозо-6-фосфат (печень), ацетил-КоА, ВЖК Ускоряют гликолиз: АДФ,
- 75. Гормональная регуляция гликолиза Тормозит: глюкагон Активируют: инсулин, адреналин (в мышцах)
- 76. Регуляция гликолиза и глюконеогенеза
- 77. РЕГУЛЯЦИЯ СИНТЕЗА И РАСПАДА ГЛИКОГЕНА
- 78. Регуляторные ферменты Гликогенсинтаза – регуляторный фермент синтеза гликогена. Неактивна в фосфорилированной форме, активна – в нефосфорилированной
- 79. Фосфорилированные и нефосфорилированные формы гликогенфосфорилазы (вверху) и гликогенсинтазы (внизу) Традиционно активные формы этих ферментов обозначаются буквой
- 80. Описание сигнального каскада адреналина и глюкагона – см. ниже ПКА = протеинкиназа А На схеме показана
- 81. РЕГУЛЯЦИЯ ЦТК
- 82. Регуляторные ферменты Цитратсинтаза: Ингибируют: НАДН, сукцинил-КоА, цитрат, АТФ Активирует: АДФ Изоцитратдегидрогеназа Ингибирует: АТФ Активируют: Ca2+, АДФ
- 84. ФИЗИОЛОГИЧЕСКИЕ ЭФФЕКТЫ ИНСУЛИНА: МЕТАБОЛИЗМ ГЛЮКОЗЫ, АМИНОКИСЛОТ, ЛИПИДОВ
- 85. Влияние на метаболизм углеводов Активация поступления глюкозы в клетки, снижение глюкозы в крови Стимуляция гликолиза Стимуляция
- 86. Влияние инсулина на концентрацию глюкозы в крови После еду глюкоза поступает в кровь Это вызывает высвобождение
- 87. Механизм действия инсулина на клетки На схеме показана мембрана клетки Глюкоза поступает снаружи клетк После связывания
- 88. Влияние на метаболизм липидов Ингибирует гормон-чувствительную липазу адипоцитов Активирует липогенез (синтез липидов) в печени, в том
- 89. Влияние на метаболизм белков и аминокислот Активация синтеза белков Активация захвата аминокислот тканями
- 90. САХАРНЫЙ ДИАБЕТ: ПРИЧИНЫ И ТИПЫ
- 91. СД 1 типа Иммунные: аутоантитела к инсулину и специфическим белкам бета-клеток. Разрушение поджелудочной железы разной степени
- 92. СД 2 типа Инсулинорезистентность – невосприимчивость (устойчивость) клеток к сигналу, который им отдает инсулин Проявляется в
- 93. ГЛЮКАГОН И АДРЕНАЛИН
- 94. Глюкагон и адреналин Глюкагон – пептидный гормон Адреналин – производное аминокислоты тирозина Оба гормона обладают сходными
- 95. ИНТЕГРАЦИЯ МЕТАБОЛИЗМА
- 96. «Метаболическая воронка» Это понятие, которое описывает унификацию получения энергии из разных источников Энергию мы можем получать
- 97. Окисление всех питательных веществ с образованием ацетил-КоА (или метаболитов цикла Кребса). Также образуются НАДН и ФАДН2.
- 98. Ацетил-КоА пируват глюкоза Серин, глицин Глюкозо-6-Ф Кетоновые тела ВЖК холестерин α-кетоглутарат Глутамат и другие а.к. оксалоацетат
- 100. Скачать презентацию

Вопросы из билетов по теме
Моносахариды, олигосахариды. Полисахариды. Важнейшие представители. Химическое строение.
Вопросы из билетов по теме
Моносахариды, олигосахариды. Полисахариды. Важнейшие представители. Химическое строение.

Структура темы
Углеводы с точки зрения химии: структура, классификация. Функции углеводов
Моносахариды
Дисахариды
Полисахариды
АТФ и
Структура темы
Углеводы с точки зрения химии: структура, классификация. Функции углеводов
Моносахариды
Дисахариды
Полисахариды
АТФ и

УГЛЕВОДЫ: ХИМИЯ И ФУНКЦИИ
УГЛЕВОДЫ: ХИМИЯ И ФУНКЦИИ

По химической структуре:
Многоатомные спирты, содержащие альдегидную (альдозы, альдосахара) или кетогруппу (кетозы,
По химической структуре:
Многоатомные спирты, содержащие альдегидную (альдозы, альдосахара) или кетогруппу (кетозы,
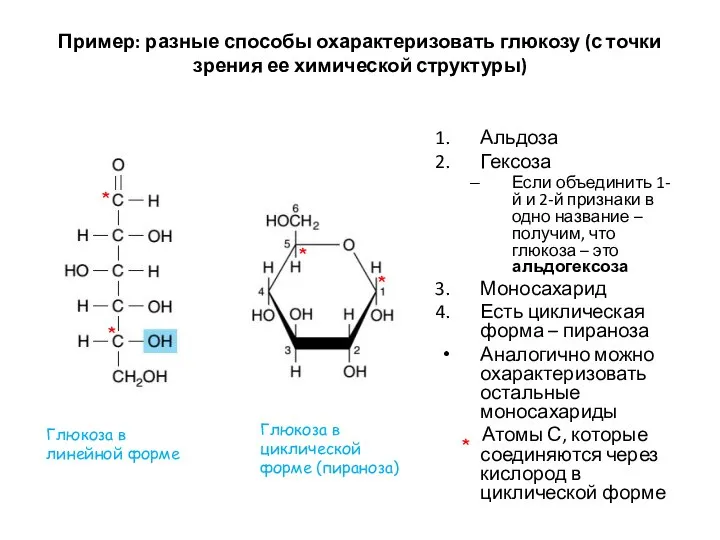
Пример: разные способы охарактеризовать глюкозу (с точки зрения ее химической структуры)
Альдоза
Гексоза
Если
Пример: разные способы охарактеризовать глюкозу (с точки зрения ее химической структуры)
Альдоза
Гексоза
Если

НАИБОЛЕЕ ВАЖНЫЕ УГЛЕВОДЫ
НАИБОЛЕЕ ВАЖНЫЕ УГЛЕВОДЫ
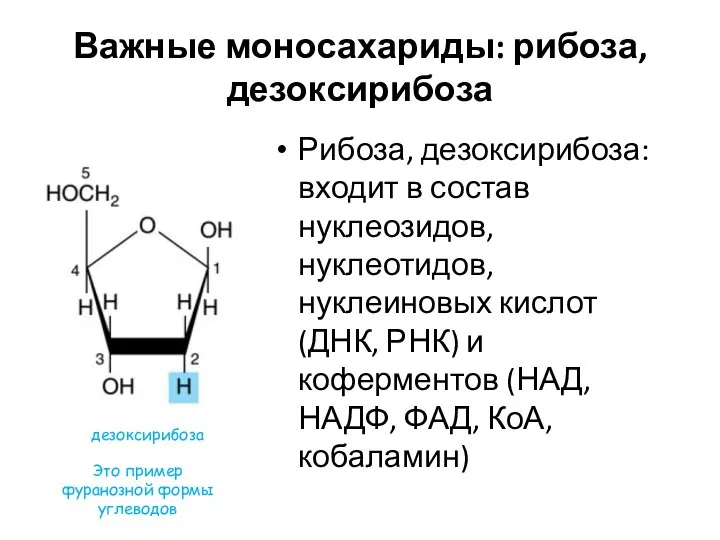
Важные моносахариды: рибоза, дезоксирибоза
Рибоза, дезоксирибоза: входит в состав нуклеозидов, нуклеотидов, нуклеиновых
Важные моносахариды: рибоза, дезоксирибоза
Рибоза, дезоксирибоза: входит в состав нуклеозидов, нуклеотидов, нуклеиновых
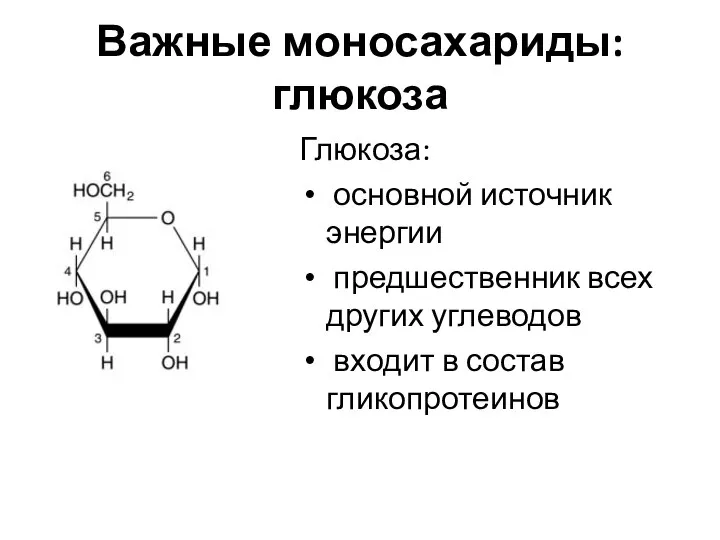
Важные моносахариды: глюкоза
Глюкоза:
основной источник энергии
предшественник всех других углеводов
входит
Важные моносахариды: глюкоза
Глюкоза:
основной источник энергии
предшественник всех других углеводов
входит
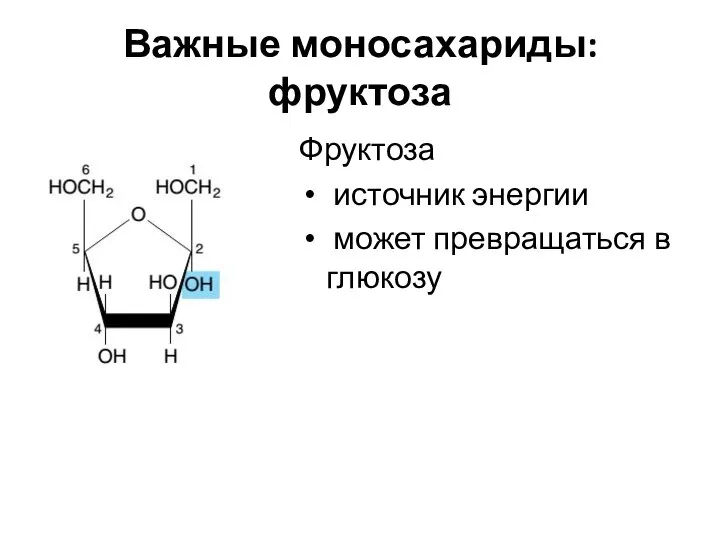
Важные моносахариды: фруктоза
Фруктоза
источник энергии
может превращаться в глюкозу
Важные моносахариды: фруктоза
Фруктоза
источник энергии
может превращаться в глюкозу
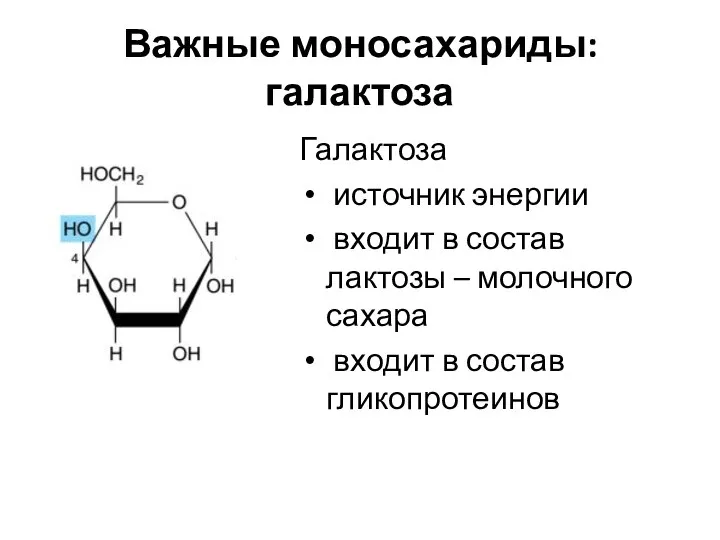
Важные моносахариды: галактоза
Галактоза
источник энергии
входит в состав лактозы – молочного
Важные моносахариды: галактоза
Галактоза
источник энергии
входит в состав лактозы – молочного
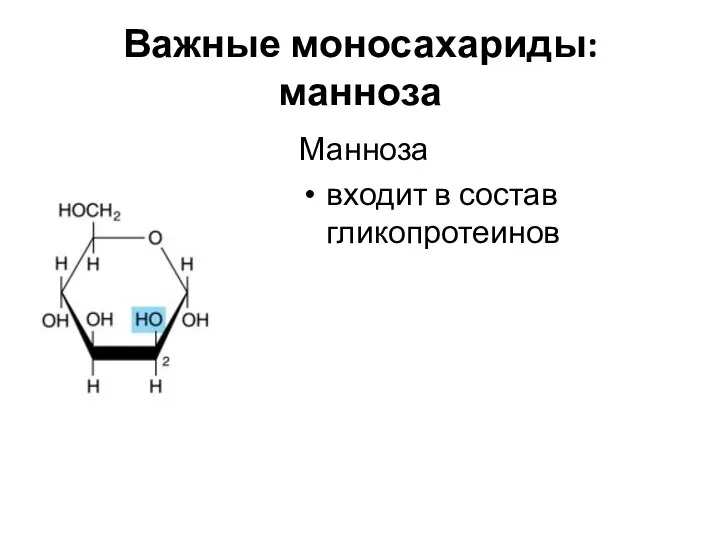
Важные моносахариды: манноза
Манноза
входит в состав гликопротеинов
Важные моносахариды: манноза
Манноза
входит в состав гликопротеинов
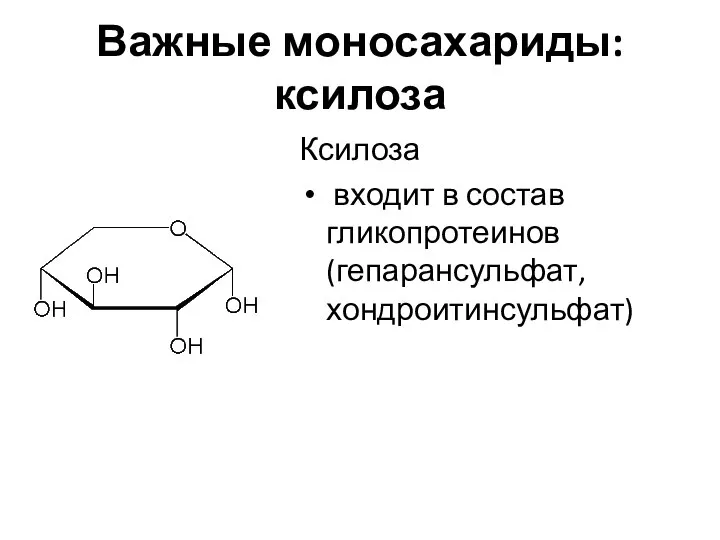
Важные моносахариды: ксилоза
Ксилоза
входит в состав гликопротеинов (гепарансульфат, хондроитинсульфат)
Важные моносахариды: ксилоза
Ксилоза
входит в состав гликопротеинов (гепарансульфат, хондроитинсульфат)
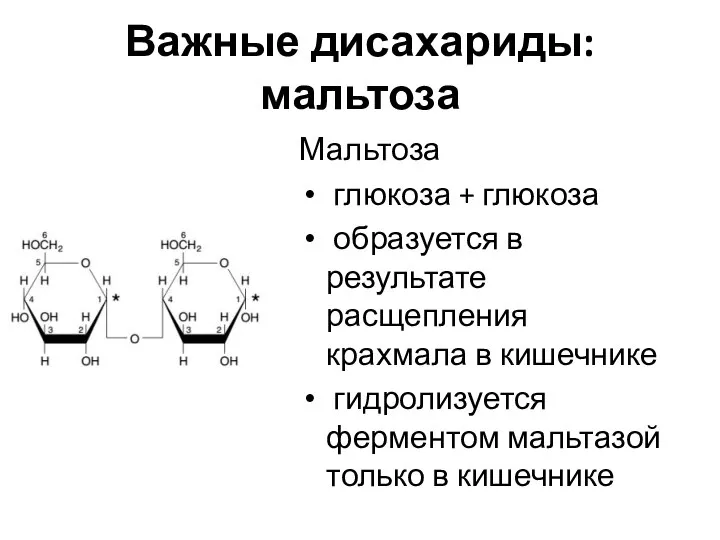
Важные дисахариды: мальтоза
Мальтоза
глюкоза + глюкоза
образуется в результате расщепления крахмала
Важные дисахариды: мальтоза
Мальтоза
глюкоза + глюкоза
образуется в результате расщепления крахмала
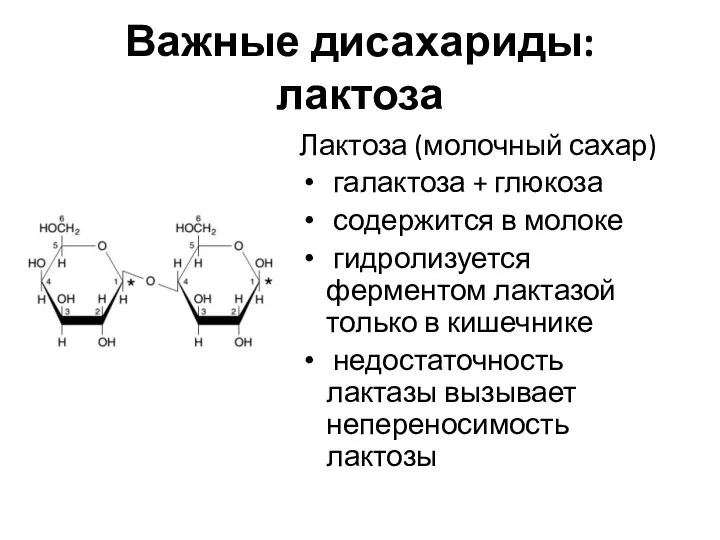
Важные дисахариды: лактоза
Лактоза (молочный сахар)
галактоза + глюкоза
содержится в молоке
Важные дисахариды: лактоза
Лактоза (молочный сахар)
галактоза + глюкоза
содержится в молоке
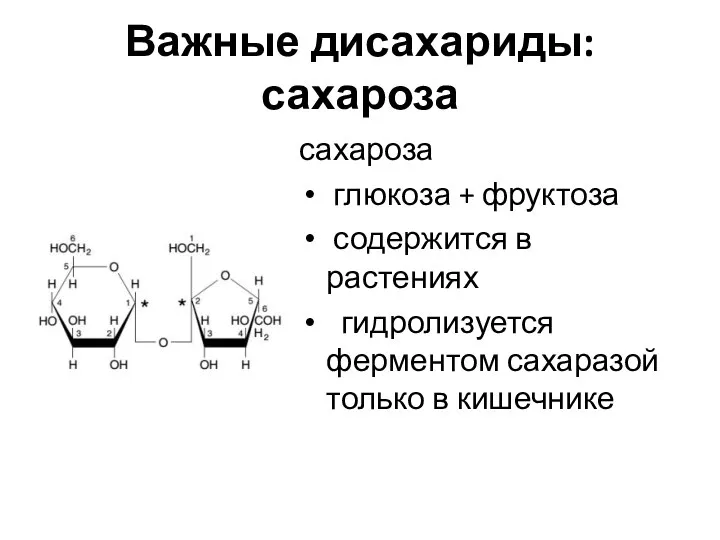
Важные дисахариды: сахароза
сахароза
глюкоза + фруктоза
содержится в растениях
гидролизуется ферментом
Важные дисахариды: сахароза
сахароза
глюкоза + фруктоза
содержится в растениях
гидролизуется ферментом

Важные полисахариды: гликоген и крахмал
Гликоген (крахмал у растений)
состоит из глюкозы
Важные полисахариды: гликоген и крахмал
Гликоген (крахмал у растений)
состоит из глюкозы

Переваривание углеводов
Сахараза, мальтаза и лактаза – ферменты класса гидролаз, которые расщепляют
Переваривание углеводов
Сахараза, мальтаза и лактаза – ферменты класса гидролаз, которые расщепляют

АТФ И ЭНЕРГИЯ
АТФ И ЭНЕРГИЯ
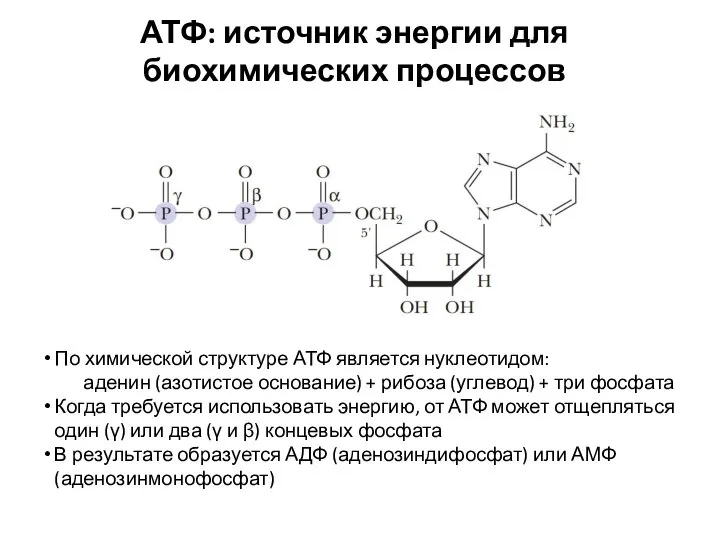
АТФ: источник энергии для биохимических процессов
По химической структуре АТФ является нуклеотидом:
аденин
АТФ: источник энергии для биохимических процессов
По химической структуре АТФ является нуклеотидом:
аденин
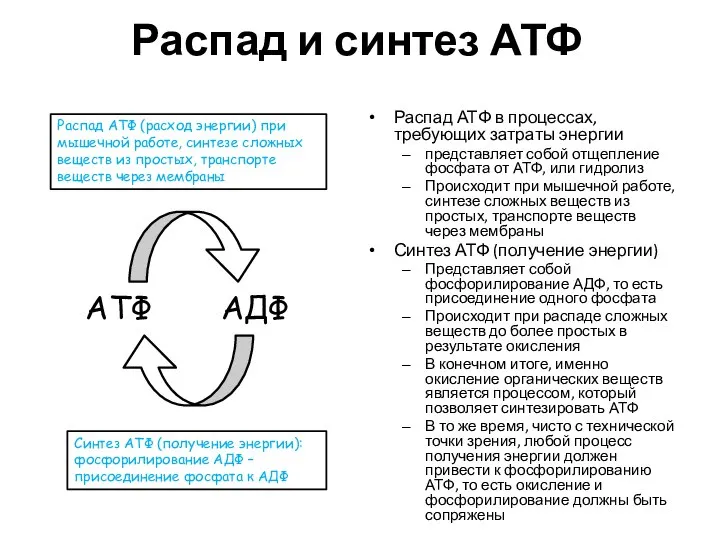
Распад и синтез АТФ
Распад АТФ в процессах, требующих затраты энергии
представляет собой
Распад и синтез АТФ
Распад АТФ в процессах, требующих затраты энергии
представляет собой

Существует всего 2 способа синтеза АТФ (т.е. фосфорилирования АДФ)
1. Субстратное фосфорилирование*
Сначала
Существует всего 2 способа синтеза АТФ (т.е. фосфорилирования АДФ)
1. Субстратное фосфорилирование*
Сначала

ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ РЕАКЦИИ (ОВР)
ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ РЕАКЦИИ (ОВР)
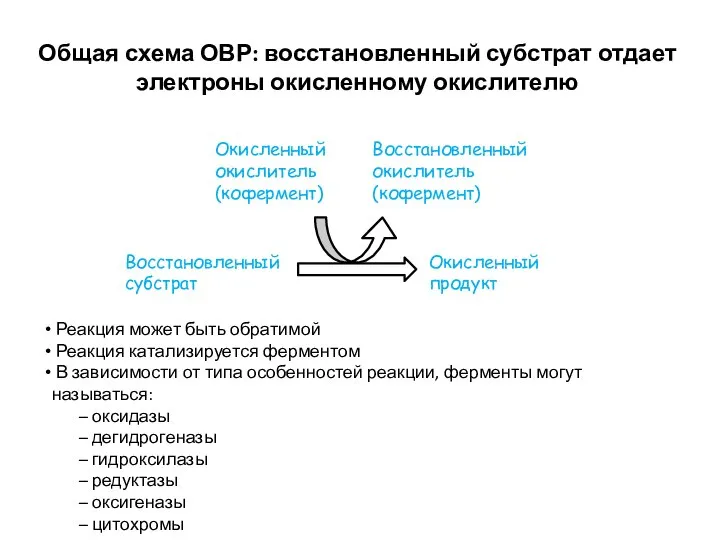
Общая схема ОВР: восстановленный субстрат отдает электроны окисленному окислителю
Восстановленный субстрат
Окисленный продукт
Окисленный
Общая схема ОВР: восстановленный субстрат отдает электроны окисленному окислителю
Восстановленный субстрат
Окисленный продукт
Окисленный

ОВР (окислительно-восстановительные реакции в биохимии)
В качестве одного из участников реакции часто
ОВР (окислительно-восстановительные реакции в биохимии)
В качестве одного из участников реакции часто
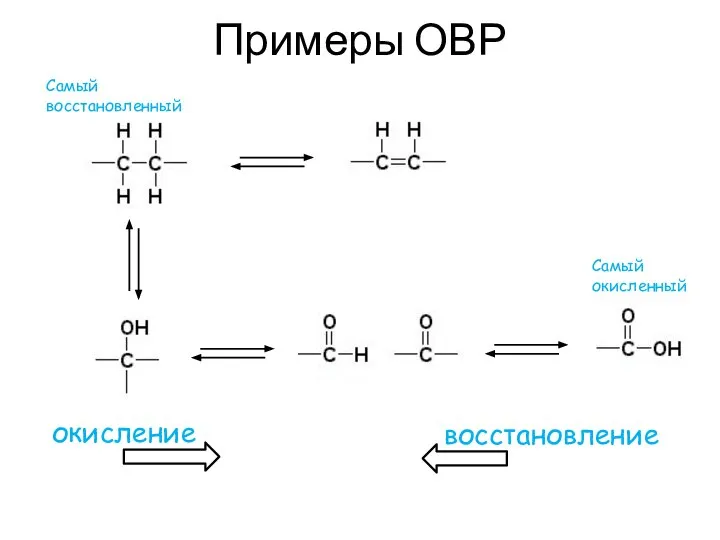
Примеры ОВР
окисление
восстановление
Самый восстановленный
Самый окисленный
Примеры ОВР
окисление
восстановление
Самый восстановленный
Самый окисленный
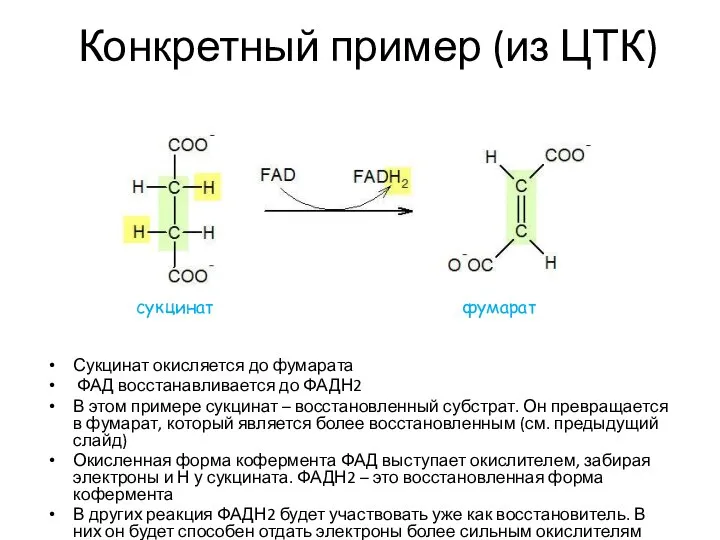
Конкретный пример (из ЦТК)
Сукцинат окисляется до фумарата
ФАД восстанавливается до ФАДН2
В
Конкретный пример (из ЦТК)
Сукцинат окисляется до фумарата
ФАД восстанавливается до ФАДН2
В

ГЛИКОЛИЗ
ГЛИКОЛИЗ
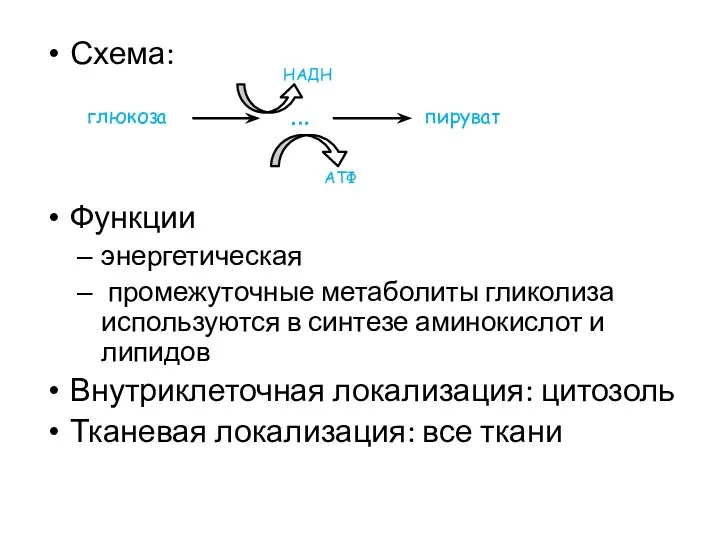
Схема:
Функции
энергетическая
промежуточные метаболиты гликолиза используются в синтезе аминокислот и липидов
Внутриклеточная локализация:
Схема:
Функции
энергетическая
промежуточные метаболиты гликолиза используются в синтезе аминокислот и липидов
Внутриклеточная локализация:

1-ю реакцию могут катализировать 2 фермента, в зависимости от ткани: гексокиназа
1-ю реакцию могут катализировать 2 фермента, в зависимости от ткани: гексокиназа
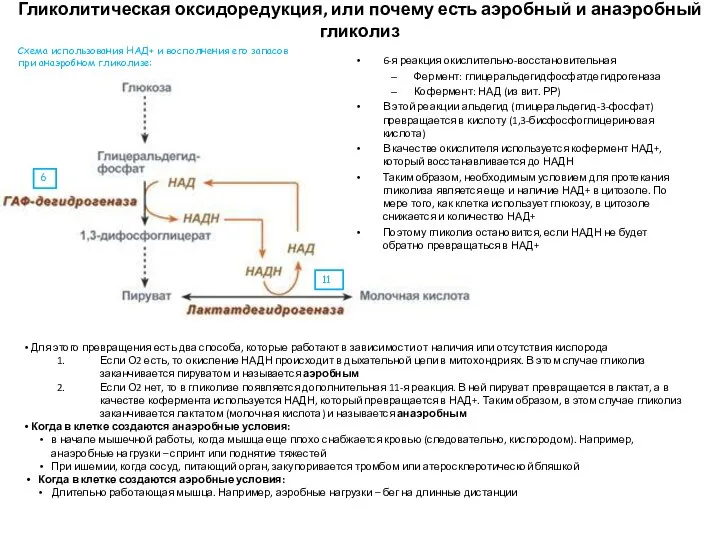
Гликолитическая оксидоредукция, или почему есть аэробный и анаэробный гликолиз
6-я реакция окислительно-восстановительная
Фермент:
Гликолитическая оксидоредукция, или почему есть аэробный и анаэробный гликолиз
6-я реакция окислительно-восстановительная
Фермент:

ГЛЮКОНЕОГЕНЕЗ
ГЛЮКОНЕОГЕНЕЗ
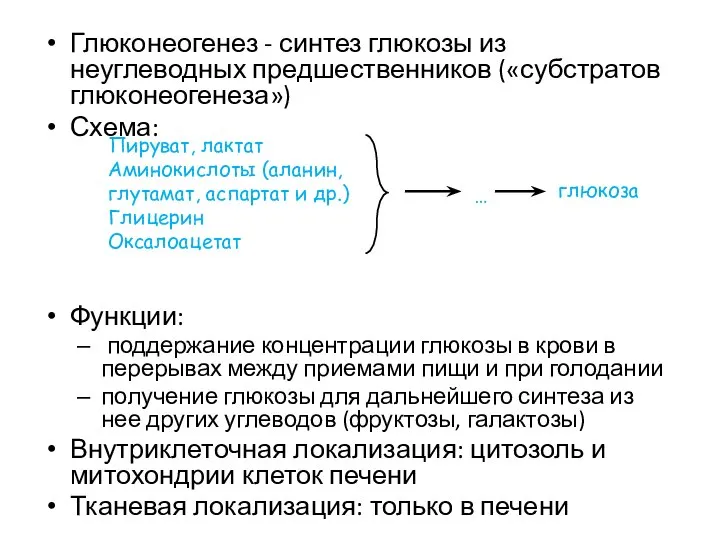
Глюконеогенез - синтез глюкозы из неуглеводных предшественников («субстратов глюконеогенеза»)
Схема:
Функции:
поддержание концентрации
Глюконеогенез - синтез глюкозы из неуглеводных предшественников («субстратов глюконеогенеза»)
Схема:
Функции:
поддержание концентрации
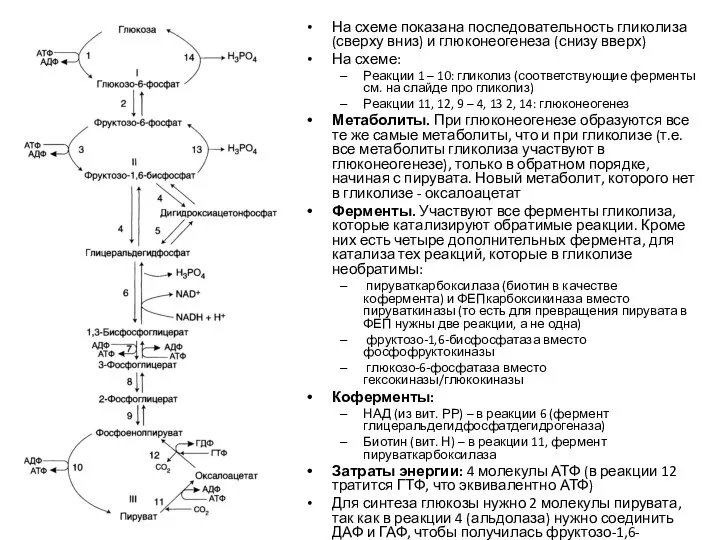
На схеме показана последовательность гликолиза (сверху вниз) и глюконеогенеза (снизу вверх)
На
На схеме показана последовательность гликолиза (сверху вниз) и глюконеогенеза (снизу вверх)
На

ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
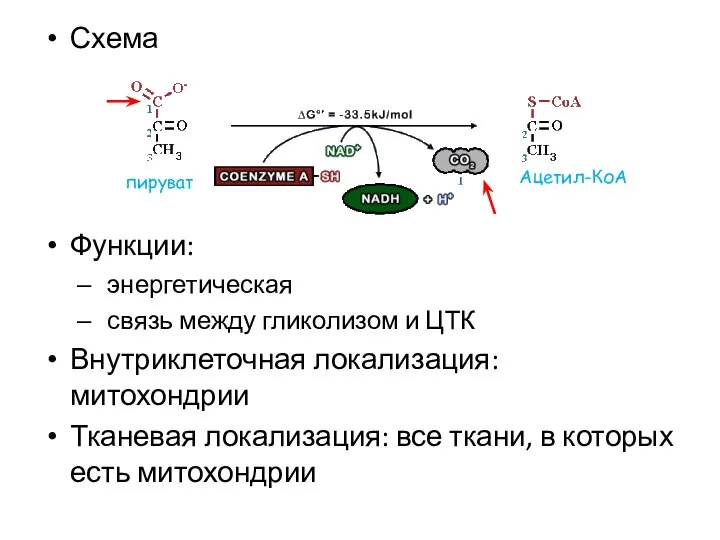
Схема
Функции:
энергетическая
связь между гликолизом и ЦТК
Внутриклеточная локализация: митохондрии
Тканевая локализация: все
Схема
Функции:
энергетическая
связь между гликолизом и ЦТК
Внутриклеточная локализация: митохондрии
Тканевая локализация: все

В результате реакции от пирувата отщепляется карбоксильная группа, которая уходит в
В результате реакции от пирувата отщепляется карбоксильная группа, которая уходит в

ЦИКЛ КРЕБСА (ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ)
ЦИКЛ КРЕБСА (ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ)
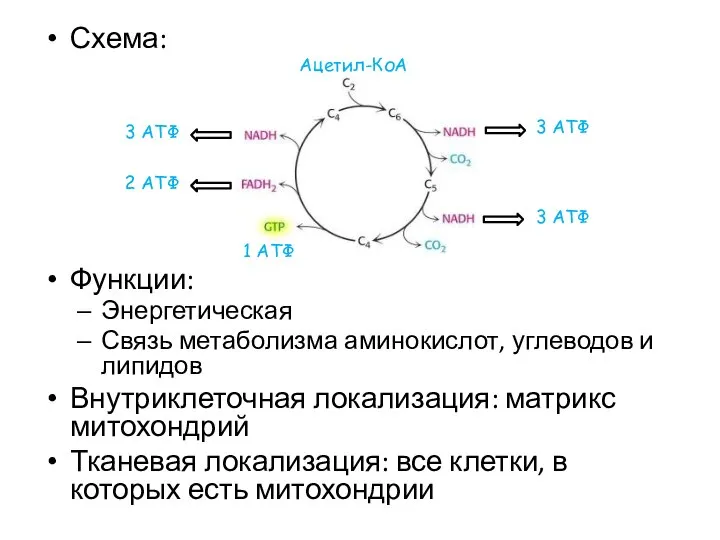
Схема:
Функции:
Энергетическая
Связь метаболизма аминокислот, углеводов и липидов
Внутриклеточная локализация: матрикс митохондрий
Тканевая локализация: все
Схема:
Функции:
Энергетическая
Связь метаболизма аминокислот, углеводов и липидов
Внутриклеточная локализация: матрикс митохондрий
Тканевая локализация: все
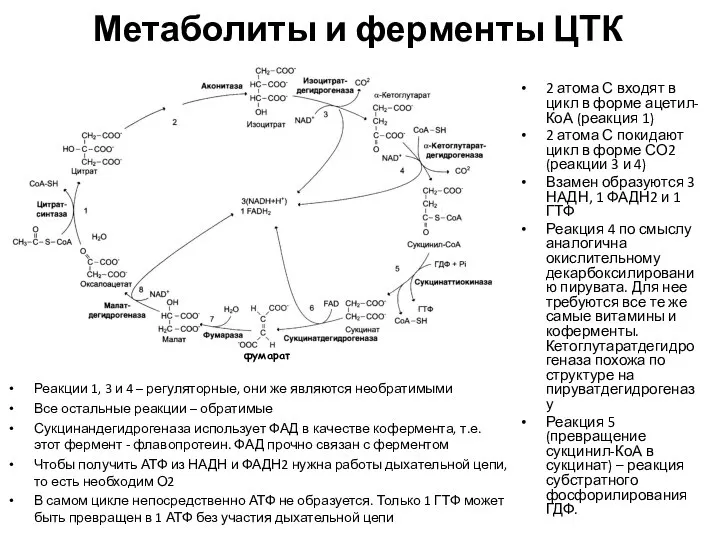
Метаболиты и ферменты ЦТК
2 атома С входят в цикл в форме
Метаболиты и ферменты ЦТК
2 атома С входят в цикл в форме

ДЫХАТЕЛЬНАЯ ЦЕПЬ И ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
ДЫХАТЕЛЬНАЯ ЦЕПЬ И ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
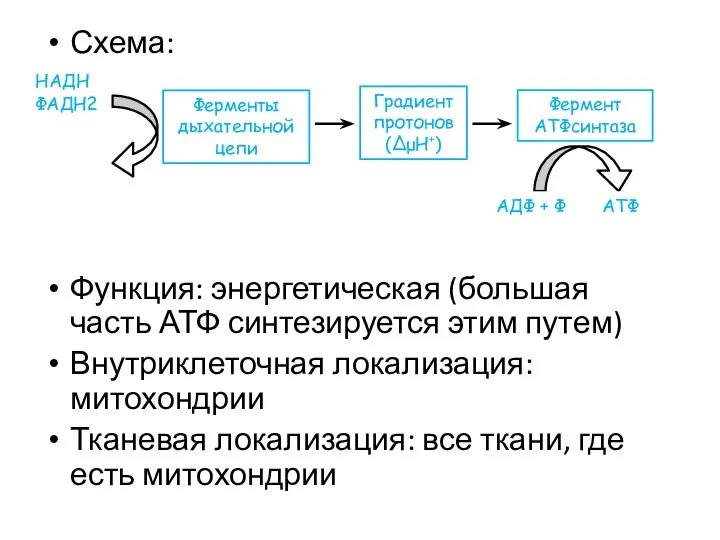
Схема:
Функция: энергетическая (большая часть АТФ синтезируется этим путем)
Внутриклеточная локализация: митохондрии
Тканевая локализация:
Схема:
Функция: энергетическая (большая часть АТФ синтезируется этим путем)
Внутриклеточная локализация: митохондрии
Тканевая локализация:
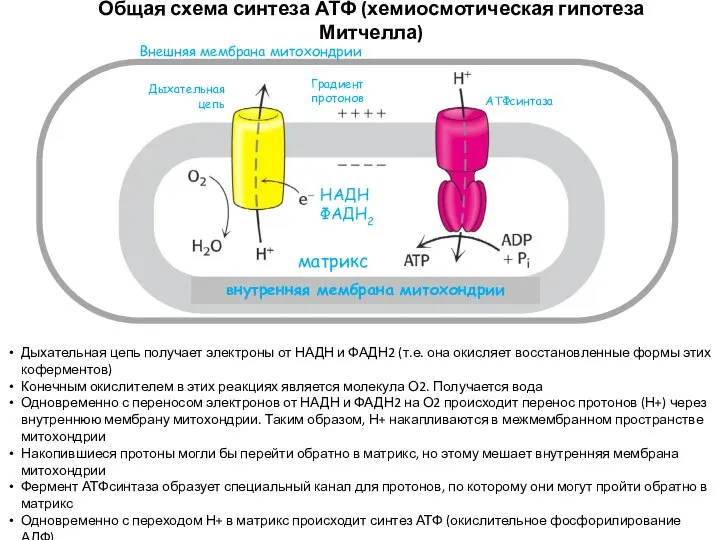
Общая схема синтеза АТФ (хемиосмотическая гипотеза Митчелла)
НАДН
ФАДН2
Дыхательная цепь
Градиент протонов
АТФсинтаза
Внешняя мембрана митохондрии
внутренняя
Общая схема синтеза АТФ (хемиосмотическая гипотеза Митчелла)
НАДН
ФАДН2
Дыхательная цепь
Градиент протонов
АТФсинтаза
Внешняя мембрана митохондрии
внутренняя
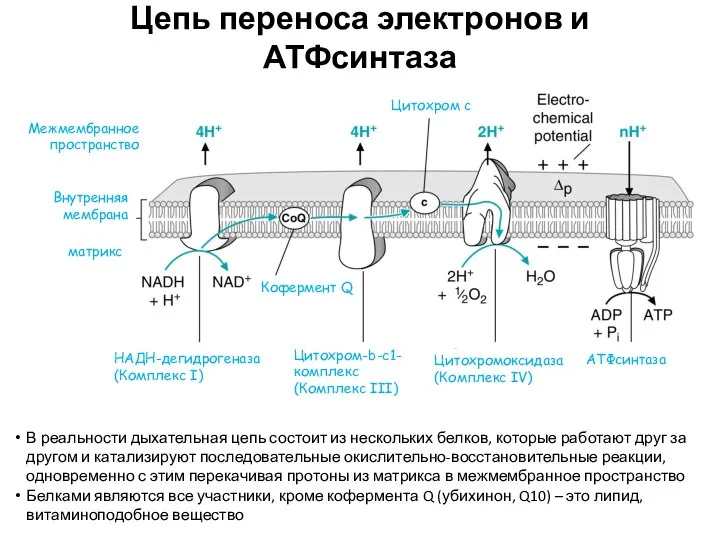
Цепь переноса электронов и АТФсинтаза
В реальности дыхательная цепь состоит из нескольких
Цепь переноса электронов и АТФсинтаза
В реальности дыхательная цепь состоит из нескольких

Подробное описание работы дыхательной цепи
Комплекс I окисляет НАДН и восстанавливается (и
Подробное описание работы дыхательной цепи
Комплекс I окисляет НАДН и восстанавливается (и

ГЛИКОГЕН: СТРУКТУРА, СИНТЕЗ, РАСПАД
ГЛИКОГЕН: СТРУКТУРА, СИНТЕЗ, РАСПАД
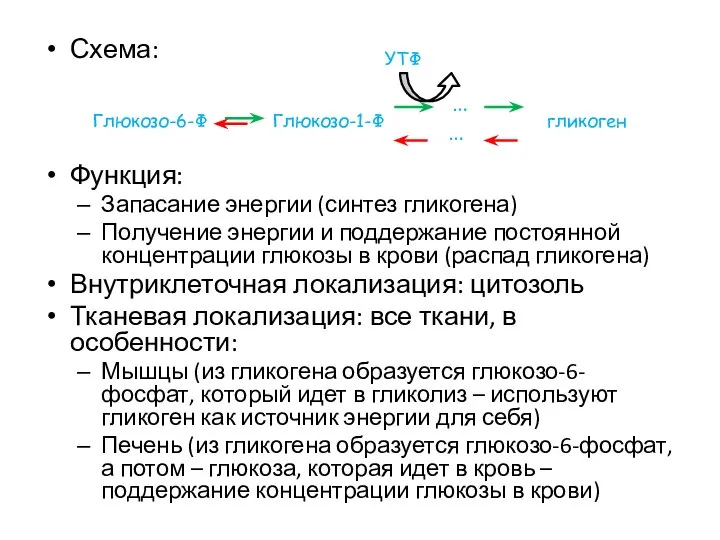
Схема:
Функция:
Запасание энергии (синтез гликогена)
Получение энергии и поддержание постоянной концентрации глюкозы в
Схема:
Функция:
Запасание энергии (синтез гликогена)
Получение энергии и поддержание постоянной концентрации глюкозы в
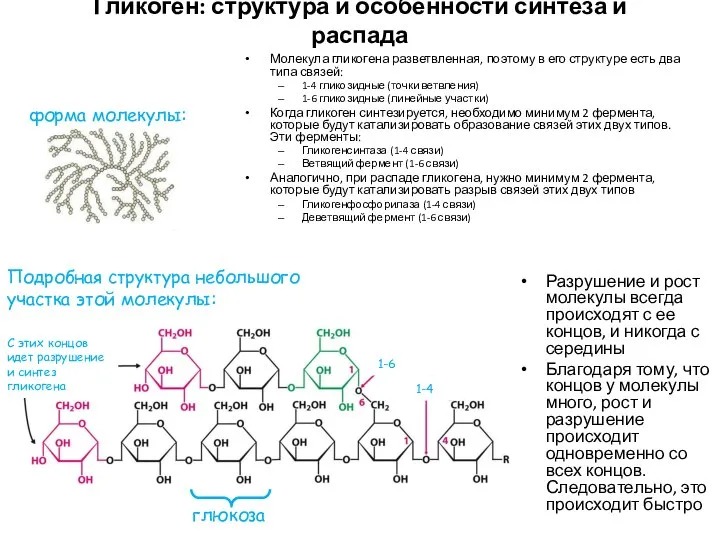
Гликоген: структура и особенности синтеза и распада
Молекула гликогена разветвленная, поэтому в
Гликоген: структура и особенности синтеза и распада
Молекула гликогена разветвленная, поэтому в
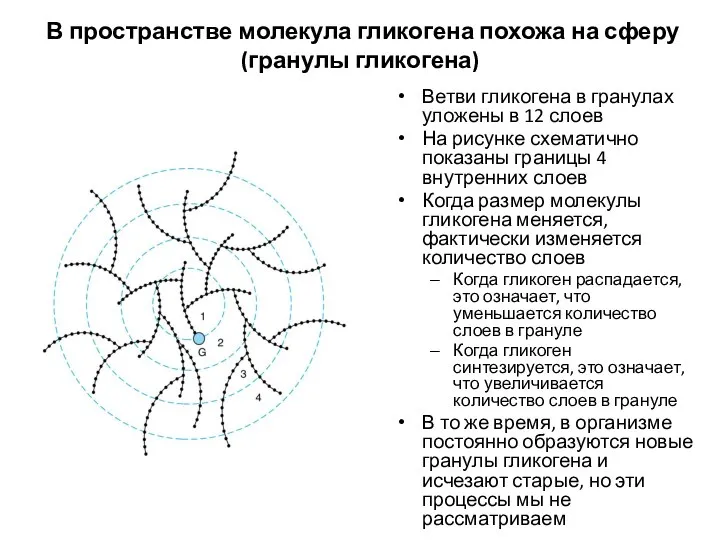
В пространстве молекула гликогена похожа на сферу (гранулы гликогена)
Ветви гликогена в
В пространстве молекула гликогена похожа на сферу (гранулы гликогена)
Ветви гликогена в

Этапы синтеза гликогена
В клетке всегда существуют небольшие молекулы гликогена, имеющие несколько
Этапы синтеза гликогена
В клетке всегда существуют небольшие молекулы гликогена, имеющие несколько
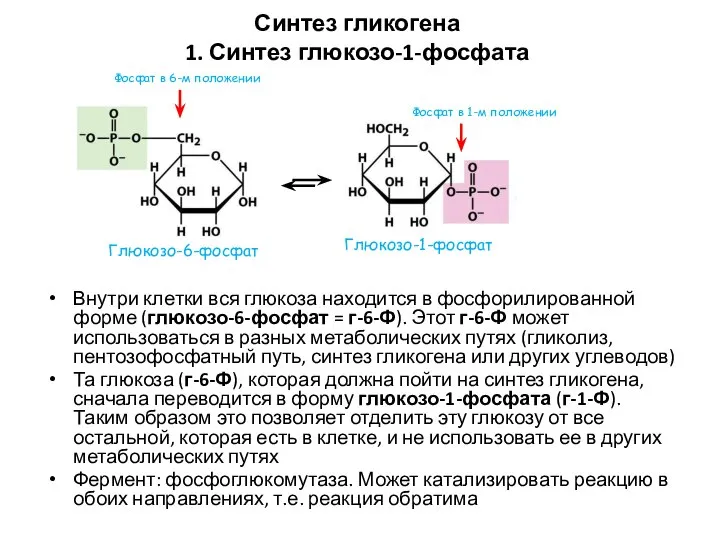
Синтез гликогена
1. Синтез глюкозо-1-фосфата
Внутри клетки вся глюкоза находится в фосфорилированной форме
Синтез гликогена
1. Синтез глюкозо-1-фосфата
Внутри клетки вся глюкоза находится в фосфорилированной форме
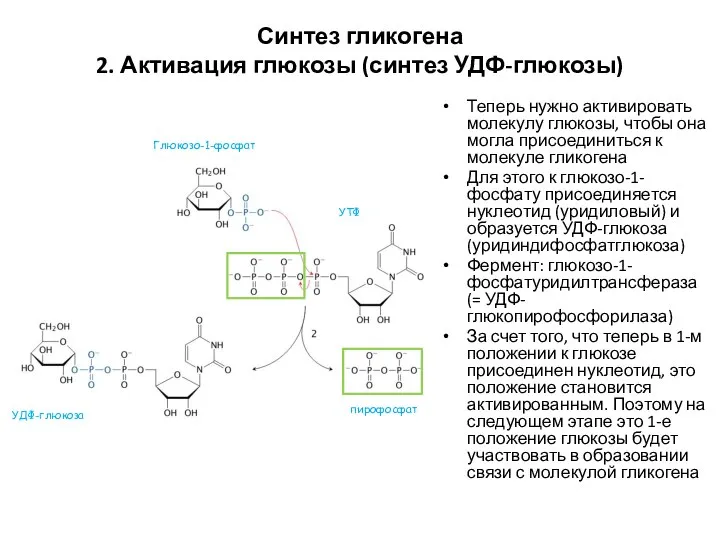
Синтез гликогена
2. Активация глюкозы (синтез УДФ-глюкозы)
Теперь нужно активировать молекулу глюкозы, чтобы
Синтез гликогена
2. Активация глюкозы (синтез УДФ-глюкозы)
Теперь нужно активировать молекулу глюкозы, чтобы
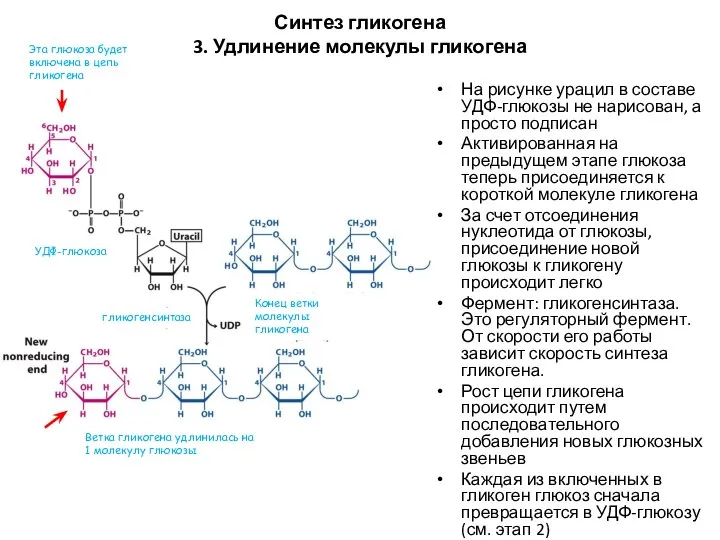
Синтез гликогена
3. Удлинение молекулы гликогена
На рисунке урацил в составе УДФ-глюкозы не
Синтез гликогена
3. Удлинение молекулы гликогена
На рисунке урацил в составе УДФ-глюкозы не
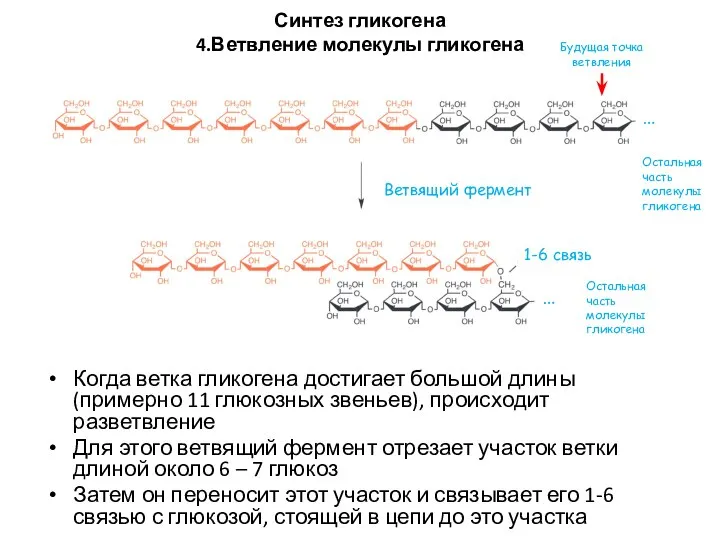
Синтез гликогена
4.Ветвление молекулы гликогена
Когда ветка гликогена достигает большой длины (примерно
Синтез гликогена
4.Ветвление молекулы гликогена
Когда ветка гликогена достигает большой длины (примерно

Этапы распада гликогена
Ферменты укорачивают отдельные ветки молекул гликогена или удаляют некоторые
Этапы распада гликогена
Ферменты укорачивают отдельные ветки молекул гликогена или удаляют некоторые
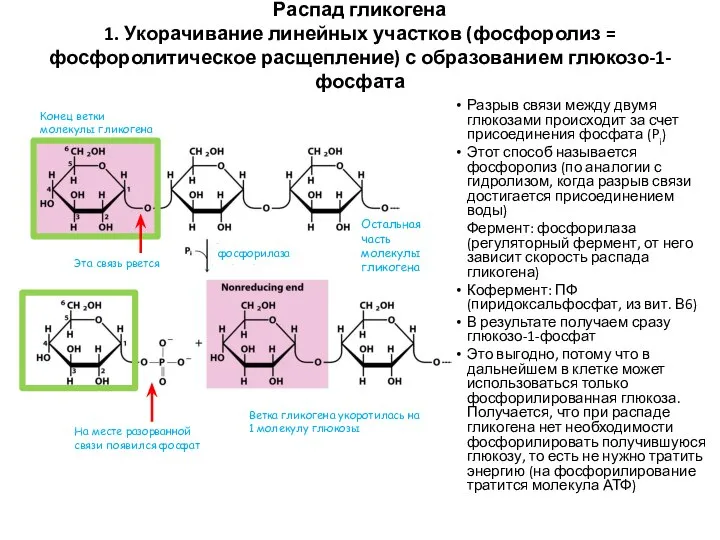
Распад гликогена
1. Укорачивание линейных участков (фосфоролиз = фосфоролитическое расщепление) с образованием
Распад гликогена 1. Укорачивание линейных участков (фосфоролиз = фосфоролитическое расщепление) с образованием
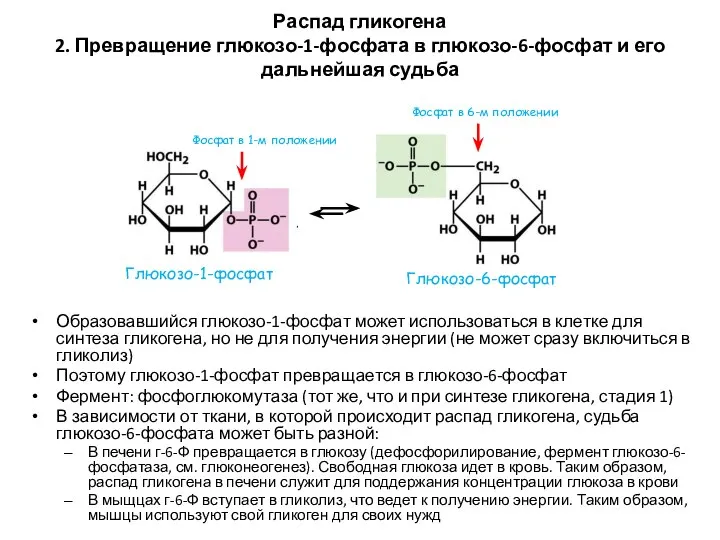
Распад гликогена
2. Превращение глюкозо-1-фосфата в глюкозо-6-фосфат и его дальнейшая судьба
Образовавшийся глюкозо-1-фосфат
Распад гликогена
2. Превращение глюкозо-1-фосфата в глюкозо-6-фосфат и его дальнейшая судьба
Образовавшийся глюкозо-1-фосфат
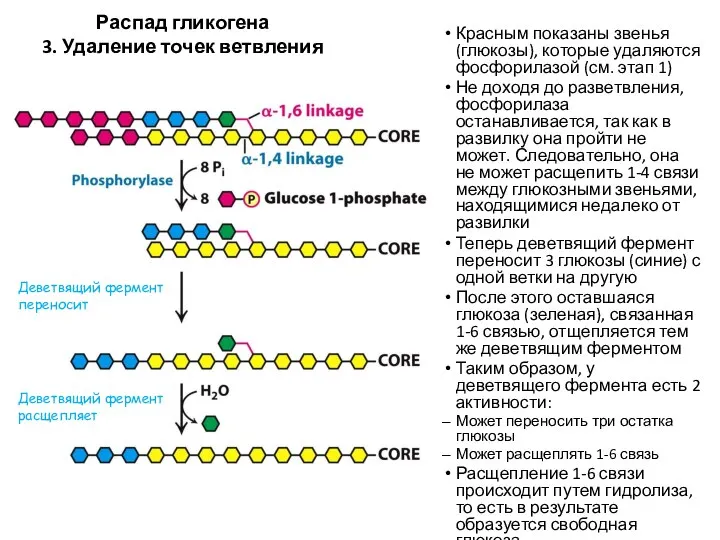
Распад гликогена
3. Удаление точек ветвления
Красным показаны звенья (глюкозы), которые удаляются фосфорилазой
Распад гликогена
3. Удаление точек ветвления
Красным показаны звенья (глюкозы), которые удаляются фосфорилазой

Нарушения метаболизма гликогена - гликогенозы
Ферменты, участвующие в синтезе и распаде гликогена,
Нарушения метаболизма гликогена - гликогенозы
Ферменты, участвующие в синтезе и распаде гликогена,

ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
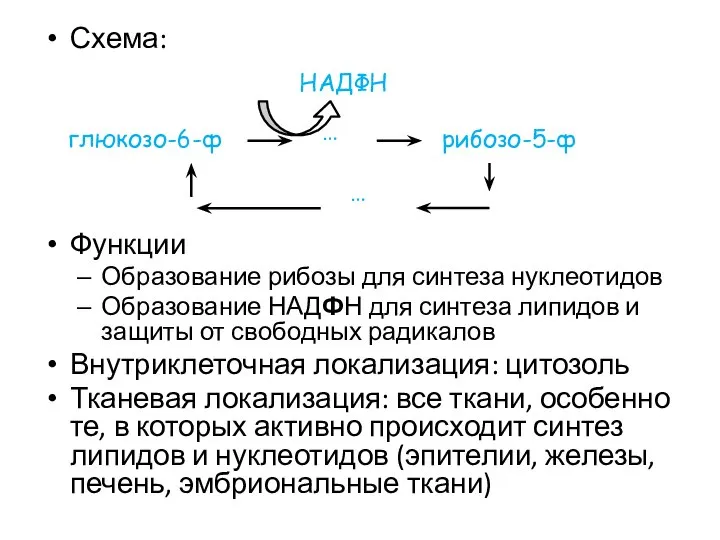
Схема:
Функции
Образование рибозы для синтеза нуклеотидов
Образование НАДФН для синтеза липидов и защиты
Схема:
Функции
Образование рибозы для синтеза нуклеотидов
Образование НАДФН для синтеза липидов и защиты
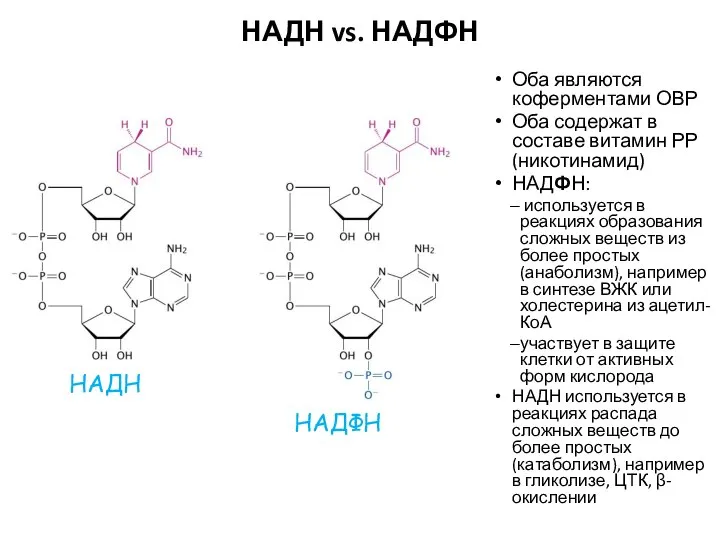
НАДН vs. НАДФН
Оба являются коферментами ОВР
Оба содержат в составе витамин РР
НАДН vs. НАДФН
Оба являются коферментами ОВР
Оба содержат в составе витамин РР

Две стадии:
Окислительная: глюкозо-6-фосфат (С6) превращается в рибулозо-5-фосфат (С5), образуются 2 НАДФН
Неокислительная:
Две стадии:
Окислительная: глюкозо-6-фосфат (С6) превращается в рибулозо-5-фосфат (С5), образуются 2 НАДФН
Неокислительная:
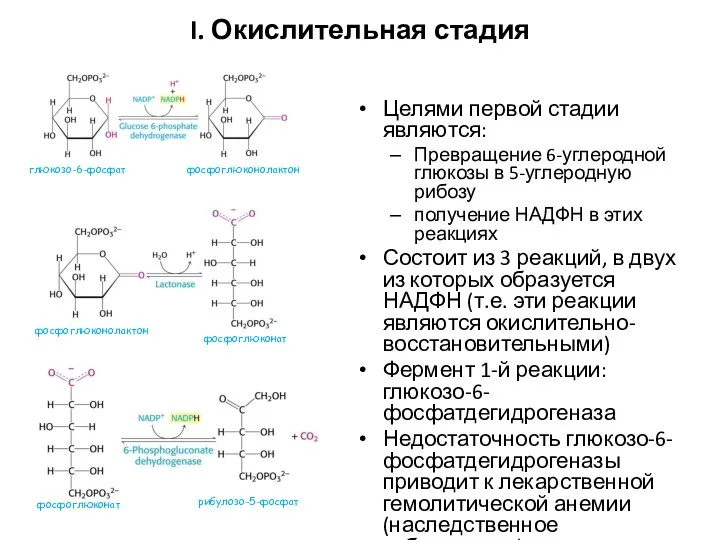
I. Окислительная стадия
Целями первой стадии являются:
Превращение 6-углеродной глюкозы в 5-углеродную рибозу
получение
I. Окислительная стадия
Целями первой стадии являются:
Превращение 6-углеродной глюкозы в 5-углеродную рибозу
получение
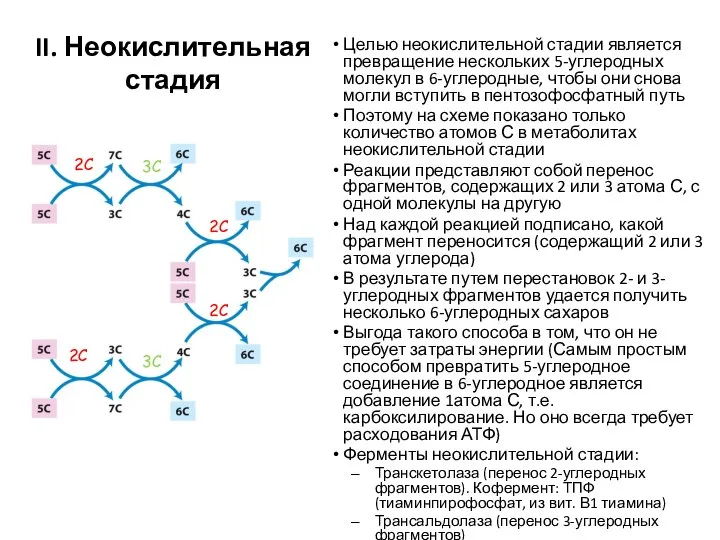
Целью неокислительной стадии является превращение нескольких 5-углеродных молекул в 6-углеродные, чтобы
Целью неокислительной стадии является превращение нескольких 5-углеродных молекул в 6-углеродные, чтобы

Полная схема пентозофосфатного пути
Обратите внимание на метаболиты неокислительной стадии: среди них
Полная схема пентозофосфатного пути
Обратите внимание на метаболиты неокислительной стадии: среди них

Нарушения пентозофосфатного пути
Наследственное заболевание, связанное с нарушением работы пентозофосфатного пути: лекарственная
Нарушения пентозофосфатного пути
Наследственное заболевание, связанное с нарушением работы пентозофосфатного пути: лекарственная

Участие глутатиона и пентозофосфатного пути в защите от свободных радикалов
Глутатионпероксидаза при
Участие глутатиона и пентозофосфатного пути в защите от свободных радикалов
Глутатионпероксидаза при

РЕГУЛЯЦИЯ МЕТАБОЛИЗМА УГЛЕВОДОВ
РЕГУЛЯЦИЯ МЕТАБОЛИЗМА УГЛЕВОДОВ

Регуляторные ферменты
В любом метаболическом пути есть два типа ферментов (с точки
Регуляторные ферменты
В любом метаболическом пути есть два типа ферментов (с точки
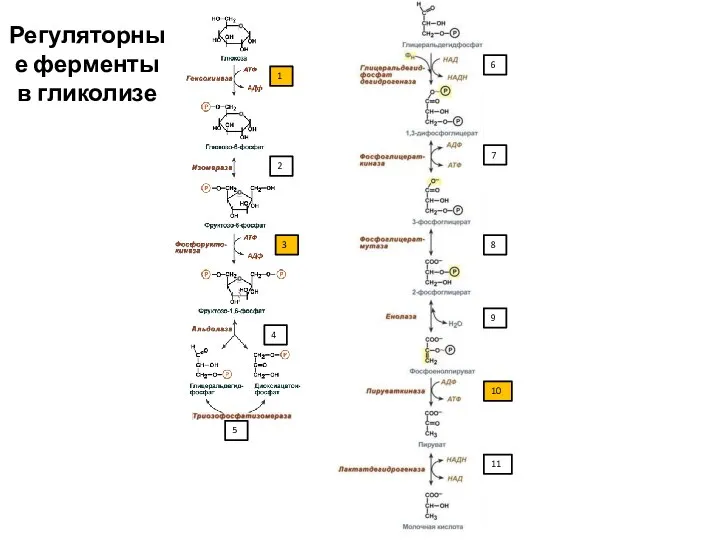
1
2
3
4
5
6
7
8
9
10
11
Регуляторные ферменты в гликолизе
1
2
3
4
5
6
7
8
9
10
11
Регуляторные ферменты в гликолизе

Общие принципы регуляции работы ферментов
Изменение скорости работы ферментов происходит путем их:
Общие принципы регуляции работы ферментов
Изменение скорости работы ферментов происходит путем их:

Регуляторные ферменты гликолиза
Необратимые реакции (они же регуляторные = ключевые = скорость-лимитирующие
Регуляторные ферменты гликолиза
Необратимые реакции (они же регуляторные = ключевые = скорость-лимитирующие

Аллостерические регуляторы гликолиза
Тормозят гликолиз: АТФ, цитрат, глюкозо-6-фосфат (мышцы), фруктозо-6-фосфат (печень), ацетил-КоА,
Аллостерические регуляторы гликолиза
Тормозят гликолиз: АТФ, цитрат, глюкозо-6-фосфат (мышцы), фруктозо-6-фосфат (печень), ацетил-КоА,

Гормональная регуляция гликолиза
Тормозит: глюкагон
Активируют: инсулин, адреналин (в мышцах)
Гормональная регуляция гликолиза
Тормозит: глюкагон
Активируют: инсулин, адреналин (в мышцах)
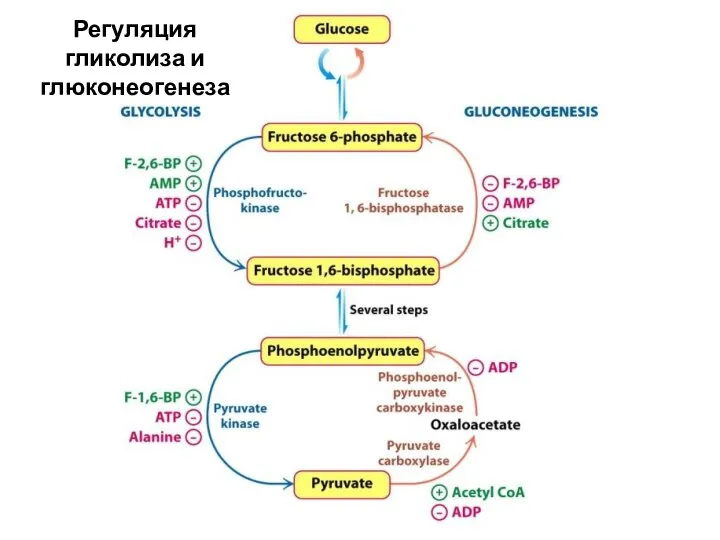
Регуляция гликолиза и глюконеогенеза
Регуляция гликолиза и глюконеогенеза

РЕГУЛЯЦИЯ СИНТЕЗА И РАСПАДА ГЛИКОГЕНА
РЕГУЛЯЦИЯ СИНТЕЗА И РАСПАДА ГЛИКОГЕНА

Регуляторные ферменты
Гликогенсинтаза – регуляторный фермент синтеза гликогена. Неактивна в фосфорилированной форме,
Регуляторные ферменты
Гликогенсинтаза – регуляторный фермент синтеза гликогена. Неактивна в фосфорилированной форме,
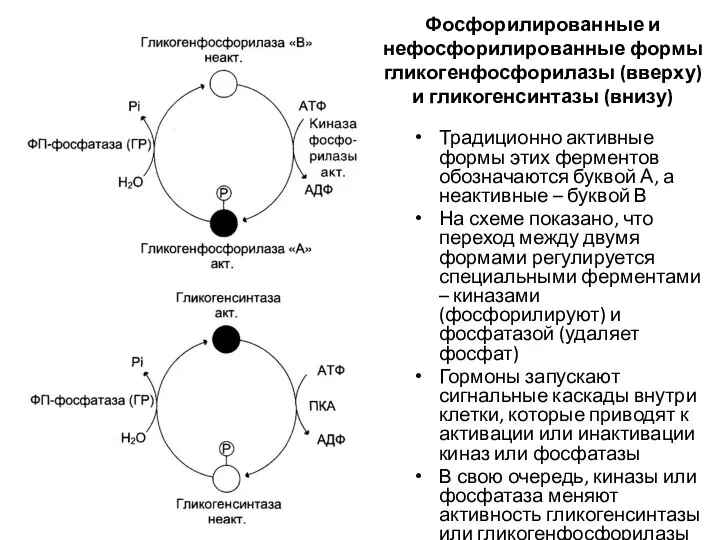
Фосфорилированные и нефосфорилированные формы гликогенфосфорилазы (вверху) и гликогенсинтазы (внизу)
Традиционно активные формы
Фосфорилированные и нефосфорилированные формы гликогенфосфорилазы (вверху) и гликогенсинтазы (внизу)
Традиционно активные формы
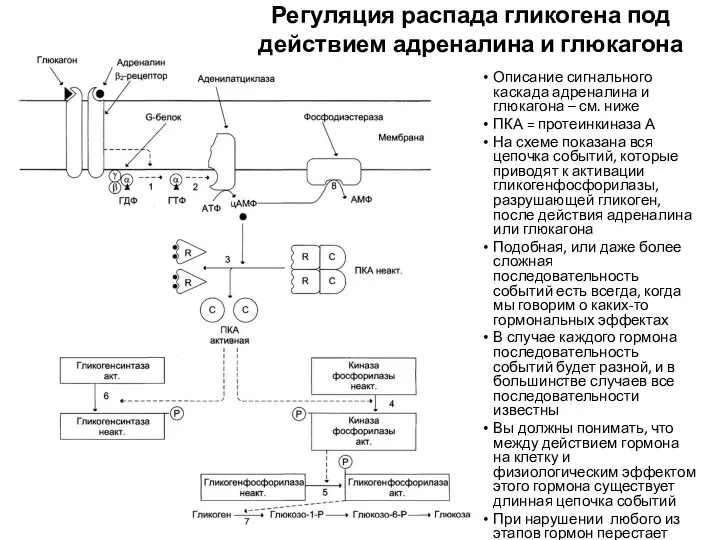
Описание сигнального каскада адреналина и глюкагона – см. ниже
ПКА = протеинкиназа
Описание сигнального каскада адреналина и глюкагона – см. ниже
ПКА = протеинкиназа

РЕГУЛЯЦИЯ ЦТК
РЕГУЛЯЦИЯ ЦТК

Регуляторные ферменты
Цитратсинтаза:
Ингибируют: НАДН, сукцинил-КоА, цитрат, АТФ
Активирует: АДФ
Изоцитратдегидрогеназа
Ингибирует: АТФ
Активируют: Ca2+, АДФ
α-кетоглутаратдегидрогеназный комплекс
Ингибируют:
Регуляторные ферменты
Цитратсинтаза:
Ингибируют: НАДН, сукцинил-КоА, цитрат, АТФ
Активирует: АДФ
Изоцитратдегидрогеназа
Ингибирует: АТФ
Активируют: Ca2+, АДФ
α-кетоглутаратдегидрогеназный комплекс
Ингибируют:


ФИЗИОЛОГИЧЕСКИЕ ЭФФЕКТЫ ИНСУЛИНА: МЕТАБОЛИЗМ ГЛЮКОЗЫ, АМИНОКИСЛОТ, ЛИПИДОВ
ФИЗИОЛОГИЧЕСКИЕ ЭФФЕКТЫ ИНСУЛИНА: МЕТАБОЛИЗМ ГЛЮКОЗЫ, АМИНОКИСЛОТ, ЛИПИДОВ

Влияние на метаболизм углеводов
Активация поступления глюкозы в клетки, снижение глюкозы в
Влияние на метаболизм углеводов
Активация поступления глюкозы в клетки, снижение глюкозы в

Влияние инсулина на концентрацию глюкозы в крови
После еду глюкоза поступает в
Влияние инсулина на концентрацию глюкозы в крови
После еду глюкоза поступает в
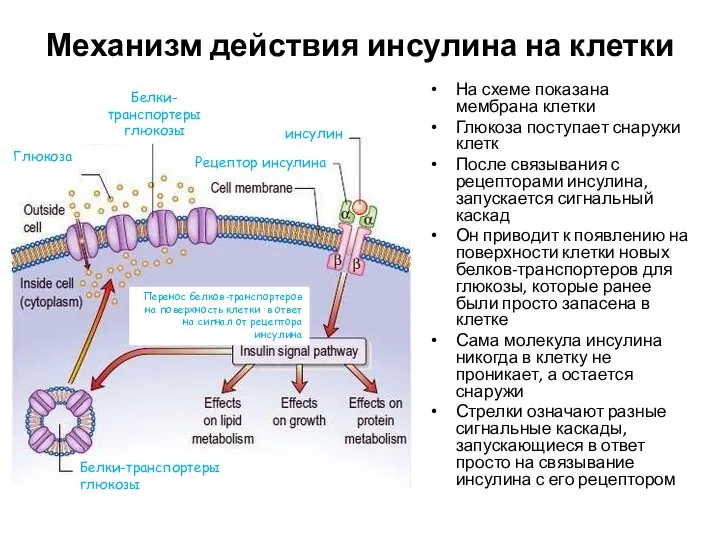
Механизм действия инсулина на клетки
На схеме показана мембрана клетки
Глюкоза поступает снаружи
Механизм действия инсулина на клетки
На схеме показана мембрана клетки
Глюкоза поступает снаружи

Влияние на метаболизм липидов
Ингибирует гормон-чувствительную липазу адипоцитов
Активирует липогенез (синтез липидов) в
Влияние на метаболизм липидов
Ингибирует гормон-чувствительную липазу адипоцитов
Активирует липогенез (синтез липидов) в

Влияние на метаболизм белков и аминокислот
Активация синтеза белков
Активация захвата аминокислот тканями
Влияние на метаболизм белков и аминокислот
Активация синтеза белков
Активация захвата аминокислот тканями

САХАРНЫЙ ДИАБЕТ: ПРИЧИНЫ И ТИПЫ
САХАРНЫЙ ДИАБЕТ: ПРИЧИНЫ И ТИПЫ

СД 1 типа
Иммунные: аутоантитела к инсулину и специфическим белкам бета-клеток. Разрушение
СД 1 типа
Иммунные: аутоантитела к инсулину и специфическим белкам бета-клеток. Разрушение

СД 2 типа
Инсулинорезистентность – невосприимчивость (устойчивость) клеток к сигналу, который им
СД 2 типа
Инсулинорезистентность – невосприимчивость (устойчивость) клеток к сигналу, который им

ГЛЮКАГОН И АДРЕНАЛИН
ГЛЮКАГОН И АДРЕНАЛИН

Глюкагон и адреналин
Глюкагон – пептидный гормон
Адреналин – производное аминокислоты тирозина
Оба гормона
Глюкагон и адреналин
Глюкагон – пептидный гормон
Адреналин – производное аминокислоты тирозина
Оба гормона

ИНТЕГРАЦИЯ МЕТАБОЛИЗМА
ИНТЕГРАЦИЯ МЕТАБОЛИЗМА

«Метаболическая воронка»
Это понятие, которое описывает унификацию получения энергии из разных источников
Энергию
«Метаболическая воронка»
Это понятие, которое описывает унификацию получения энергии из разных источников
Энергию
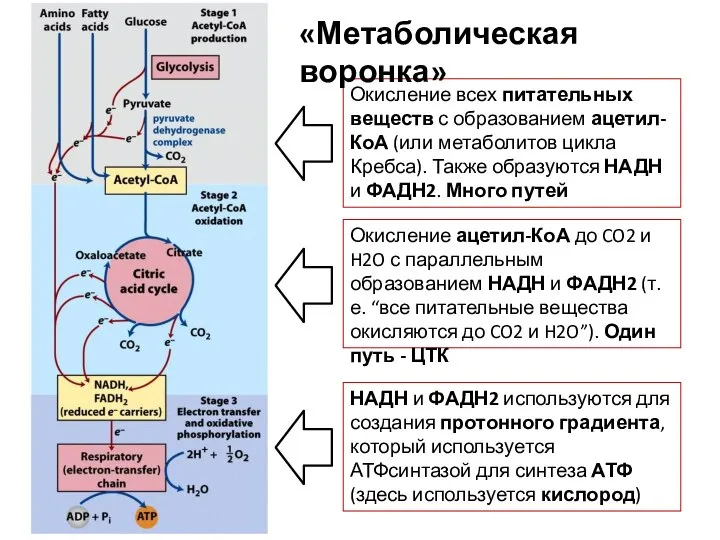
Окисление всех питательных веществ с образованием ацетил-КоА (или метаболитов цикла Кребса).
Окисление всех питательных веществ с образованием ацетил-КоА (или метаболитов цикла Кребса).
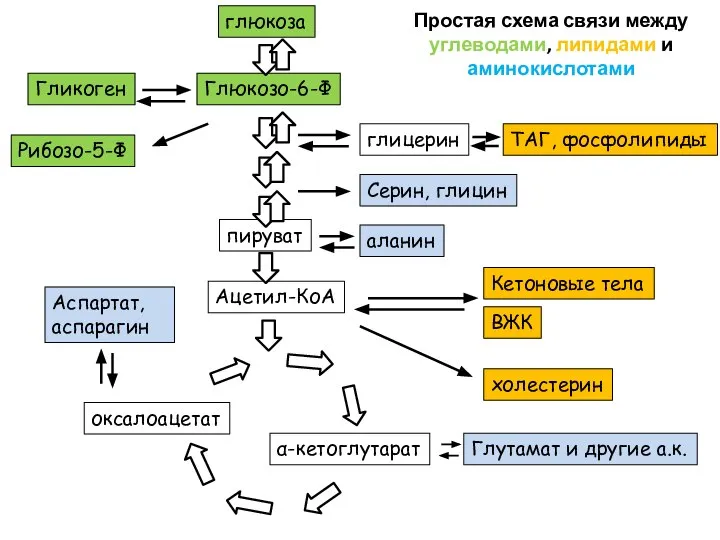
Ацетил-КоА
пируват
глюкоза
Серин, глицин
Глюкозо-6-Ф
Кетоновые тела
ВЖК
холестерин
α-кетоглутарат
Глутамат и другие а.к.
оксалоацетат
Аспартат, аспарагин
аланин
Гликоген
Рибозо-5-Ф
глицерин
ТАГ, фосфолипиды
Простая схема связи между
Ацетил-КоА
пируват
глюкоза
Серин, глицин
Глюкозо-6-Ф
Кетоновые тела
ВЖК
холестерин
α-кетоглутарат
Глутамат и другие а.к.
оксалоацетат
Аспартат, аспарагин
аланин
Гликоген
Рибозо-5-Ф
глицерин
ТАГ, фосфолипиды
Простая схема связи между
 ИНТЕРЕСНЫЕ ФАКТЫ ПРО ЖИВОТНЫХ
ИНТЕРЕСНЫЕ ФАКТЫ ПРО ЖИВОТНЫХ МЫШЦЫ. РАБОТА МЫШЦ. Подготовила учитель биологии и химии – Протопопова Оксана Евгеньевна
МЫШЦЫ. РАБОТА МЫШЦ. Подготовила учитель биологии и химии – Протопопова Оксана Евгеньевна Кровеносная система человека
Кровеносная система человека Беременность и роды. Изменения в организме самок во время беременности
Беременность и роды. Изменения в организме самок во время беременности Анализаторы. Сенсорная система
Анализаторы. Сенсорная система Обмен веществ и энергии – основное свойство живых существ (8 класс)
Обмен веществ и энергии – основное свойство живых существ (8 класс) Селекция ореха грецкого
Селекция ореха грецкого Исследование внутренних факторов: площади испаряющей поверхности и отношения этой поверхности к объему растений
Исследование внутренних факторов: площади испаряющей поверхности и отношения этой поверхности к объему растений Динамика популяций
Динамика популяций Структурно-функциональная организация половой системы самцов
Структурно-функциональная организация половой системы самцов Введение в геологию
Введение в геологию Строение и функционирование пищеварительной системы рыб
Строение и функционирование пищеварительной системы рыб Разнообразие растительного мира. Урок 29
Разнообразие растительного мира. Урок 29 Проект на тему Мурахи
Проект на тему Мурахи Чукотское море Подготовил: Ученик 8 класса Капустин Сергей
Чукотское море Подготовил: Ученик 8 класса Капустин Сергей  Животные в природных сообществах
Животные в природных сообществах Домовой воробей- птица 2022
Домовой воробей- птица 2022 Плесневые грибы. Дрожжи
Плесневые грибы. Дрожжи Презентация на тему Обоняние и вкус
Презентация на тему Обоняние и вкус  Різноманітність голонасінних, їх значення
Різноманітність голонасінних, їх значення Медицинская гельминтология. Ленточные черви – Cestodae
Медицинская гельминтология. Ленточные черви – Cestodae Презентация на тему "Плоды биология" - скачать презентации по Биологии
Презентация на тему "Плоды биология" - скачать презентации по Биологии Кинология − наука о собаках
Кинология − наука о собаках Презентация по биологии Водоросли
Презентация по биологии Водоросли Породы кроликов
Породы кроликов Нервная ткань: нейроциты, глиоциты, нервные волокн. Тема 12
Нервная ткань: нейроциты, глиоциты, нервные волокн. Тема 12 Дятлы
Дятлы Многоклеточные организмы без настоящих тканей
Многоклеточные организмы без настоящих тканей