- Терминация репликации. Теломеры. Их репликация и роль в функционировании эукариотической клетки

Содержание
- 2. Primary non-transformed cell cultures tend to change with every passage. Population doubling time increases, cell morphology
- 3. Cells in Culture (5, 10, 15 and 40 p)
- 4. Терминация репликации у Е.coli Последовательности, которые обеспечивают терминацию, у Е.coli называются ter-сайтами. Они содержат короткую (около
- 5. Движение репликативных вилок навстречу друг другу сопровождается гомологичной рекомбинацией между дочерними хроматидами. В том случае, если
- 6. Окончание репликации ter (terminus) находятся на противоположной стороне от ориджина кольцевой хромосомы E. coli. Ter сайты
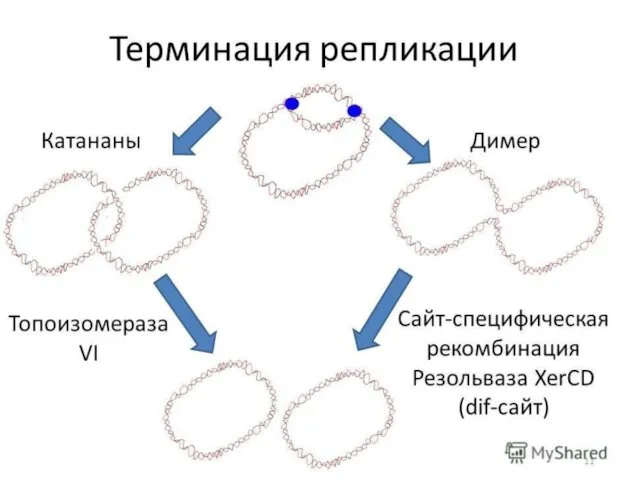
- 7. Встреча двух репликативных вилок в конце цикла репликации бактериальной хромосомы сопровождается несколькими событиями, которые необходимы для
- 10. Герман Джозеф Мёллер ( Hermann Joseph "H. J." Muller; декабрь 1890, Нью-Йорк — апрель 1967, Индианаполис)
- 11. Ба́рбара Мак-Кли́нток ( Barbara McClintock; июнь 1902, Коннектикут сентябрь 1992, Нью-Йорк) Лауреат Нобелевской премии по физиологии
- 12. История расшифровки механизма естественного старения Вплоть до конца прошлого века механизмы старения оставались совершенно непонятными. Несмотря
- 13. Леонард Хейфлик (Leonard Hayflick 20 мая1928, Филадельфия Медаль Джона Скотта (1914)
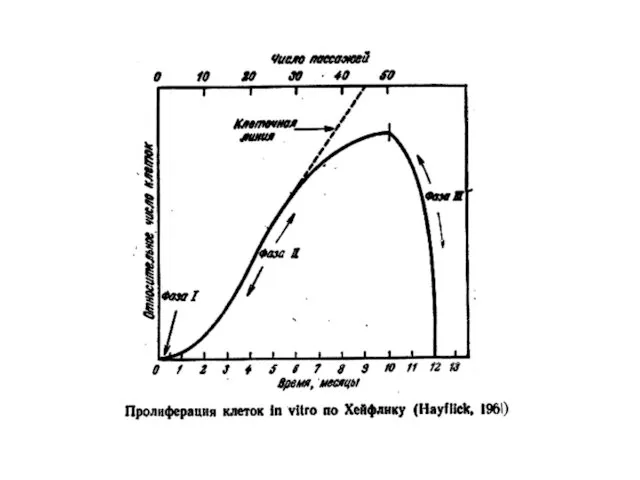
- 14. Лимит Хейфлика Хейфлик культивировал клетки легкого эмбриона человека и обнаружил, что нормальные диплоидные клетки в культуре
- 16. В результате работ Хейфлика стало ясно, что причина ограниченной способности клеток к пролиферации лежит в самих
- 17. А.М. Оловников
- 18. Недорепликация теломер Прямоугольниками на отстающей нити изображены РНК-праймеры Короткие стрелки – фрагменты Оказаки
- 19. Теория Оловникова А. М. Оловников в 1971 г. предложил формулу для расчета продолжительности жизни любого клона
- 20. Теломерная теория старения Старение происходит на клеточном уровне: при каждой репликации укорачиваются теломеры хромосом, т.е. дочерняя
- 21. Теория недорепликации Процесс удвоения отстающей цепи ДНК начинается с синтеза коротких РНК-праймеров, или затравок, с З'-концов
- 22. Нобелевская премия 2009 г. в области физиологии и медицины Ученые из США Элайзабет Блекборн, Керол Грейдер
- 24. Фредерик Сенгер Джожеф Галл Frederick Sanger Joseph Gall август 1918 – ноябрь 2013 апрель 1928 две
- 25. Рабочий геном макронуклеуса получается из неактивного микронуклеуса, причем этому процессу сопутствует почти полное уничтожение ДНК микронуклеуса
- 26. Схема геномных перестроек, происходящих в процессе развития макронуклеуса из микронуклеуса у Oxytricha. Часть последовательностей (IES, internally
- 27. Структура теломерного повтора инфузории окситрихи У окситрихи повторяющейся единицей является октамер 5'-ССССАААА-3'/ З'-GGGGТТТТ-З’, расположенный на концах
- 28. Теломерные повторы
- 29. Схема организации теломерного конца хромосомы у окситрихи 1 – повтор из гуаниновых нуклеотидов, 2 – теломерный
- 30. Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с молекулами цитозина, как это бывает
- 31. Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с молекулами цитозина, как это бывает
- 33. Теломеры дрозофилы У дрозофилы на концах хромосом расположены теломероспецифичные ретротранспозоны, НеТ-А и ТАRТ. Они имеют похожее
- 34. НЕРЕГУЛЯРНЫЕ ТЕЛОМЕРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
- 35. Строение теломер Теломеры человека содержат от 4 до 15 тысяч пар оснований и состоят из повторяющихся
- 36. Динамика укорочения теломер в первичной культуре фибробластов, растущих in vitro, каждое удвоение количества клеток сопровождается уменьшением
- 37. Гипотеза изменения взаимодействий ДНК и ядерного матрикса при укорачивании теломер При укорачивании теломер происходит сдвиг ДНК
- 38. Предсказание теломеразы А.М.Оловниковым Термин «теломераза» ввела Э.Блекборн в 1987 г., однако необходимость существования такого фермента предсказал
- 39. Работа теломеразы(1) Перед началом цикла репликации ДНК теломераза добавляет несколько копий теломерных повторов на З'-конец ДНК.
- 40. Удлинение теломер теломеразой у тетрахимены Теломераза обладает своей молекулой РНК, имеющей матричный участок, с помощью которого
- 41. Теломераза Теломераза - крупный фермент, состоящий из двух основных субъединиц, кодируемых разными генами: 1. TR (hTR)
- 42. Роль теломеразной РНК 1. Изменение специфических оснований у Тетрахимены или дрожжей путем внесения мутации в матрицу
- 43. hnRNP hnRNPs – heterogeneous nuclear ribonucleoproteins – обширное семейство связывющихся с нуклеиновыми кислотами белков часто обнаруживаемые
- 44. Взаимодействие теломер с теломеразой
- 45. Теломеры. Строение Gomez DEGomez DE, Armando RGGomez DE, Armando RG, Farina HGGomez DE, Armando RG, Farina
- 46. Теломеры человека и дрожжей
- 47. Теломеры в процессе старения
- 49. Сигнал от открытой теломеры
- 50. Объединение концов «незащищенных» хромосом
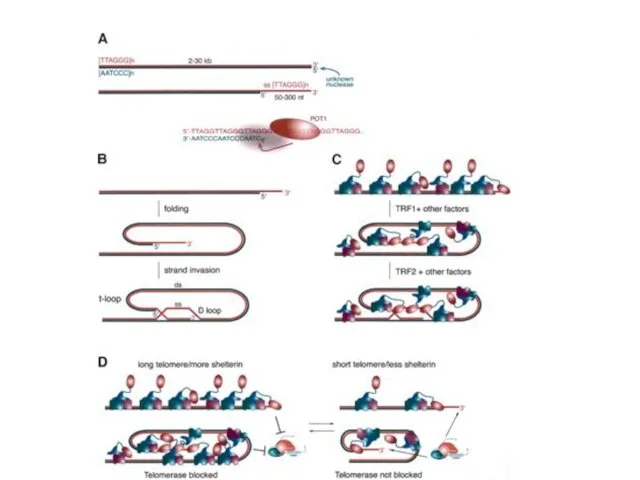
- 51. Теломеры человека и дрожжей
- 52. Белки, связанные с теломерами 1. Поддерживающие пространственную структуру теломер - семейство TRF и другие белки с
- 53. Теломерные белки у человека
- 54. Белки, связанные с теломерами в клетках человека ТRF1 - Теlоmеге repeat binding factor 1 (Pin2). Негативный
- 55. Белки TRF1 и TRF2
- 56. TRF 1 Резкое увеличение концентрации ТRF1 при его экспрессии в культуре клеток под вирусным промотором приводит
- 57. TRF 1 Первое сообщение о белке TRF 1 (Теlomeric repeat binding factor 1) было опубликовано в
- 58. ТRF2 Белок ТRF2 был найден в конце 90-х годов. По структуре он сходен с ТRF1, но
- 59. ТRF2 Пороговая для сенильного торможения пролиферации длина теломер у контрольных клеток составляет 6—7 тыс. пар нуклеотидов.
- 60. TNKS и TNKL У человека и позвоночных животных имеется два изозима танкиразы, (142 и 127 кДа
- 61. TRF 1 ТRF1может быть поли-АDР-рибозилирован, что сопровождается его диссоциацией от ДНК. Эту посттрансляционную модификацию катализирует танкираза
- 62. Белки, взаимодействующие с TRF1 Теломерный белок TINF2 или ТIN2 (ТRF1 intracting nuclear factor 2) - 40кДа,
- 63. Теломерная теория старения объясняет многие давно известные, но без нее непонятные факты 1. Почему злокачественные клетки
- 64. Трудности теломерной теории 1. Теломеры в клетках мыши ~ в 10 раз длиннее, чем в клетках
- 65. Возможное объяснение трудностей теломерной теории старения Для старения и смерти от старости вовсе не обязательно, чтобы
- 66. Репарационные белки, связанные с теломерами Кu (состоит из субъединиц Кu 70 и Ки 80(86) - аутоантиген,
- 67. Влияние АТМ
- 68. Роль белка ретинобластомы в клеточном цикле
- 69. Репликация теломер, зависимая от гомологическоцй рекомбинации при ALT Возможны четыре различных механизма, в зависимости от вовлеченных
- 70. Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных PML-тельцах RAD52 – Связывается с
- 71. Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных PML-тельцах (2) MRE11/RAD50/NBS1 (MRN-комплекс) –
- 72. Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных PML-тельцах (3) WRN и BLM
- 73. Белки, связанные с теломерами 1. Поддерживающие пространственную структуру теломер - семейство TRF и другие белки с
- 74. Гомологи белков человека, ассоциированных с теломерами
- 75. Гомологи белков человека, ассоциированных с теломерами (2)
- 76. Гомологи белков человека, ассоциированных с теломерами (3) при отсутствии функциональных характеристик вместо тривиального названия приводится лишь
- 77. SIRT-белки (сиртуины) Известно, что в клетках каждой ткани функционируют только те гены, которые экспрессируют белки, нужные
- 80. Недорепарация теломер теломерная ДНК повреждается в среднем сильней, чем ДНК из внутренних областей хромосом. имеются сведения
- 81. Причины недорепарации 1. Репарация концов ДНК может встречать те же трудности, что и репликация. 2. Теломеры
- 82. Репарация теломер
- 85. Нарушение функции теломер
- 87. Скачать презентацию

Primary non-transformed cell cultures tend to change with every passage. Population
Primary non-transformed cell cultures tend to change with every passage. Population
Cells in Culture (5, 10, 15 and 40 p)
Cells in Culture (5, 10, 15 and 40 p)
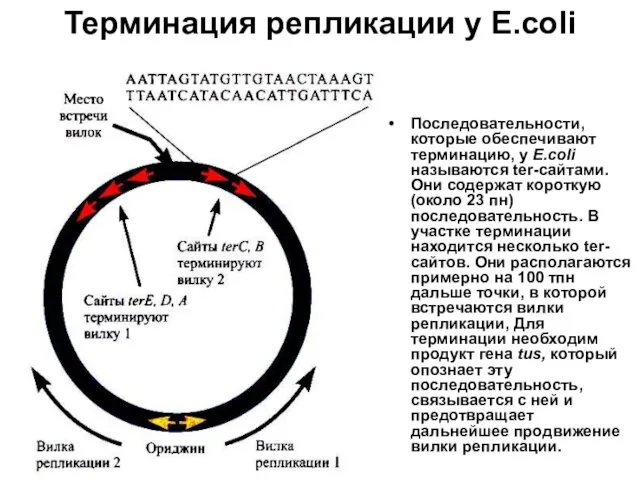
Терминация репликации у Е.coli
Последовательности, которые обеспечивают терминацию, у Е.coli называются ter-сайтами.
Терминация репликации у Е.coli
Последовательности, которые обеспечивают терминацию, у Е.coli называются ter-сайтами.
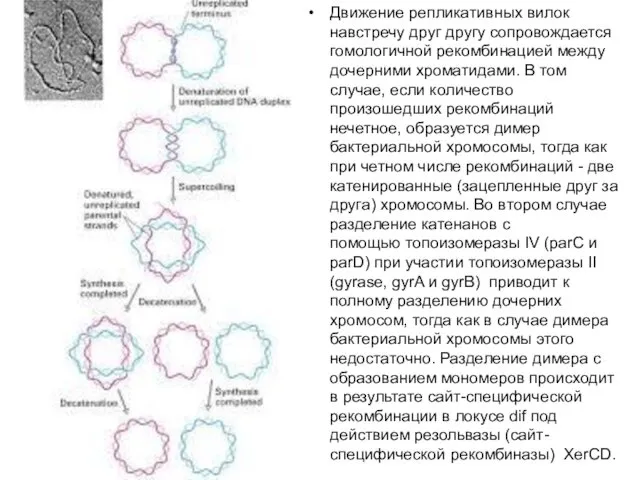
Движение репликативных вилок навстречу друг другу сопровождается гомологичной рекомбинацией между дочерними
Движение репликативных вилок навстречу друг другу сопровождается гомологичной рекомбинацией между дочерними
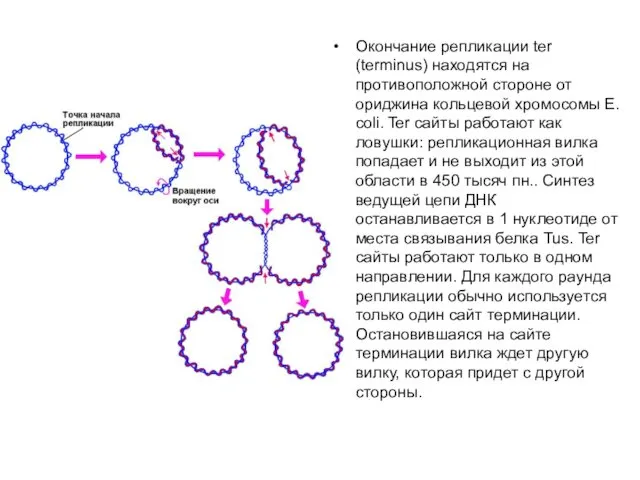
Окончание репликации ter (terminus) находятся на противоположной стороне от ориджина кольцевой
Окончание репликации ter (terminus) находятся на противоположной стороне от ориджина кольцевой
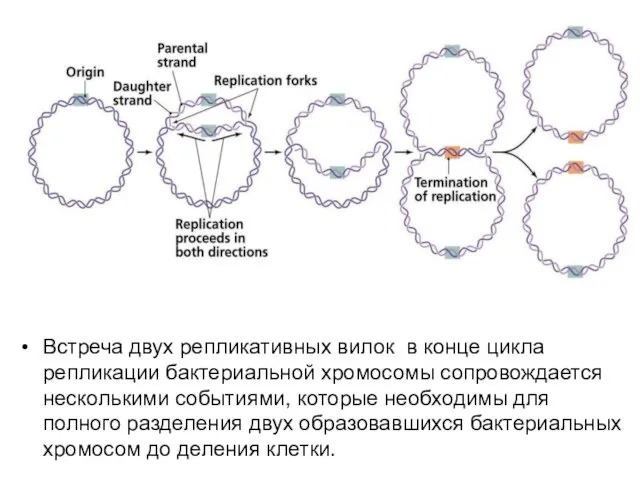
Встреча двух репликативных вилок в конце цикла репликации бактериальной хромосомы сопровождается несколькими
Встреча двух репликативных вилок в конце цикла репликации бактериальной хромосомы сопровождается несколькими

Герман Джозеф Мёллер ( Hermann Joseph "H. J." Muller; декабрь 1890, Нью-Йорк — апрель 1967, Индианаполис)
Американский генетик,
Герман Джозеф Мёллер ( Hermann Joseph "H. J." Muller; декабрь 1890, Нью-Йорк — апрель 1967, Индианаполис)
Американский генетик,

Ба́рбара Мак-Кли́нток ( Barbara McClintock; июнь 1902, Коннектикут сентябрь 1992, Нью-Йорк)
Лауреат Нобелевской премии по физиологии и
Ба́рбара Мак-Кли́нток ( Barbara McClintock; июнь 1902, Коннектикут сентябрь 1992, Нью-Йорк)
Лауреат Нобелевской премии по физиологии и

История расшифровки механизма естественного старения
Вплоть до конца прошлого века механизмы старения
История расшифровки механизма естественного старения
Вплоть до конца прошлого века механизмы старения

Леонард Хейфлик (Leonard Hayflick 20 мая1928, Филадельфия
Медаль Джона Скотта (1914)
Леонард Хейфлик (Leonard Hayflick 20 мая1928, Филадельфия
Медаль Джона Скотта (1914)

Лимит Хейфлика
Хейфлик культивировал клетки легкого эмбриона человека и обнаружил, что нормальные
Лимит Хейфлика
Хейфлик культивировал клетки легкого эмбриона человека и обнаружил, что нормальные

В результате работ Хейфлика стало ясно, что причина ограниченной способности клеток
В результате работ Хейфлика стало ясно, что причина ограниченной способности клеток

А.М. Оловников
А.М. Оловников
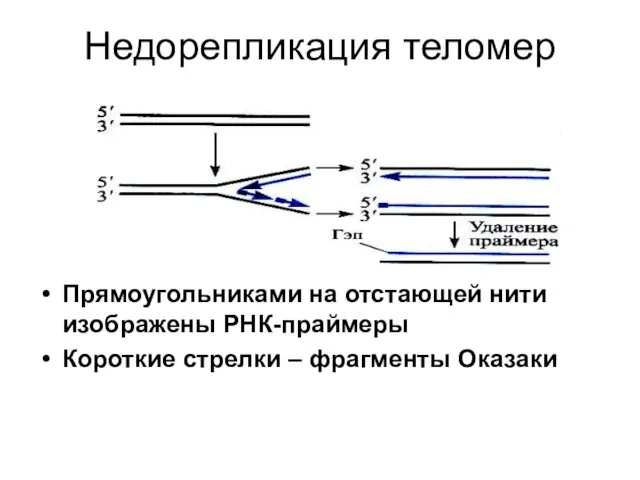
Недорепликация теломер
Прямоугольниками на отстающей нити изображены РНК-праймеры
Короткие стрелки – фрагменты Оказаки
Недорепликация теломер
Прямоугольниками на отстающей нити изображены РНК-праймеры
Короткие стрелки – фрагменты Оказаки

Теория Оловникова
А. М. Оловников в 1971 г. предложил формулу для
Теория Оловникова
А. М. Оловников в 1971 г. предложил формулу для

Теломерная теория старения
Старение происходит на клеточном уровне: при каждой репликации укорачиваются
Теломерная теория старения
Старение происходит на клеточном уровне: при каждой репликации укорачиваются

Теория недорепликации
Процесс удвоения отстающей цепи ДНК начинается с синтеза коротких РНК-праймеров,
Теория недорепликации
Процесс удвоения отстающей цепи ДНК начинается с синтеза коротких РНК-праймеров,

Нобелевская премия 2009 г.
в области физиологии и медицины
Ученые из США Элайзабет
Нобелевская премия 2009 г.
в области физиологии и медицины
Ученые из США Элайзабет


Фредерик Сенгер Джожеф Галл Frederick Sanger Joseph Gall
август 1918 –
Фредерик Сенгер Джожеф Галл Frederick Sanger Joseph Gall август 1918 –
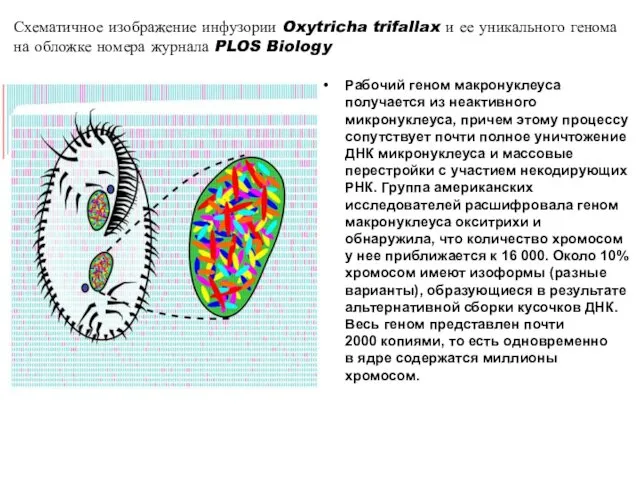
Рабочий геном макронуклеуса получается из неактивного микронуклеуса, причем этому процессу сопутствует
Рабочий геном макронуклеуса получается из неактивного микронуклеуса, причем этому процессу сопутствует
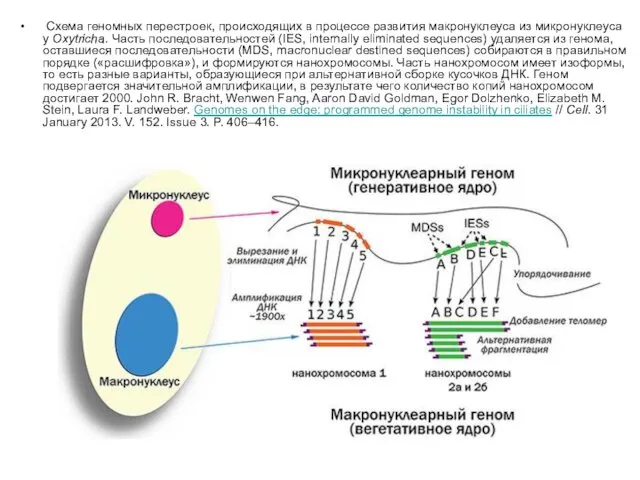
Схема геномных перестроек, происходящих в процессе развития макронуклеуса из микронуклеуса у Oxytricha. Часть
Схема геномных перестроек, происходящих в процессе развития макронуклеуса из микронуклеуса у Oxytricha. Часть
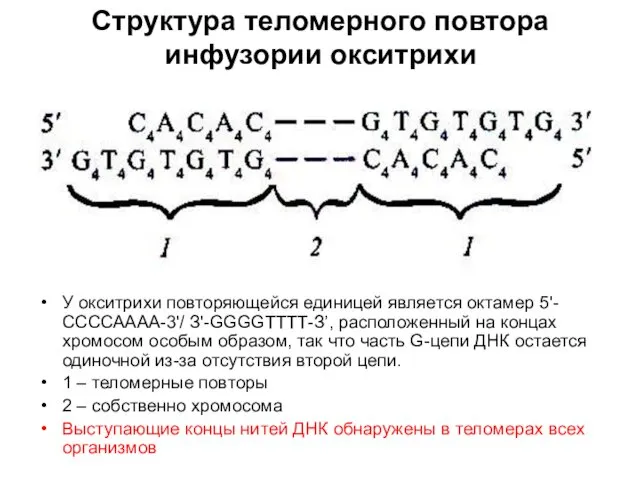
Структура теломерного повтора инфузории окситрихи
У окситрихи повторяющейся единицей является октамер 5'-ССССАААА-3'/
Структура теломерного повтора инфузории окситрихи
У окситрихи повторяющейся единицей является октамер 5'-ССССАААА-3'/
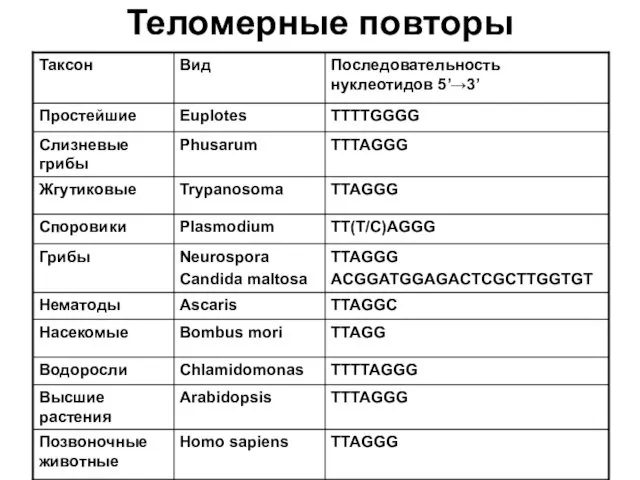
Теломерные повторы
Теломерные повторы

Схема организации теломерного конца хромосомы
у окситрихи
1 – повтор из гуаниновых нуклеотидов,
Схема организации теломерного конца хромосомы
у окситрихи
1 – повтор из гуаниновых нуклеотидов,
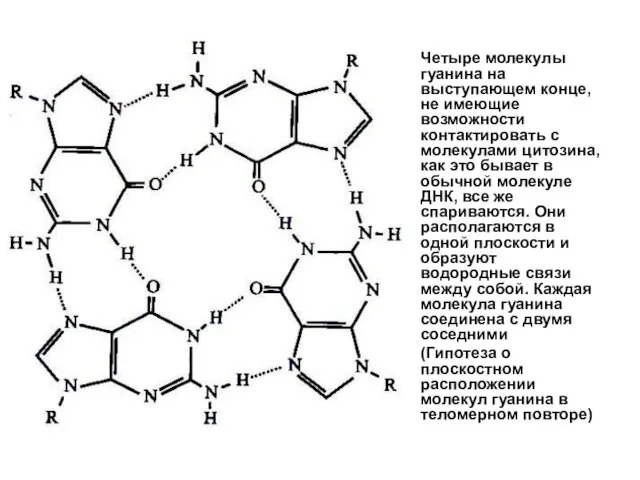
Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с
Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с
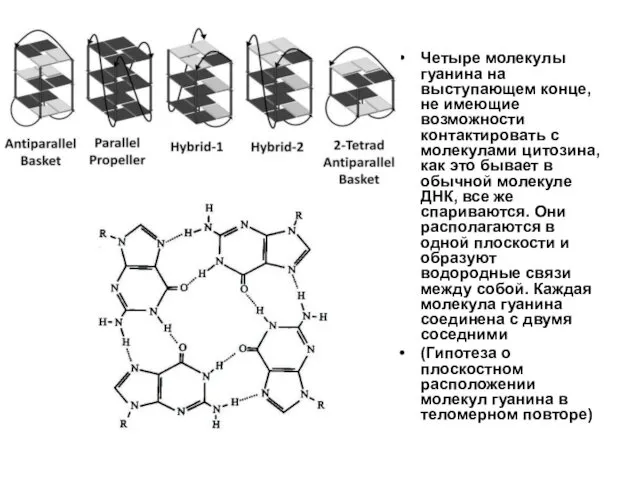
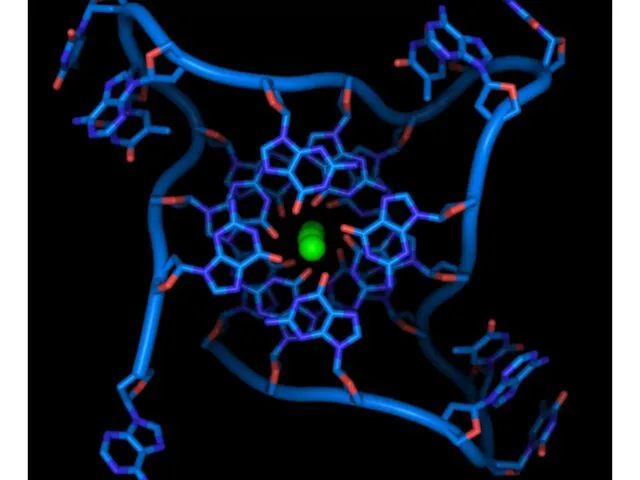
Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с
Четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с

Теломеры дрозофилы
У дрозофилы на концах хромосом расположены теломероспецифичные ретротранспозоны, НеТ-А и
Теломеры дрозофилы
У дрозофилы на концах хромосом расположены теломероспецифичные ретротранспозоны, НеТ-А и
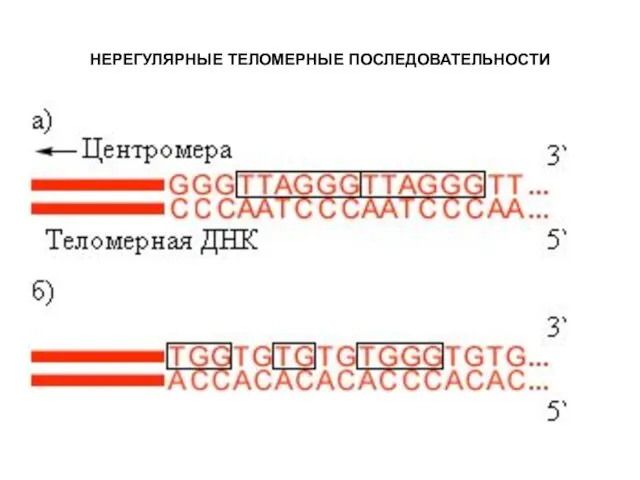
НЕРЕГУЛЯРНЫЕ ТЕЛОМЕРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
НЕРЕГУЛЯРНЫЕ ТЕЛОМЕРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ

Строение теломер
Теломеры человека содержат от 4 до 15 тысяч пар оснований
Строение теломер
Теломеры человека содержат от 4 до 15 тысяч пар оснований

Динамика укорочения теломер
в первичной культуре фибробластов, растущих in vitro, каждое удвоение
Динамика укорочения теломер
в первичной культуре фибробластов, растущих in vitro, каждое удвоение
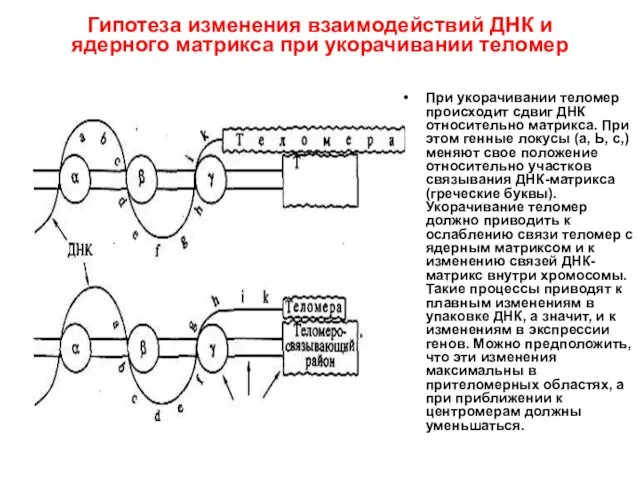
Гипотеза изменения взаимодействий ДНК и ядерного матрикса при укорачивании теломер
При
Гипотеза изменения взаимодействий ДНК и ядерного матрикса при укорачивании теломер
При

Предсказание теломеразы А.М.Оловниковым
Термин «теломераза» ввела Э.Блекборн в 1987 г., однако необходимость
Предсказание теломеразы А.М.Оловниковым
Термин «теломераза» ввела Э.Блекборн в 1987 г., однако необходимость
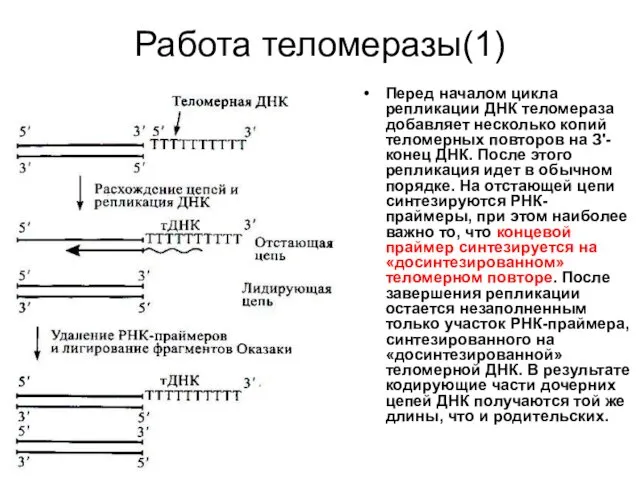
Работа теломеразы(1)
Перед началом цикла репликации ДНК теломераза добавляет несколько копий теломерных
Работа теломеразы(1)
Перед началом цикла репликации ДНК теломераза добавляет несколько копий теломерных

Удлинение теломер теломеразой у тетрахимены
Теломераза обладает своей молекулой РНК, имеющей матричный
Удлинение теломер теломеразой у тетрахимены
Теломераза обладает своей молекулой РНК, имеющей матричный
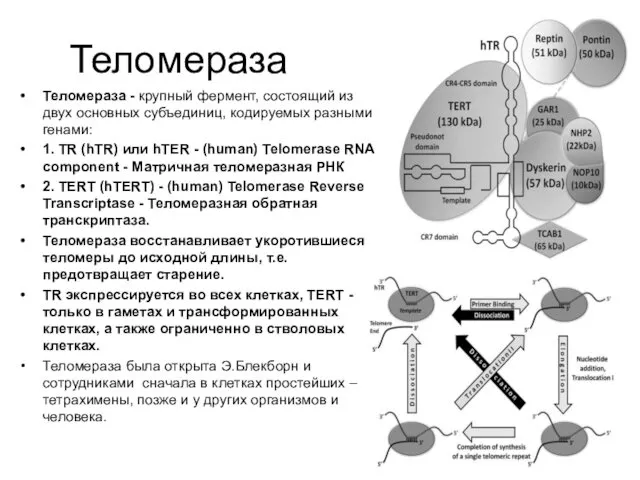
Теломераза
Теломераза - крупный фермент, состоящий из двух основных субъединиц, кодируемых разными
Теломераза
Теломераза - крупный фермент, состоящий из двух основных субъединиц, кодируемых разными

Роль теломеразной РНК
1. Изменение специфических оснований у Тетрахимены или дрожжей
Роль теломеразной РНК
1. Изменение специфических оснований у Тетрахимены или дрожжей

hnRNP
hnRNPs – heterogeneous nuclear ribonucleoproteins – обширное семейство связывющихся с
hnRNP
hnRNPs – heterogeneous nuclear ribonucleoproteins – обширное семейство связывющихся с
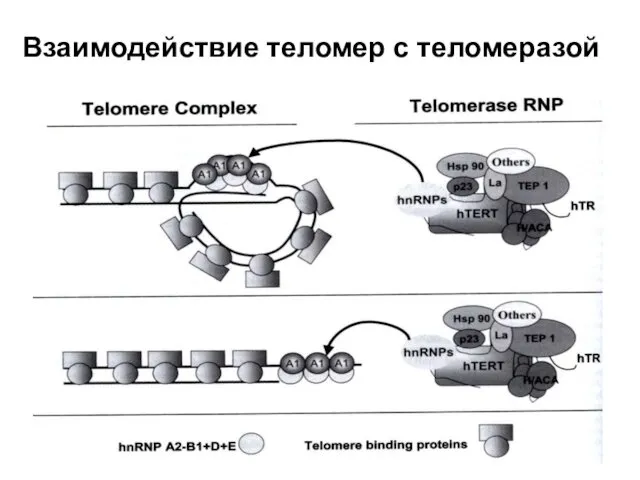
Взаимодействие теломер с теломеразой
Взаимодействие теломер с теломеразой
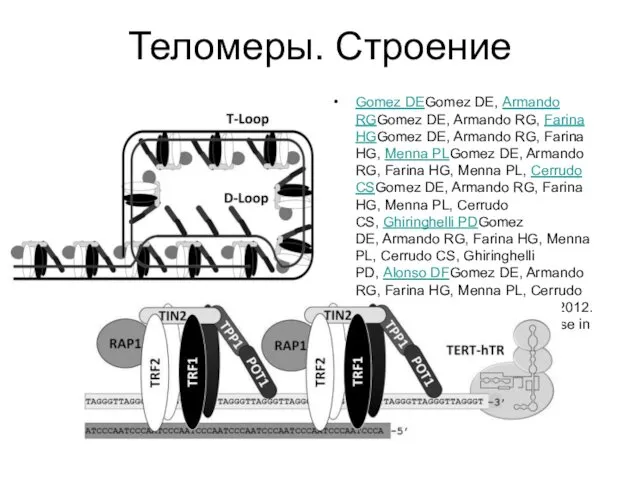
Теломеры. Строение
Gomez DEGomez DE, Armando RGGomez DE, Armando RG, Farina HGGomez DE, Armando RG, Farina HG, Menna
Теломеры. Строение
Gomez DEGomez DE, Armando RGGomez DE, Armando RG, Farina HGGomez DE, Armando RG, Farina HG, Menna
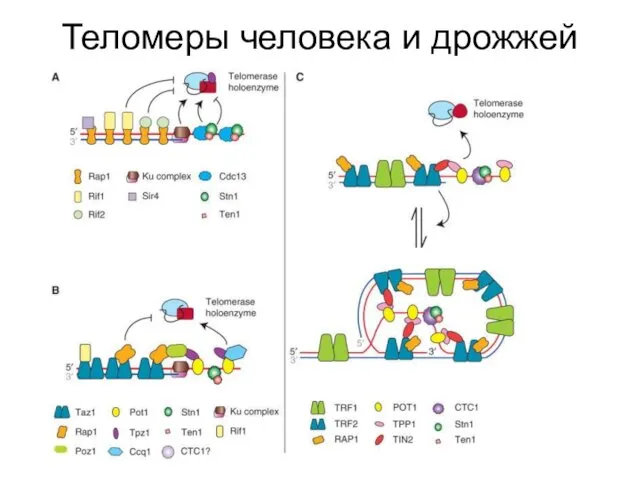
Теломеры человека и дрожжей
Теломеры человека и дрожжей
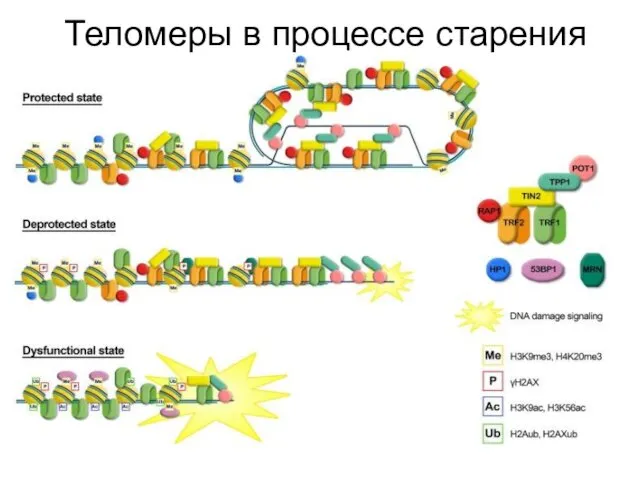
Теломеры в процессе старения
Теломеры в процессе старения
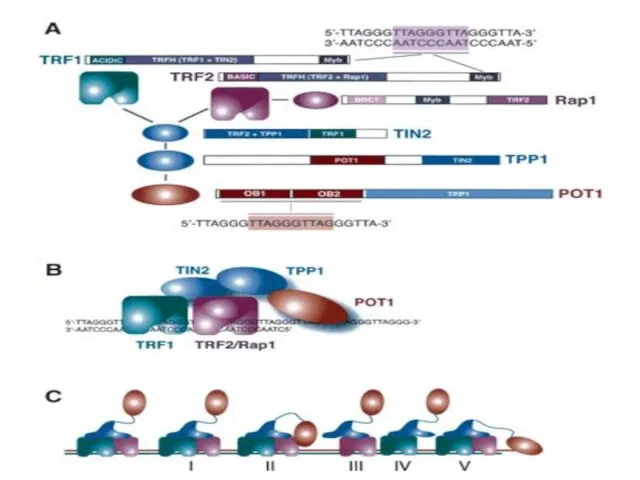
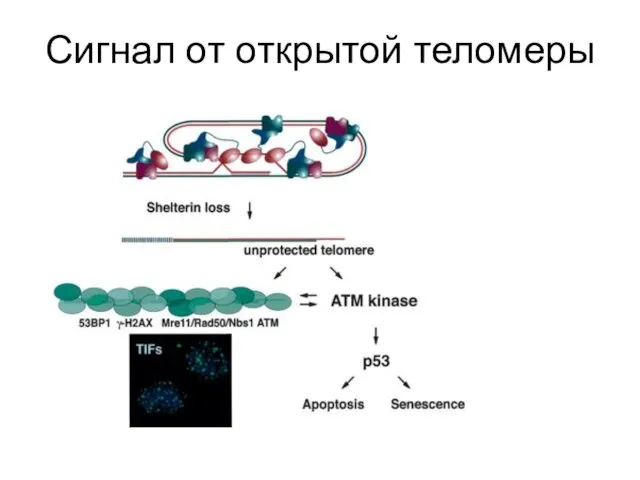
Сигнал от открытой теломеры
Сигнал от открытой теломеры
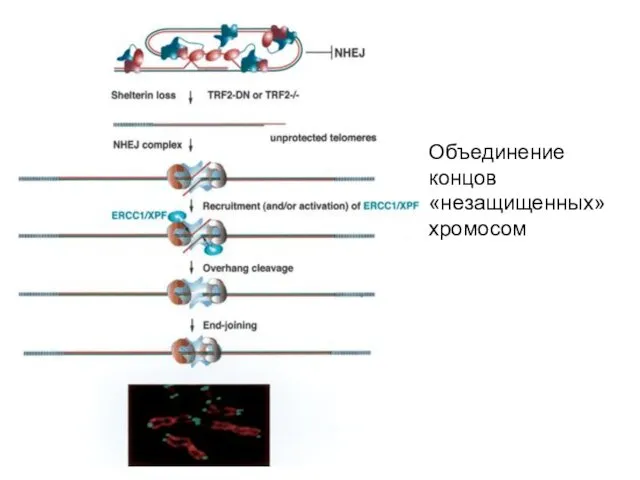
Объединение концов «незащищенных» хромосом
Объединение концов «незащищенных» хромосом
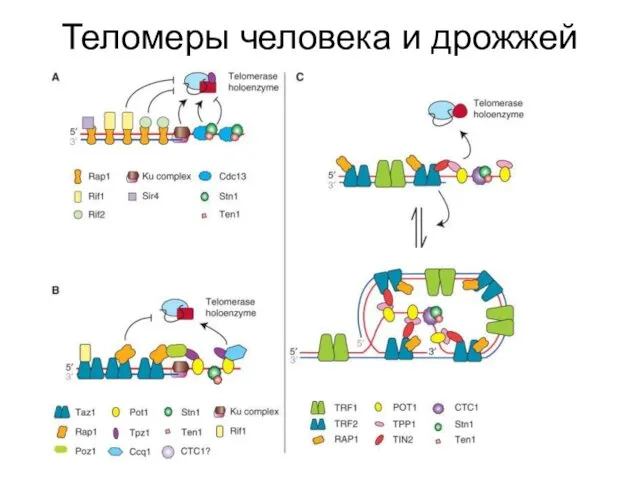
Теломеры человека и дрожжей
Теломеры человека и дрожжей

Белки, связанные с теломерами
1. Поддерживающие пространственную структуру теломер - семейство TRF
Белки, связанные с теломерами
1. Поддерживающие пространственную структуру теломер - семейство TRF
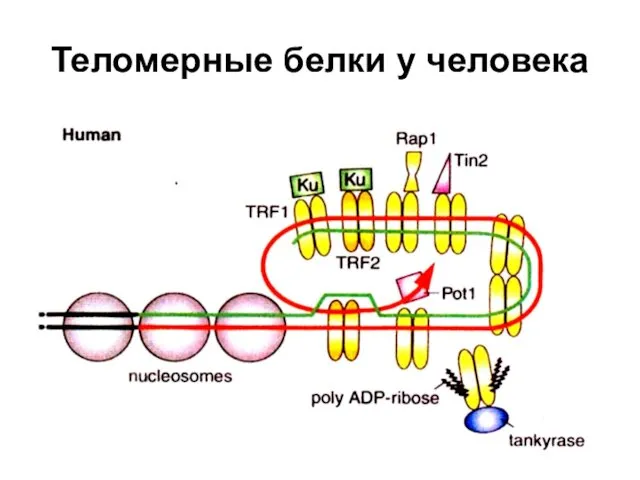
Теломерные белки у человека
Теломерные белки у человека

Белки, связанные с теломерами в клетках человека
ТRF1 - Теlоmеге repeat binding
Белки, связанные с теломерами в клетках человека
ТRF1 - Теlоmеге repeat binding
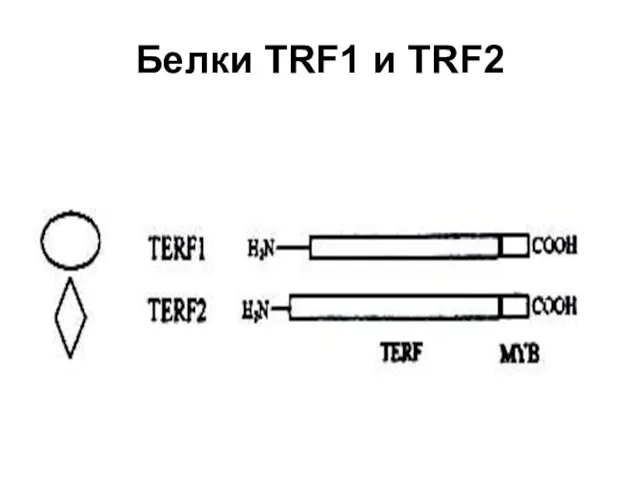
Белки TRF1 и TRF2
Белки TRF1 и TRF2

TRF 1
Резкое увеличение концентрации ТRF1 при его экспрессии в культуре клеток
TRF 1
Резкое увеличение концентрации ТRF1 при его экспрессии в культуре клеток

TRF 1
Первое сообщение о белке TRF 1 (Теlomeric repeat binding factor
TRF 1
Первое сообщение о белке TRF 1 (Теlomeric repeat binding factor

ТRF2
Белок ТRF2 был найден в конце 90-х годов. По структуре он
ТRF2
Белок ТRF2 был найден в конце 90-х годов. По структуре он

ТRF2
Пороговая для сенильного торможения пролиферации длина теломер у контрольных клеток составляет
ТRF2
Пороговая для сенильного торможения пролиферации длина теломер у контрольных клеток составляет

TNKS и TNKL
У человека и позвоночных животных имеется два изозима танкиразы,
TNKS и TNKL
У человека и позвоночных животных имеется два изозима танкиразы,

TRF 1
ТRF1может быть поли-АDР-рибозилирован, что сопровождается его диссоциацией от ДНК. Эту
TRF 1
ТRF1может быть поли-АDР-рибозилирован, что сопровождается его диссоциацией от ДНК. Эту

Белки, взаимодействующие с TRF1
Теломерный белок TINF2 или ТIN2 (ТRF1 intracting nuclear
Белки, взаимодействующие с TRF1
Теломерный белок TINF2 или ТIN2 (ТRF1 intracting nuclear

Теломерная теория старения
объясняет многие давно известные, но без нее непонятные факты
1.
Теломерная теория старения
объясняет многие давно известные, но без нее непонятные факты
1.

Трудности теломерной теории
1. Теломеры в клетках мыши ~ в 10 раз
Трудности теломерной теории
1. Теломеры в клетках мыши ~ в 10 раз

Возможное объяснение трудностей теломерной теории старения
Для старения и смерти от старости
Возможное объяснение трудностей теломерной теории старения
Для старения и смерти от старости

Репарационные белки, связанные с теломерами
Кu (состоит из субъединиц Кu 70 и
Репарационные белки, связанные с теломерами
Кu (состоит из субъединиц Кu 70 и
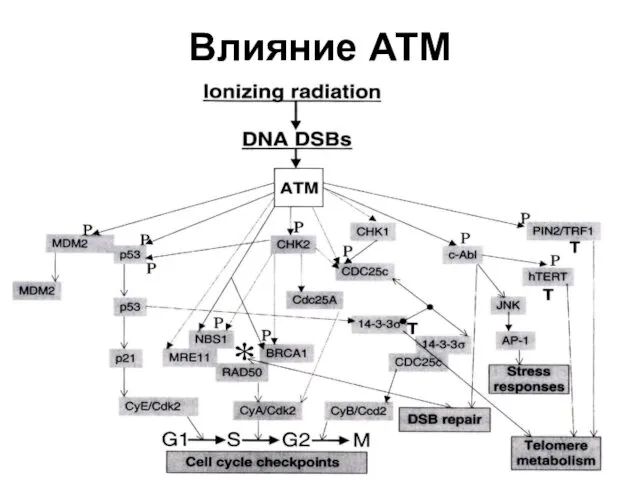
Влияние АТМ
Влияние АТМ
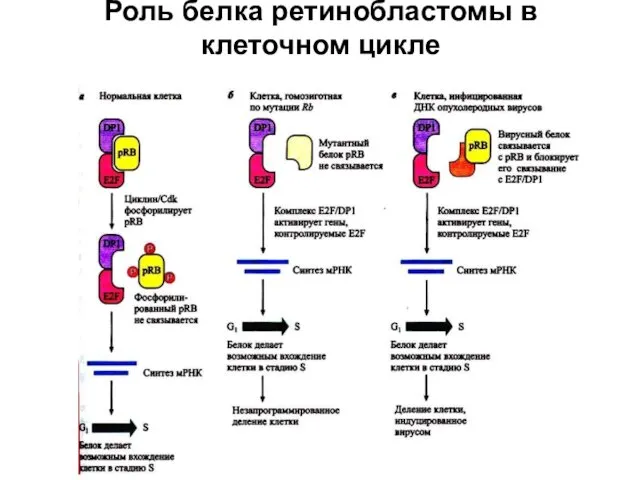
Роль белка ретинобластомы в клеточном цикле
Роль белка ретинобластомы в клеточном цикле
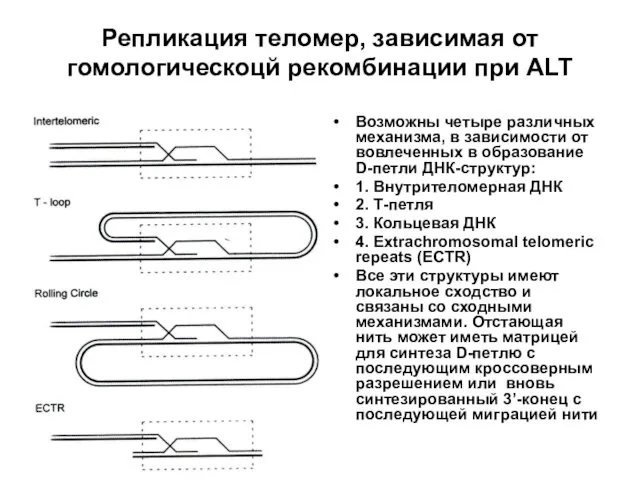
Репликация теломер, зависимая от гомологическоцй рекомбинации при ALT
Возможны четыре различных механизма,
Репликация теломер, зависимая от гомологическоцй рекомбинации при ALT
Возможны четыре различных механизма,

Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных
Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных

Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных
Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных

Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных
Белки, вовлеченные в процесс альтернативного удлинения теломер и находящиеся во внутриядерных

Белки, связанные с теломерами
1. Поддерживающие пространственную структуру теломер - семейство TRF
Белки, связанные с теломерами
1. Поддерживающие пространственную структуру теломер - семейство TRF
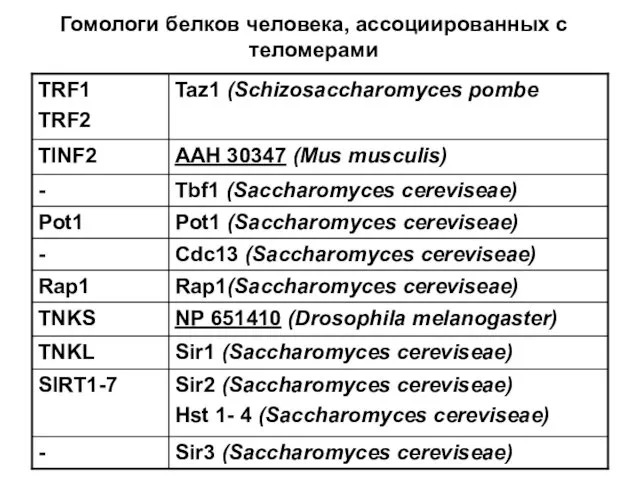
Гомологи белков человека, ассоциированных с теломерами
Гомологи белков человека, ассоциированных с теломерами
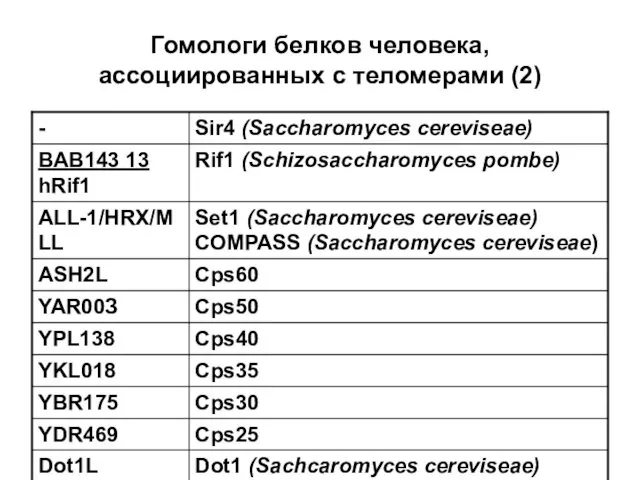
Гомологи белков человека, ассоциированных с теломерами (2)
Гомологи белков человека, ассоциированных с теломерами (2)
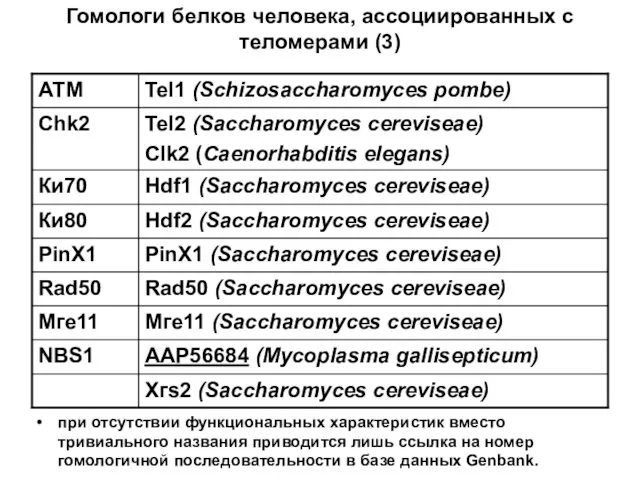
Гомологи белков человека, ассоциированных с теломерами (3)
при отсутствии функциональных характеристик вместо
Гомологи белков человека, ассоциированных с теломерами (3)
при отсутствии функциональных характеристик вместо

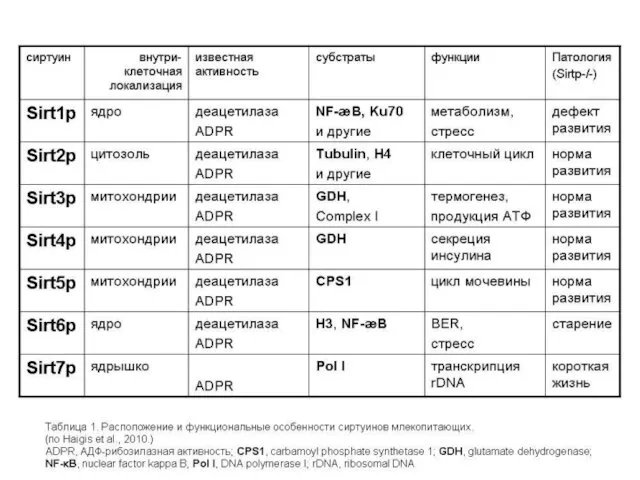
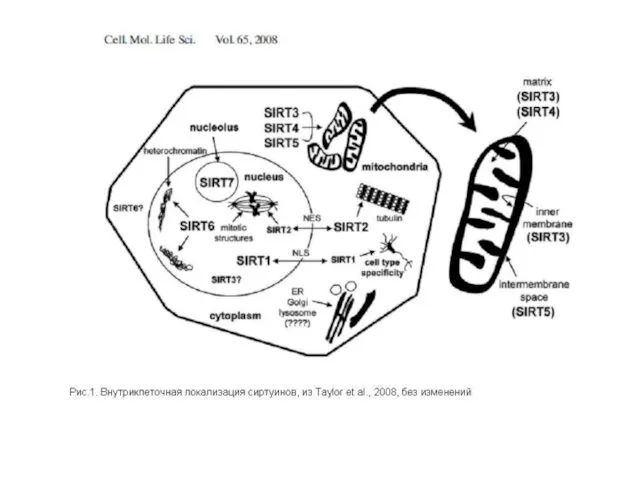
SIRT-белки (сиртуины)
Известно, что в клетках каждой ткани функционируют только те гены,
SIRT-белки (сиртуины)
Известно, что в клетках каждой ткани функционируют только те гены,

Недорепарация теломер
теломерная ДНК повреждается в среднем сильней, чем ДНК из внутренних
Недорепарация теломер
теломерная ДНК повреждается в среднем сильней, чем ДНК из внутренних

Причины недорепарации
1. Репарация концов ДНК может встречать те же трудности, что
Причины недорепарации
1. Репарация концов ДНК может встречать те же трудности, что
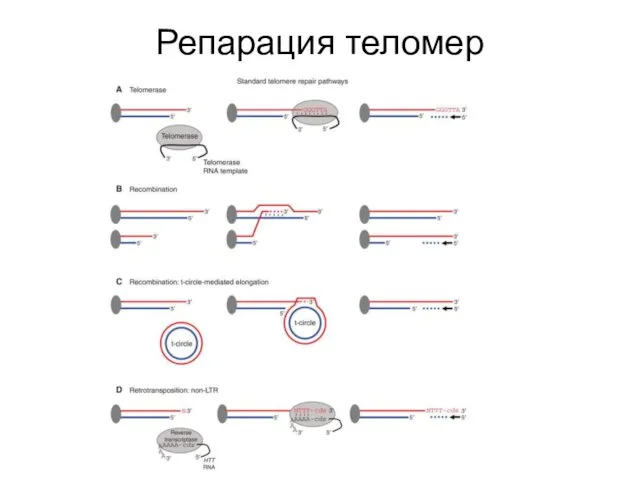
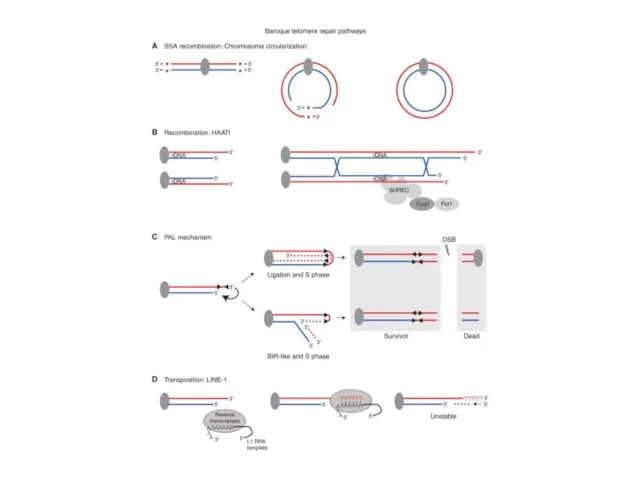
Репарация теломер
Репарация теломер
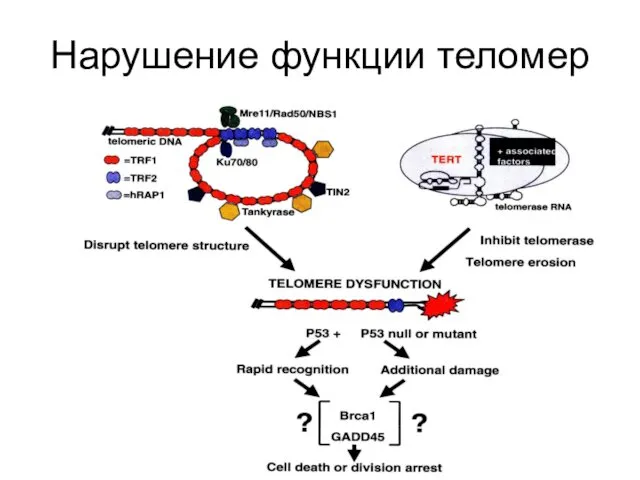
Нарушение функции теломер
Нарушение функции теломер
 Обмін речовин та енергії
Обмін речовин та енергії Заповідники України
Заповідники України Организм человека
Организм человека Презентация на тему "Класс паукообразных" - скачать презентации по Биологии
Презентация на тему "Класс паукообразных" - скачать презентации по Биологии Водоросли, мхи, лишайники М.Ф. Рженичева Источник материалов: интернет – ресурсы Детская «почемучка», ссылка указана http://www.pozn
Водоросли, мхи, лишайники М.Ф. Рженичева Источник материалов: интернет – ресурсы Детская «почемучка», ссылка указана http://www.pozn Ларвальные цестодозы
Ларвальные цестодозы Клетка. Строение
Клетка. Строение Общая характеристика. Внешнее строение. Среды жизни и места обитания.
Общая характеристика. Внешнее строение. Среды жизни и места обитания. Органы и системы органов
Органы и системы органов Генетика. Независимое наследование
Генетика. Независимое наследование Теория панспермии. Гипотеза случайного зарождения жизни
Теория панспермии. Гипотеза случайного зарождения жизни Уровни организации жизни Основные свойства живого. Урок по биологии Подготовила учитель биологии Пискунова О.А. МОУ СОШ №3
Уровни организации жизни Основные свойства живого. Урок по биологии Подготовила учитель биологии Пискунова О.А. МОУ СОШ №3 Биохимия соединительной ткани
Биохимия соединительной ткани Тема: Световая фаза фотосинтеза
Тема: Световая фаза фотосинтеза Отдел Моховидные, особенности строения и жизнедеятельности. Значение мхов
Отдел Моховидные, особенности строения и жизнедеятельности. Значение мхов Наследственность, изменчивость, фенотип, генотип, ген
Наследственность, изменчивость, фенотип, генотип, ген Тема: «Химический состав клетки. Неорганические вещества клетки» Задачи: Дать характеристику химическому составу клетки: группа
Тема: «Химический состав клетки. Неорганические вещества клетки» Задачи: Дать характеристику химическому составу клетки: группа Моногибридное скрещивание1
Моногибридное скрещивание1 Грибы
Грибы Презентация История развития жизни
Презентация История развития жизни Тип Членистоногие
Тип Членистоногие Презентация на тему "Лес просит о помощи!" - презентации по Биологии
Презентация на тему "Лес просит о помощи!" - презентации по Биологии Растительные сообщества
Растительные сообщества Ссавці. Парнокопитні. Жирафові. Жирафа степова
Ссавці. Парнокопитні. Жирафові. Жирафа степова  Домашние животные
Домашние животные Возникновение и развитие жизни на Земле
Возникновение и развитие жизни на Земле Митоз и мейоз
Митоз и мейоз Посвящается Чарльзу Дарвину к 200-летию со Дня его Рождения
Посвящается Чарльзу Дарвину к 200-летию со Дня его Рождения