- Молекулярная биология ч.4 Мессенджеры

Содержание
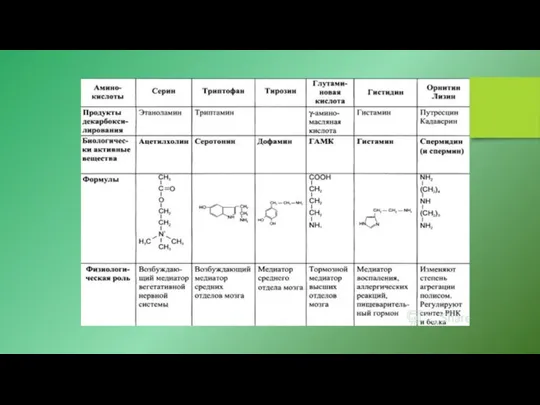
- 10. мембранные рецепторы 5-гидрокситриптамина (5-HT), нейромедиатора и гормона, известного под названием серотонин, взаимодействующие также с множеством медицинских
- 19. 1. Связывание агониста индуцирует открывание канала, образуемого рецептором. Примерами могут служить никотиновый ацетилхолиновый рецептор (nAChR), рецептор
- 21. Примеры молекулярной организации некоторых структур, участвующих в путях сигнальной трансдукции. А - рецептор мембраны клетки, связывающий
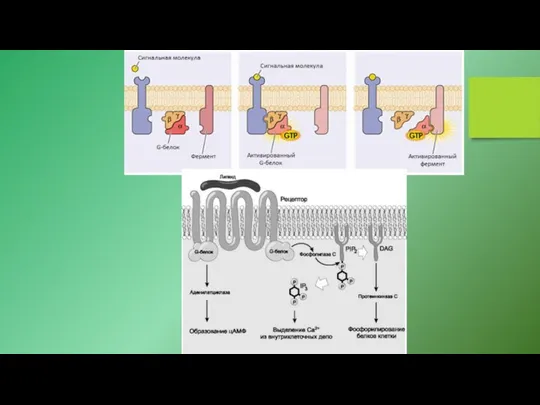
- 22. G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим
- 23. Принцип работы гетеротримерных ГТФ-связывающих белков (гетеротримерных G-белков) Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).
- 26. Скачать презентацию





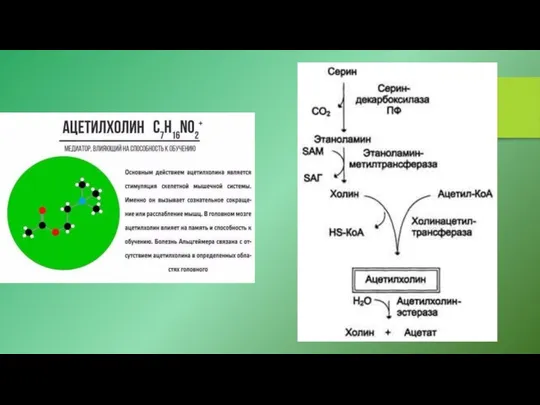
мембранные рецепторы 5-гидрокситриптамина (5-HT), нейромедиатора и гормона, известного под названием серотонин,
мембранные рецепторы 5-гидрокситриптамина (5-HT), нейромедиатора и гормона, известного под названием серотонин,
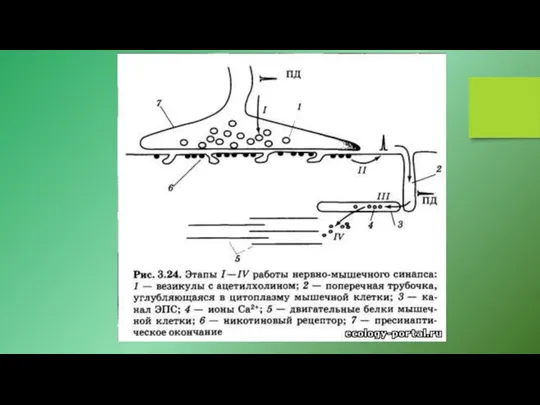

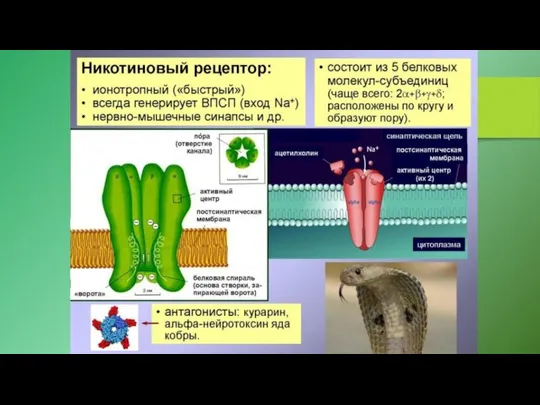


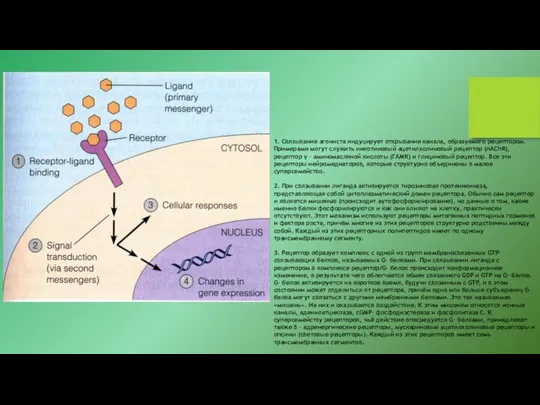
1. Связывание агониста индуцирует открывание канала, образуемого рецептором. Примерами могут служить
1. Связывание агониста индуцирует открывание канала, образуемого рецептором. Примерами могут служить
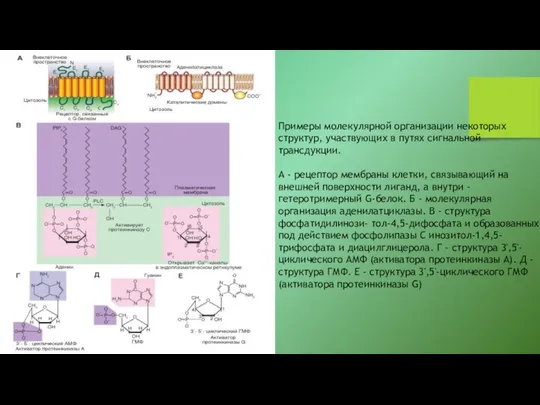
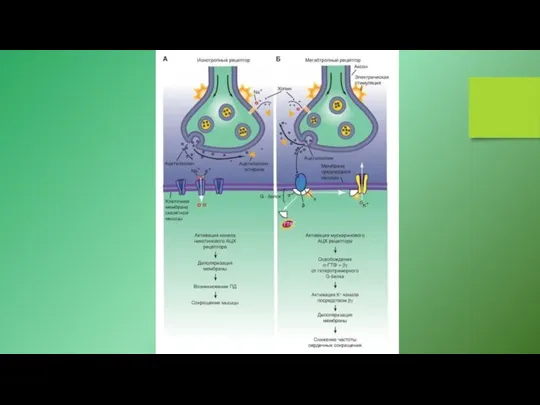
Примеры молекулярной организации некоторых структур, участвующих в путях сигнальной трансдукции.
А -
Примеры молекулярной организации некоторых структур, участвующих в путях сигнальной трансдукции.
А -

G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов
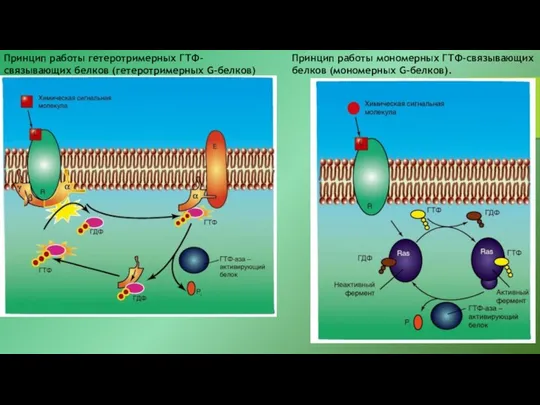
Принцип работы гетеротримерных ГТФ-связывающих белков (гетеротримерных G-белков)
Принцип работы мономерных ГТФ-связывающих белков
Принцип работы гетеротримерных ГТФ-связывающих белков (гетеротримерных G-белков)
Принцип работы мономерных ГТФ-связывающих белков

 Влияние густоты посева семян на урожай моркови Автор проекта: ученик 1 «В» класса Гимназии №1 Баженов Дмитрий Руководитель: Крае
Влияние густоты посева семян на урожай моркови Автор проекта: ученик 1 «В» класса Гимназии №1 Баженов Дмитрий Руководитель: Крае Презентация на тему "Как живут растения?" - скачать презентации по Биологии
Презентация на тему "Как живут растения?" - скачать презентации по Биологии Память
Память L’écureuil et sa famille adoptive
L’écureuil et sa famille adoptive Природа как ценность. Антропоцентризм и натуроцентризм
Природа как ценность. Антропоцентризм и натуроцентризм Генетические закономерности
Генетические закономерности Презентация на тему "Чарльз Роберт Дарвин" - скачать презентации по Биологии
Презентация на тему "Чарльз Роберт Дарвин" - скачать презентации по Биологии Презентация Репликация ДНК
Презентация Репликация ДНК Изучение повреждения молекулы ДНК в присутствии противоопухолевых препаратов, индуцированных электронным пучком
Изучение повреждения молекулы ДНК в присутствии противоопухолевых препаратов, индуцированных электронным пучком Морские жители
Морские жители Презентация на тему "Необычные деревья" - скачать презентации по Биологии
Презентация на тему "Необычные деревья" - скачать презентации по Биологии Презентация на тему Мендель Грегор Иоганн
Презентация на тему Мендель Грегор Иоганн Презентация к уроку по теме : Класс Паукообразные работу выполнила учитель Авдеевской СОШ Зарайского района Моско
Презентация к уроку по теме : Класс Паукообразные работу выполнила учитель Авдеевской СОШ Зарайского района Моско Презентация на тему "Строение сердца. Сердечный цикл." - скачать бесплатно презентации по Биологии
Презентация на тему "Строение сердца. Сердечный цикл." - скачать бесплатно презентации по Биологии Презентация Анти-спид
Презентация Анти-спид Скелет рыб Урок 7 класс. Учитель Еременко Е. Ю.
Скелет рыб Урок 7 класс. Учитель Еременко Е. Ю. Гормони. Молекулярні механізми передачі гормонального сигналу
Гормони. Молекулярні механізми передачі гормонального сигналу Основы учения об эволюции. Эволюционная теория Ч. Дарвина
Основы учения об эволюции. Эволюционная теория Ч. Дарвина Доказательства происхождения человека от животных
Доказательства происхождения человека от животных Таламус. Горизонтальный и фронтальный срезы
Таламус. Горизонтальный и фронтальный срезы Радиация в биосфере
Радиация в биосфере Коралловые полипы О коралловых полипах. Выполнил: Убишев Данияр. Учитель: Кадырова Т.М.
Коралловые полипы О коралловых полипах. Выполнил: Убишев Данияр. Учитель: Кадырова Т.М.  Живородящая ящерица и её содержание в неволе
Живородящая ящерица и её содержание в неволе Презентация на тему Закон Биогенетический
Презентация на тему Закон Биогенетический  Природоохранительная акция Покормите птиц зимой
Природоохранительная акция Покормите птиц зимой Хозяйственно-биологические особенности птиц
Хозяйственно-биологические особенности птиц Железы внешней, внутренней и смешанной секреции
Железы внешней, внутренней и смешанной секреции  Органы и системы органов животных
Органы и системы органов животных