- Структура и функции нуклеиновых кислот

Содержание
- 2. СТРУКТУРА И ФУНКЦИИ НУКЛЕИНОВЫХ КИСЛОТ МОЛЕКУЛЯРНАЯ БИОТЕХНОЛОГИЯ ч. 2
- 3. СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ Хронология открытий, подготовивших создание Уотсоном и Криком модели двойной спирали ДНК 1868г. Обнаружен
- 4. Нуклеиновые кислоты являются нерегулярными полимерами, мономеры которых - нуклеотиды. Нуклеотид = нуклеозид + фосфорная кислота =
- 5. Нуклеотиды соединяются друг с другом в полимерную цепочку с помощью фосфодиэфирных связей. Азотистые основания не принимают
- 6. Существует два класса азотистых оснований. Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла. Пиримидины:
- 7. Принципы строения ДНК 1. Нерегулярность. Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование
- 8. Принципы строения ДНК Нерегулярность, антипараллельность,комплементарность, наличие регулярной вторичной структуры
- 9. ФУНКЦИИ ДНК 1. ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода. 2. Воспроизведение
- 10. РЕПЛИКАЦИЯ ДНК Процесс, осуществляемый комплексом ферментов и белков, выполняющих топологическую функцию, суть которого в образовании идентичных
- 11. Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи и всегда в направлении 5'
- 12. Синтез одной нити ДНК идет короткими фрагментами, которые сшиваются с помощью ДНК-лигазы. У бактерий и высших
- 13. ФЕРМЕНТЫ РЕПЛИКАЦИИ ДНК-полимеразы Эти ферменты присутствуют во всех прокариотических и эукариотических клетках. Сравнительные характеристики ДНК-полимераз E.
- 14. К репликации имеют отношение полимеразы I и III. Причем именно полимераза III является репликазой, т.е. она
- 15. Геликазы Геликазы - ферменты, денатурирующие ДНК. Для начала “работы” геликазы требуется одноцепочечный участок ДНК, т.е. геликаза
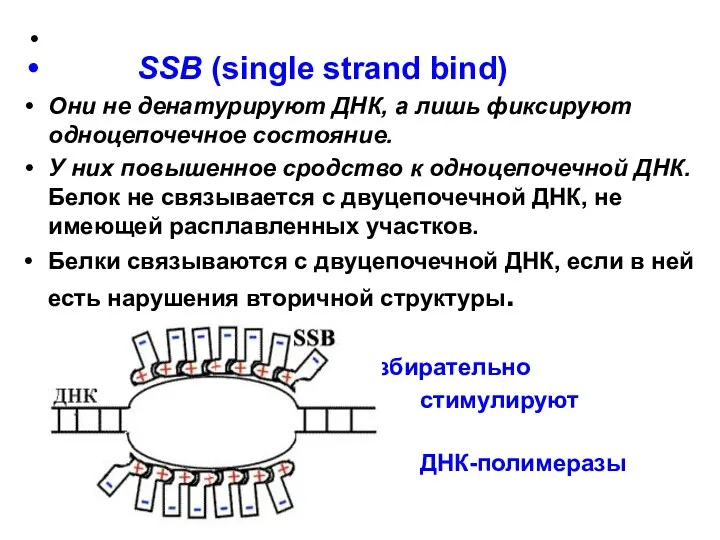
- 16. SSB (single strand bind) Они не денатурируют ДНК, а лишь фиксируют одноцепочечное состояние. У них повышенное
- 17. Топоизомеразы Топоизомеразы - ферменты, изменяющие топологию ДНК, т.е. катализирующие переходы в молекулах ДНК, связанные с изменением
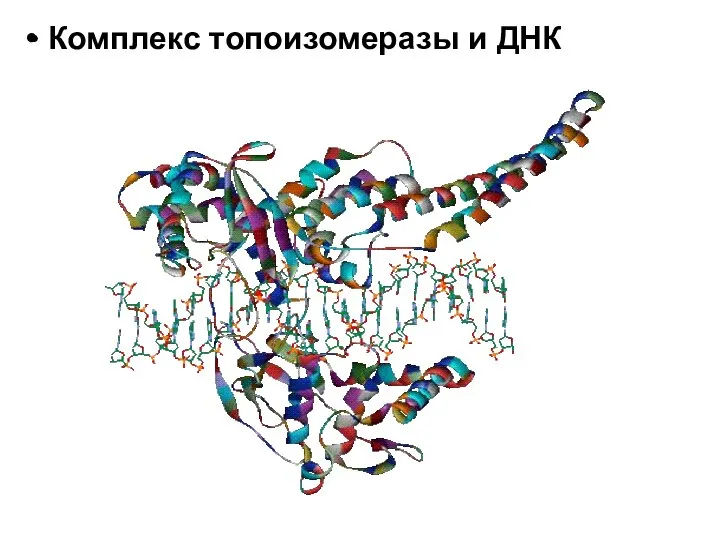
- 18. Комплекс топоизомеразы и ДНК
- 19. Механизм действия топоизомеразы В процессе репликации расплетание цепей ДНК может приводить к образованию особых структур: суперскрученных
- 20. РНК-праймаза Синтезирует РНК-затравки для синтеза дочерних нитей ДНК ДНК - лигазы Необходимы для соединения цепей ДНК
- 21. ИНИЦИАЦИЯ РЕПЛИКАЦИИ У Е.coli Белок-инициатор DnaA играет ключевую роль в инициации репликации хромосомы у многих бактерий.
- 23. Элонгация обеих нитей синтезирующейся ДНК
- 24. Терминация репликации
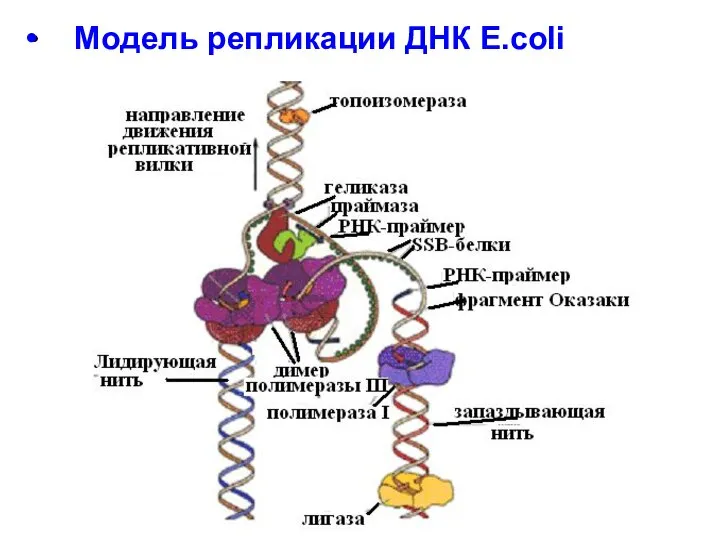
- 25. Модель репликации ДНК E.coli
- 26. Особенности репликации ДНК эукариот Каждая эукариотическая хромосома - полирепликон.
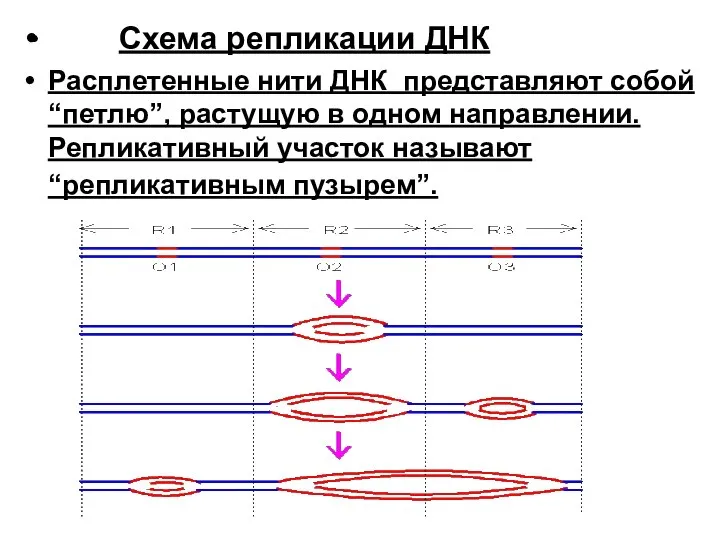
- 28. Cхема репликации ДНК Расплетенные нити ДНК представляют собой “петлю”, растущую в одном направлении. Репликативный участок называют
- 29. Репликация концов ДНК хромосом эукариот 3'-оверхенги, т.е. выступающие 3'-концы материнских цепей узнаются теломеразой - ферментом, содержащим
- 31. Белки и ферменты репликации
- 33. Скачать презентацию

СТРУКТУРА И ФУНКЦИИ
НУКЛЕИНОВЫХ КИСЛОТ
МОЛЕКУЛЯРНАЯ БИОТЕХНОЛОГИЯ ч. 2
СТРУКТУРА И ФУНКЦИИ
НУКЛЕИНОВЫХ КИСЛОТ
МОЛЕКУЛЯРНАЯ БИОТЕХНОЛОГИЯ ч. 2

СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
Хронология открытий, подготовивших создание Уотсоном и Криком
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
Хронология открытий, подготовивших создание Уотсоном и Криком

Нуклеиновые кислоты являются нерегулярными полимерами, мономеры которых - нуклеотиды.
Нуклеотид = нуклеозид
Нуклеиновые кислоты являются нерегулярными полимерами, мономеры которых - нуклеотиды.
Нуклеотид = нуклеозид
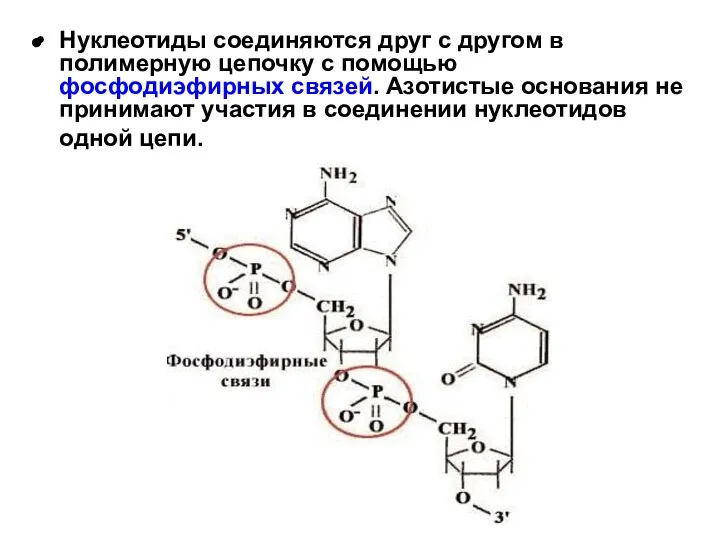
Нуклеотиды соединяются друг с другом в полимерную цепочку с помощью фосфодиэфирных
Нуклеотиды соединяются друг с другом в полимерную цепочку с помощью фосфодиэфирных
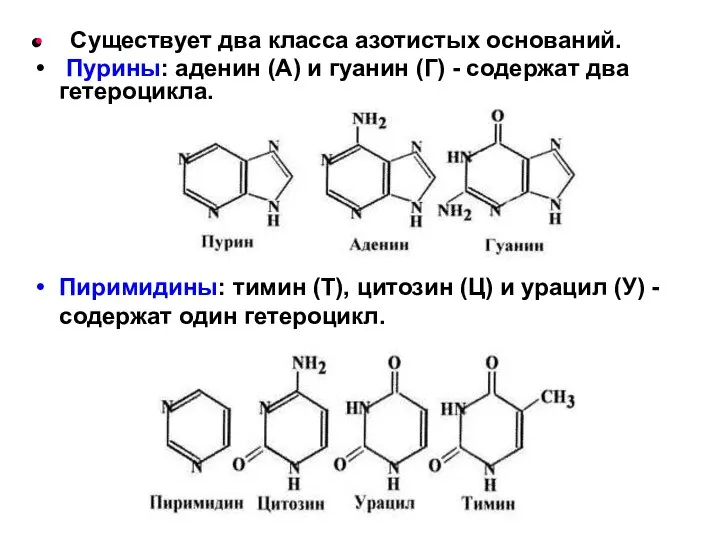
Существует два класса азотистых оснований.
Пурины: аденин (А) и гуанин
Существует два класса азотистых оснований.
Пурины: аденин (А) и гуанин

Принципы строения ДНК
1. Нерегулярность.
Существует регулярный сахарофосфатный остов, к которому присоединены
Принципы строения ДНК
1. Нерегулярность.
Существует регулярный сахарофосфатный остов, к которому присоединены
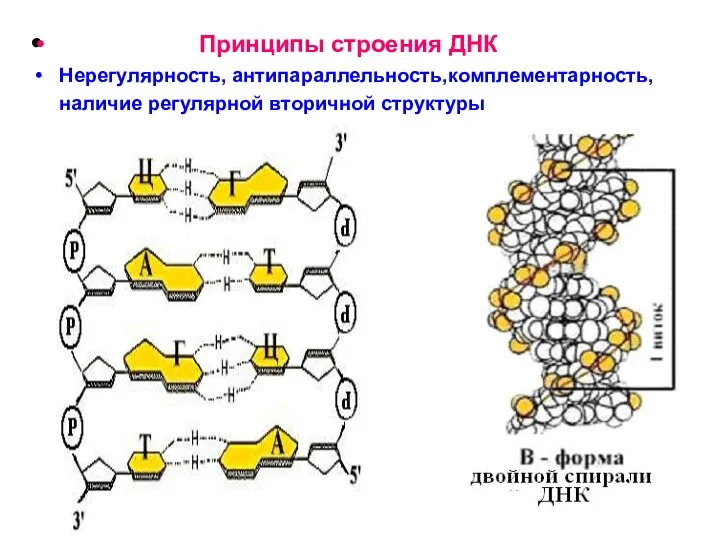
Принципы строения ДНК
Нерегулярность, антипараллельность,комплементарность, наличие регулярной вторичной структуры
Принципы строения ДНК
Нерегулярность, антипараллельность,комплементарность, наличие регулярной вторичной структуры

ФУНКЦИИ ДНК
1. ДНК является носителем генетической информации.
Функция обеспечивается фактом
ФУНКЦИИ ДНК
1. ДНК является носителем генетической информации.
Функция обеспечивается фактом

РЕПЛИКАЦИЯ ДНК
Процесс, осуществляемый комплексом ферментов и белков,
РЕПЛИКАЦИЯ ДНК
Процесс, осуществляемый комплексом ферментов и белков,

Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи
Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи
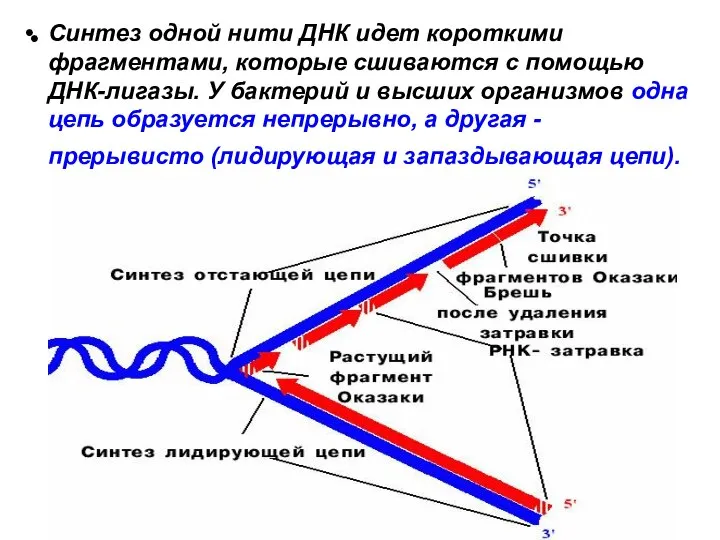
Синтез одной нити ДНК идет короткими фрагментами, которые сшиваются с помощью
Синтез одной нити ДНК идет короткими фрагментами, которые сшиваются с помощью

ФЕРМЕНТЫ РЕПЛИКАЦИИ
ДНК-полимеразы
Эти ферменты присутствуют во всех прокариотических и эукариотических клетках.
Сравнительные характеристики
ФЕРМЕНТЫ РЕПЛИКАЦИИ
ДНК-полимеразы
Эти ферменты присутствуют во всех прокариотических и эукариотических клетках.
Сравнительные характеристики

К репликации имеют отношение полимеразы I и III.
Причем именно полимераза
К репликации имеют отношение полимеразы I и III.
Причем именно полимераза

Геликазы
Геликазы - ферменты, денатурирующие ДНК.
Для начала “работы” геликазы требуется
Геликазы
Геликазы - ферменты, денатурирующие ДНК.
Для начала “работы” геликазы требуется
SSB (single strand bind)
Они не денатурируют ДНК, а лишь фиксируют
SSB (single strand bind)
Они не денатурируют ДНК, а лишь фиксируют

Топоизомеразы
Топоизомеразы - ферменты, изменяющие топологию ДНК, т.е. катализирующие переходы в
Топоизомеразы
Топоизомеразы - ферменты, изменяющие топологию ДНК, т.е. катализирующие переходы в
Комплекс топоизомеразы и ДНК
Комплекс топоизомеразы и ДНК

Механизм действия топоизомеразы
В процессе репликации расплетание цепей ДНК может приводить к
Механизм действия топоизомеразы
В процессе репликации расплетание цепей ДНК может приводить к

РНК-праймаза
Синтезирует РНК-затравки для синтеза дочерних нитей ДНК
ДНК - лигазы
Необходимы
РНК-праймаза
Синтезирует РНК-затравки для синтеза дочерних нитей ДНК
ДНК - лигазы
Необходимы

ИНИЦИАЦИЯ РЕПЛИКАЦИИ У Е.coli
Белок-инициатор DnaA играет ключевую роль в инициации репликации
ИНИЦИАЦИЯ РЕПЛИКАЦИИ У Е.coli
Белок-инициатор DnaA играет ключевую роль в инициации репликации


Элонгация обеих нитей синтезирующейся ДНК
Элонгация обеих нитей синтезирующейся ДНК

Терминация репликации
Терминация репликации
Модель репликации ДНК E.coli
Модель репликации ДНК E.coli

Особенности репликации ДНК эукариот
Каждая эукариотическая хромосома - полирепликон.
Особенности репликации ДНК эукариот
Каждая эукариотическая хромосома - полирепликон.

Cхема репликации ДНК
Расплетенные нити ДНК представляют собой “петлю”, растущую в
Cхема репликации ДНК
Расплетенные нити ДНК представляют собой “петлю”, растущую в
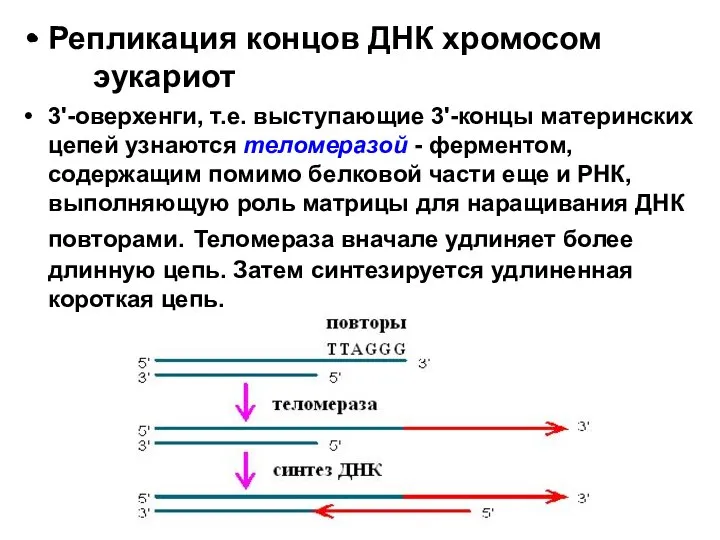
Репликация концов ДНК хромосом эукариот
3'-оверхенги, т.е. выступающие 3'-концы материнских цепей
Репликация концов ДНК хромосом эукариот
3'-оверхенги, т.е. выступающие 3'-концы материнских цепей


Белки и ферменты репликации
Белки и ферменты репликации
 Очень медленно идет, Дом она с собой несет. Испугается чего-то, В панцирь с головой уйдет
Очень медленно идет, Дом она с собой несет. Испугается чего-то, В панцирь с головой уйдет ВОЗНИКНОВЕНИЕ ЖИЗНИ
ВОЗНИКНОВЕНИЕ ЖИЗНИ  Физиология пищеварения
Физиология пищеварения Изоляция – эволюционный фактор
Изоляция – эволюционный фактор Основные понятия генетики. Моногибридное скрещивание
Основные понятия генетики. Моногибридное скрещивание Следы жизнедеятельности животных
Следы жизнедеятельности животных Решение задач по генетике Памятка для учащихся 9 – 10 классов Составила: Белова Римма Васильевна, учитель биологии ГБОУ СОШ с. Чёр
Решение задач по генетике Памятка для учащихся 9 – 10 классов Составила: Белова Римма Васильевна, учитель биологии ГБОУ СОШ с. Чёр Ихтиофауна памятника природы Долина реки Издревая
Ихтиофауна памятника природы Долина реки Издревая Тип Кишечнополостные
Тип Кишечнополостные Растения и животные луга УЧИТЕЛЬ Макарова Светлана Павловна средняя школа № 65 г.Кирова
Растения и животные луга УЧИТЕЛЬ Макарова Светлана Павловна средняя школа № 65 г.Кирова Биоразнообразие природы Московской области
Биоразнообразие природы Московской области Galápagos tortoise
Galápagos tortoise Мыши - семейство млекопитающих отряда грызунов
Мыши - семейство млекопитающих отряда грызунов Сельское хозяйство. Растениеводство.
Сельское хозяйство. Растениеводство. Пингвины
Пингвины Ұлулар
Ұлулар Кости и органы. Самостоятельная работа. Вариант 2
Кости и органы. Самостоятельная работа. Вариант 2 Презентация на тему "Дикие - дикие кошки" - скачать презентации по Биологии
Презентация на тему "Дикие - дикие кошки" - скачать презентации по Биологии Конечный мозг. Большие полушария. Кора
Конечный мозг. Большие полушария. Кора Презентация на тему "БЕЛКИ, КАК БИОПОЛИМЕРЫ, ИХ СОСТАВ" - скачать бесплатно презентации по Биологии
Презентация на тему "БЕЛКИ, КАК БИОПОЛИМЕРЫ, ИХ СОСТАВ" - скачать бесплатно презентации по Биологии Способы размножения растений
Способы размножения растений Ярмарка знаний. Цветок, плод, семя. 6 класс
Ярмарка знаний. Цветок, плод, семя. 6 класс Химические реакции с участием моносахаридов. Структура и функции ди-, олиго- и полисахаридов
Химические реакции с участием моносахаридов. Структура и функции ди-, олиго- и полисахаридов Предмет физиологии центральной нервной системы
Предмет физиологии центральной нервной системы Обитатели воды
Обитатели воды Чайный гриб
Чайный гриб Ткани растений
Ткани растений Завдання для екзамену з дисципліни “Ветеринарна мікробіологія”
Завдання для екзамену з дисципліни “Ветеринарна мікробіологія”