- Репликация ДНК. (Лекция 3)

Содержание
- 2. План лекции: Механизмы репликации ДНК Репликационная вилка Современная модель репликации Ферменты репликации Особенности репликации про- и
- 3. Механизмы репликации ДНК Во время репликации каждая из цепей родительской ДНК служит матрицей для синтеза комплементарной
- 4. Механизмы репликации ДНК Три альтернативные гипотезы. 1958 г. Подтверждена гипотеза полуконсервативной репликации (эксперимент Мезельсона-Сталя) Схематически представленные
- 5. Репликационная вилка 1957 г. А.Корнберг обнаружил ДНК-полимеразу I – фермент, катализирующий процесс полимеризации ДНК из нуклеотидов.
- 6. Механизмы репликации ДНК у прокариот Реплицирующуюся ДНК бактерии можно наблюдать в электронном микроскопе, у нее вначале
- 7. Современная модель репликации © http://www.nature.com/encode/#/threads Схема репликации ДНК у E.coli. На панели а схематически показаны белки
- 8. Ферменты репликации Чтобы репликационная вилка могла продвигаться вперёд, вся хромосома впереди неё должна быстро вращаться. Это
- 9. ДНК – полимераза I: репаративный синтез ДНК в двух процессах: вырезает повреждённые участки одной цепи и
- 10. Действие топоизомераз I и II © http://helicase.pbworks.com/w/page/17605720/Tyler-Huff © http://mol-biol4masters.masters.grkraj.org/html/Deoxy_Ribose_Nucleic_Acid4-Super_Coiling.htm
- 11. Типы репликации ДНК © http://biology.ru/course/content/chapter8/section2/paragraph2/theory.html#.VcIFtPPtlBc
- 12. В кольцевой хромосоме репликационный глазок образует Ө-структуру. Ө-механизм
- 13. Если в кольцевой ДНК разрывает одну цепь и свободный 3’-конец наращивается с помощью ДНК-полимеразы, то вновь
- 14. В митохондриальной ДНК наблюдается тип репликации носящий название D-петель (displacement – вытеснение). D-петля © http://genesdev.cshlp.org/content/19/20/2466/F1.expansion.html
- 15. Специфические мембранные белки обеспечивают контакт ДНК с внутренней поверхностью цитоплазматической мембраны. Наиболее типичным является формирование подобного
- 16. А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране в точке (или точках) репликации;
- 17. Нуклеозидной природы Ненуклеозидной природы Ингибирование репликации
- 18. Отличия репликации эукариот В эукариотическом геноме имеется множество точек начала репликации (А-Т-богатых сайтов) и различные формы
- 19. Отличия репликации эукариот Участки эукариотической ДНК обвиты вокруг белковых частиц, образуя нуклеосомы, поэтому репликация этой ДНК,
- 20. Отличия репликации эукариот Репликация концов (теломер) линейной эукариотической хромосомы создает определенные трудности, которые решаются с помощью
- 21. - отсчёта клеточных делений и старения, вследствие недорепликации последовательностей ДНК на концах хромосом (теломерных участков). Чтобы
- 22. Теломерная гипотеза старения Максимальное число делений зависит от типа клеток и для большинства человеческих клеток этот
- 23. Лимит (предел) Хейфлика Лимит Хейфлика для различных животных граница количества делений соматических клеток Эта граница была
- 24. © http://www.pereplet.ru/nauka/Soros/pdf/0005_008.pdf Строение и работа теломеразы В результате деятельности теломеразы длина теломерных участков хромосом клетки увеличивается
- 25. В ходе исследования этого фермента (состоящего, из РНК-компонента и белкового компонента) выяснилось, что РНК-компонент экспрессируется на
- 26. В октябре 2009 года в Стокгольме объявлены имена лауреатов Нобелевской премии по физиологии и медицине. Это
- 28. Скачать презентацию

План лекции:
Механизмы репликации ДНК
Репликационная вилка
Современная модель репликации
Ферменты репликации
План лекции:
Механизмы репликации ДНК
Репликационная вилка
Современная модель репликации
Ферменты репликации

Механизмы репликации ДНК
Во время репликации каждая из цепей родительской ДНК служит
Механизмы репликации ДНК
Во время репликации каждая из цепей родительской ДНК служит

Механизмы репликации ДНК
Три альтернативные гипотезы.
1958 г. Подтверждена гипотеза полуконсервативной репликации (эксперимент
Механизмы репликации ДНК
Три альтернативные гипотезы.
1958 г. Подтверждена гипотеза полуконсервативной репликации (эксперимент

Репликационная вилка
1957 г. А.Корнберг обнаружил ДНК-полимеразу I – фермент, катализирующий процесс
Репликационная вилка
1957 г. А.Корнберг обнаружил ДНК-полимеразу I – фермент, катализирующий процесс

Механизмы репликации ДНК у прокариот
Реплицирующуюся ДНК бактерии можно наблюдать в электронном
Механизмы репликации ДНК у прокариот
Реплицирующуюся ДНК бактерии можно наблюдать в электронном
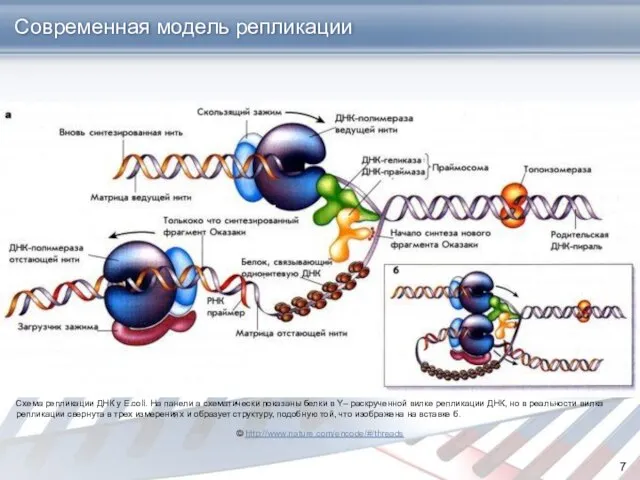
Современная модель репликации
© http://www.nature.com/encode/#/threads
Схема репликации ДНК у E.coli. На панели а схематически
Современная модель репликации
© http://www.nature.com/encode/#/threads
Схема репликации ДНК у E.coli. На панели а схематически

Ферменты репликации
Чтобы репликационная вилка могла продвигаться вперёд, вся хромосома впереди неё
Ферменты репликации
Чтобы репликационная вилка могла продвигаться вперёд, вся хромосома впереди неё
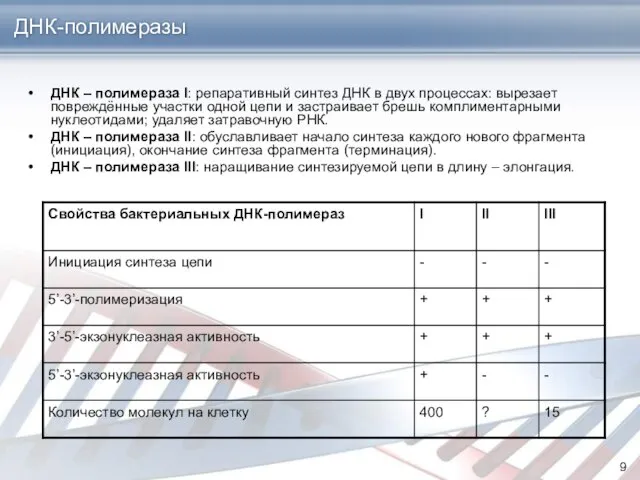
ДНК – полимераза I: репаративный синтез ДНК в двух процессах: вырезает
ДНК – полимераза I: репаративный синтез ДНК в двух процессах: вырезает
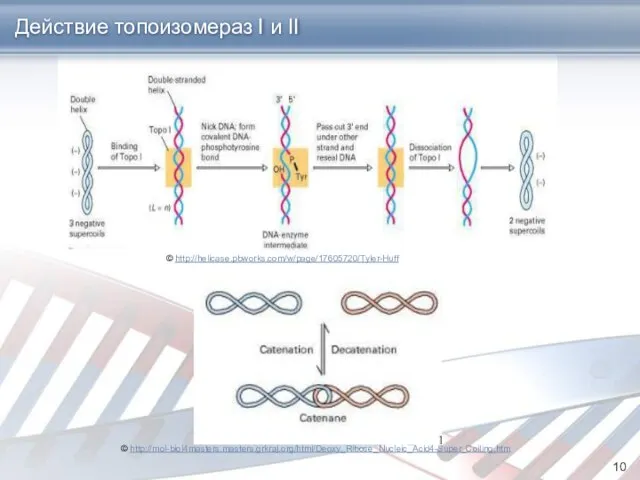
Действие топоизомераз I и II
© http://helicase.pbworks.com/w/page/17605720/Tyler-Huff
© http://mol-biol4masters.masters.grkraj.org/html/Deoxy_Ribose_Nucleic_Acid4-Super_Coiling.htm
Действие топоизомераз I и II
© http://helicase.pbworks.com/w/page/17605720/Tyler-Huff
© http://mol-biol4masters.masters.grkraj.org/html/Deoxy_Ribose_Nucleic_Acid4-Super_Coiling.htm
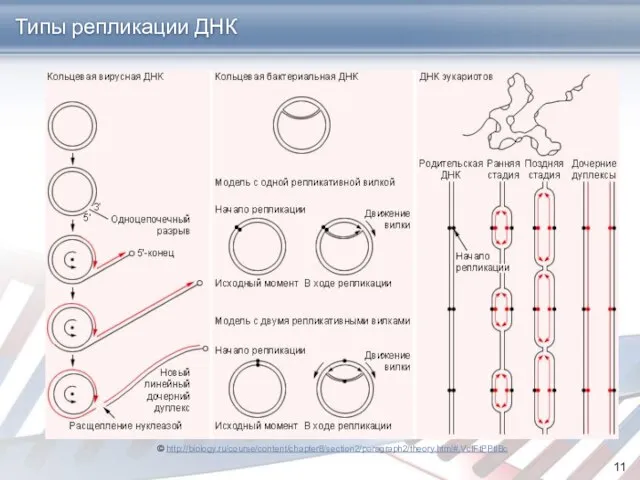
Типы репликации ДНК
© http://biology.ru/course/content/chapter8/section2/paragraph2/theory.html#.VcIFtPPtlBc
Типы репликации ДНК
© http://biology.ru/course/content/chapter8/section2/paragraph2/theory.html#.VcIFtPPtlBc

В кольцевой хромосоме репликационный глазок образует Ө-структуру.
Ө-механизм
В кольцевой хромосоме репликационный глазок образует Ө-структуру.
Ө-механизм
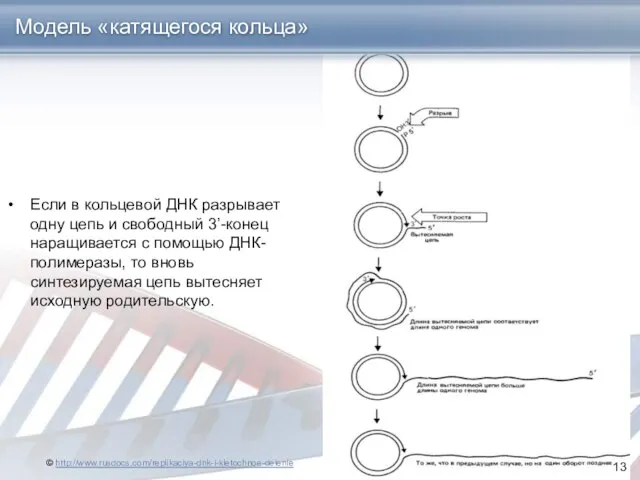
Если в кольцевой ДНК разрывает одну цепь и свободный 3’-конец наращивается
Если в кольцевой ДНК разрывает одну цепь и свободный 3’-конец наращивается
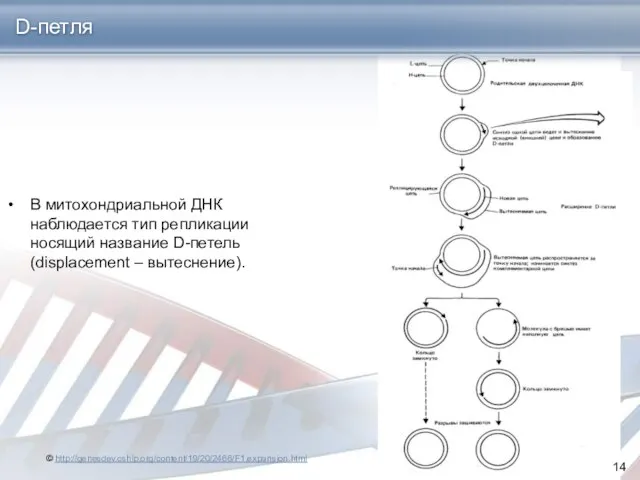
В митохондриальной ДНК наблюдается тип репликации носящий название D-петель (displacement –
В митохондриальной ДНК наблюдается тип репликации носящий название D-петель (displacement –

Специфические мембранные белки обеспечивают контакт ДНК с внутренней поверхностью цитоплазматической мембраны.
Специфические мембранные белки обеспечивают контакт ДНК с внутренней поверхностью цитоплазматической мембраны.

А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране
А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране
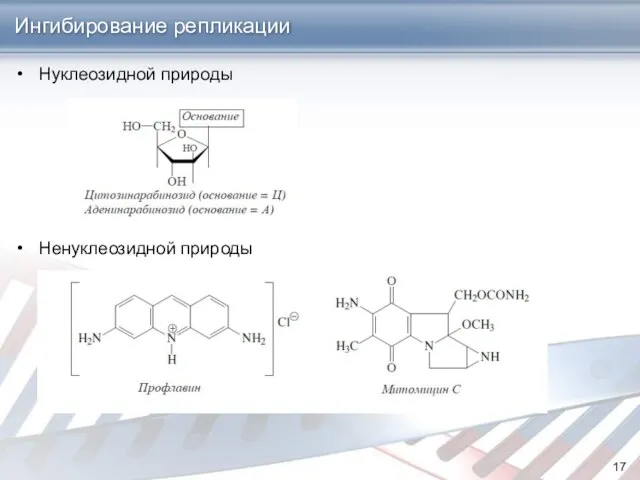
Нуклеозидной природы
Ненуклеозидной природы
Ингибирование репликации
Нуклеозидной природы
Ненуклеозидной природы
Ингибирование репликации
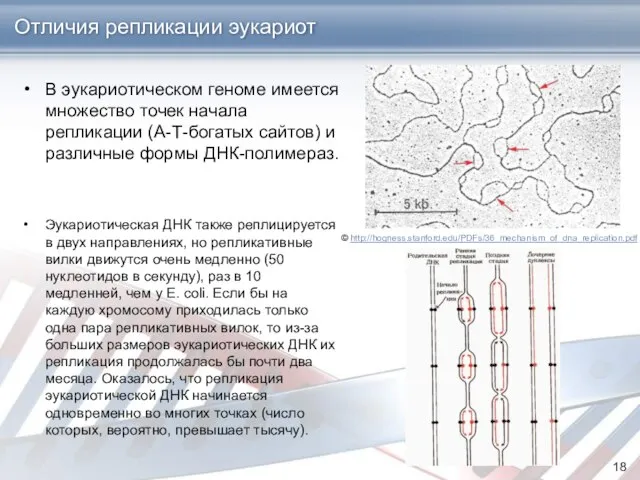
Отличия репликации эукариот
В эукариотическом геноме имеется множество точек начала репликации (А-Т-богатых
Отличия репликации эукариот
В эукариотическом геноме имеется множество точек начала репликации (А-Т-богатых
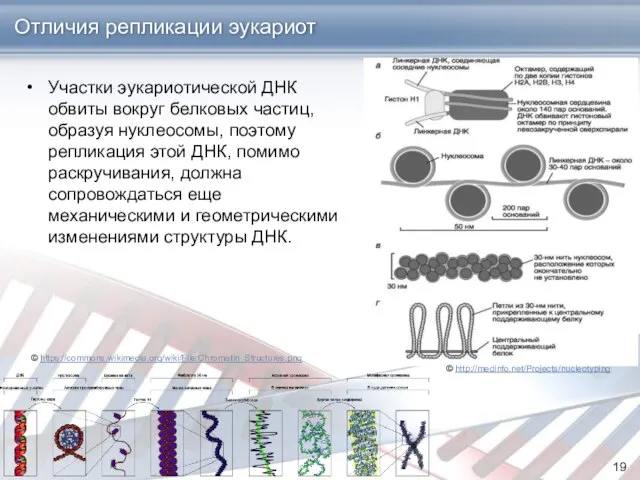
Отличия репликации эукариот
Участки эукариотической ДНК обвиты вокруг белковых частиц, образуя нуклеосомы,
Отличия репликации эукариот
Участки эукариотической ДНК обвиты вокруг белковых частиц, образуя нуклеосомы,
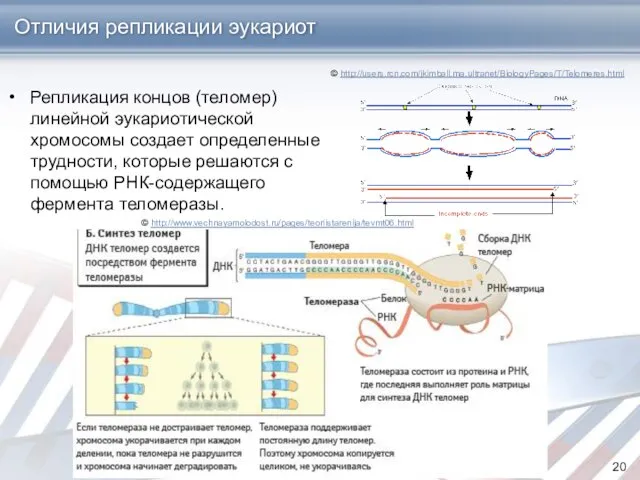
Отличия репликации эукариот
Репликация концов (теломер) линейной эукариотической хромосомы создает определенные трудности,
Отличия репликации эукариот
Репликация концов (теломер) линейной эукариотической хромосомы создает определенные трудности,

- отсчёта клеточных делений и старения, вследствие недорепликации последовательностей ДНК на
- отсчёта клеточных делений и старения, вследствие недорепликации последовательностей ДНК на
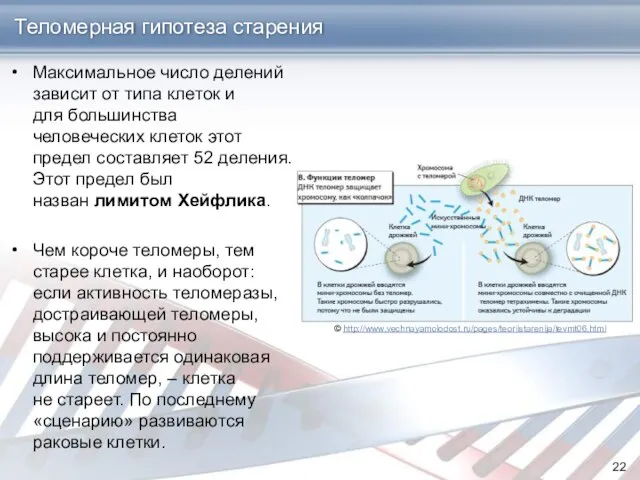
Теломерная гипотеза старения
Максимальное число делений зависит от типа клеток и для большинства человеческих
Теломерная гипотеза старения
Максимальное число делений зависит от типа клеток и для большинства человеческих

Лимит (предел) Хейфлика
Лимит Хейфлика для различных животных
граница количества делений соматических клеток
Лимит (предел) Хейфлика
Лимит Хейфлика для различных животных
граница количества делений соматических клеток
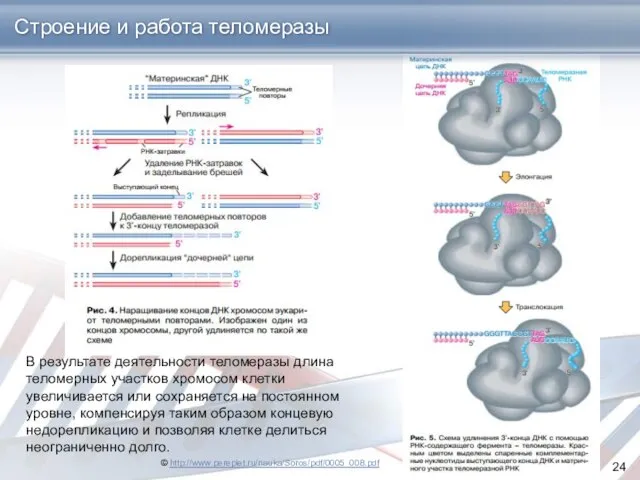
© http://www.pereplet.ru/nauka/Soros/pdf/0005_008.pdf
Строение и работа теломеразы
В результате деятельности теломеразы длина теломерных
© http://www.pereplet.ru/nauka/Soros/pdf/0005_008.pdf
Строение и работа теломеразы
В результате деятельности теломеразы длина теломерных

В ходе исследования этого фермента (состоящего, из РНК-компонента и белкового компонента)
В ходе исследования этого фермента (состоящего, из РНК-компонента и белкового компонента)

В октябре 2009 года в Стокгольме объявлены имена лауреатов Нобелевской премии
В октябре 2009 года в Стокгольме объявлены имена лауреатов Нобелевской премии
 Презентация на тему "Наследственные заболевания" - скачать бесплатно презентации по Биологии
Презентация на тему "Наследственные заболевания" - скачать бесплатно презентации по Биологии Анатомия и физиология репродуктивной системы. Мужские половые органы
Анатомия и физиология репродуктивной системы. Мужские половые органы Антропогенез. Долгопяты: величие и закат. (часть 5)
Антропогенез. Долгопяты: величие и закат. (часть 5) Круги кровообращения
Круги кровообращения Гормоны коркового вещества надпочечников кортикостероиды
Гормоны коркового вещества надпочечников кортикостероиды Ядовитые грибы и ягоды
Ядовитые грибы и ягоды Как избавиться от сорняков и травы навсегда
Как избавиться от сорняков и травы навсегда Презентация на тему "Болото и его обитатели" - скачать презентации по Биологии
Презентация на тему "Болото и его обитатели" - скачать презентации по Биологии Строение и функции органа зрения
Строение и функции органа зрения Презентация на тему "Чарльз Дарвин" - скачать презентации по Биологии
Презентация на тему "Чарльз Дарвин" - скачать презентации по Биологии Презентация на тему "Кровеносная и дыхательная система" - скачать презентации по Биологии
Презентация на тему "Кровеносная и дыхательная система" - скачать презентации по Биологии Тип кольчатые черви Учитель биологии МОУ «СОШ № 20» г. Энгельса Егубова Л.В.
Тип кольчатые черви Учитель биологии МОУ «СОШ № 20» г. Энгельса Егубова Л.В. Развитие жизни на Земле
Развитие жизни на Земле Многообразие млекопитающих
Многообразие млекопитающих Человек – часть живой природы
Человек – часть живой природы ГЕНЕТИКА ЧЕЛОВЕКА Мне необходимо разобраться самому, а чтобы разобраться самому, надо думать сообща. Борис Вас
ГЕНЕТИКА ЧЕЛОВЕКА Мне необходимо разобраться самому, а чтобы разобраться самому, надо думать сообща. Борис Вас Презентация на тему СТРОЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ.
Презентация на тему СТРОЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. Главные части и органоиды клеток
Главные части и органоиды клеток  Анализаторы. 1.Чем анализатор отличается от органа чувств? 2. В чем выражена специфичность анализатора? 3. Что такое иллюзии и отче
Анализаторы. 1.Чем анализатор отличается от органа чувств? 2. В чем выражена специфичность анализатора? 3. Что такое иллюзии и отче Prezentatsia
Prezentatsia Предмет и задачи микробиологии и иммунологии
Предмет и задачи микробиологии и иммунологии Растительный и животный мир Чувашии
Растительный и животный мир Чувашии Презентация на тему "Определение типа темперамента учащихся" - скачать презентации по Биологии
Презентация на тему "Определение типа темперамента учащихся" - скачать презентации по Биологии Презентация на тему "Изучение зависимости количества стрекоз от времени дня и погодных условий" - скачать презентации по Био
Презентация на тему "Изучение зависимости количества стрекоз от времени дня и погодных условий" - скачать презентации по Био Представители редких и исчезающих видов растений Южного Урала
Представители редких и исчезающих видов растений Южного Урала Развитие жизни в Кайнозойскую эру Работу выполнили ученики 11 класса Клыжко В. Никитин Е.
Развитие жизни в Кайнозойскую эру Работу выполнили ученики 11 класса Клыжко В. Никитин Е. Презентация на тему "Дмитрий Иванович Мендилеев" - скачать презентации по Биологии
Презентация на тему "Дмитрий Иванович Мендилеев" - скачать презентации по Биологии Классификация. Систематика
Классификация. Систематика