- Типы взаимодействия. Хищничество

Содержание
- 2. Lepus arcticus
- 4. Lepus americanus 1860 1860
- 5. Bergerud A.T. 1983 Prey switching in simple ecosystem. Scientific American, 249, p. 116-124. Lynx canadensis Bergerud
- 6. Виды Тип взаимодействия А B Конкуренция - - Аменсализм 0 - Нейтрализм 0 0 Хищничество +
- 7. Питты Кат-Тьена Pitta ellioti полосатобрюхая Pitta soror синепоясничная Pitta moluccensis молуккская
- 10. \
- 13. Коллективная охота стаи волков на лося на острове Айл Роял
- 15. dN/dt=rN[(K-N)/K], где r=r max- врожденная скорость роста популяции К- предельная плотность насыщения, N - численность К
- 16. УРАВНЕНИЯ ХИЩНИЧЕСТВА (Lotka, 1925; Volterra, 1926, 1931) dN1/dt=r1N1-p1N1N2, dN2/dt=p2N1N2-d2N2, где N1 - плотность популяции жертвы, N2
- 17. ГИПОТЕТИЧЕСКАЯ ИЗОКЛИНА ЖЕРТВЫ (dN/dt=0). Внутри заштрихованной области плотность жертвы возрастает, а снаружи падает При средней плотности
- 18. НЕЭФФЕКТИВНЫЙ ХИЩНИК. Колебания хищника и жертвы затухают. Система стремится к устойчивому равновесию Точка равновесия ВЫСОКОЭФФЕКТИВНЫЙ ХИЩНИК
- 19. РОСТ ИНФУЗОРИИ-ЖЕРТВЫ PARAMECIUM CAUDATUM И ХИЩНОЙ ИНФУЗОРИИ DADINIUM NASUTUM В ОДНОРОДНОЙ И РАЗНОРОДНОЙ СРЕДАХ (GAUSE, 1934)
- 20. КОЛЕБАНИЯ ЧИСЛЕННОСТИ ХИЩНОГО КЛЕЩА THYPHLODROMUS OCCIDENTALIS И ЕГО ЖЕРТВЫ - КЛЕЩА EOTETRANYCHYS SEXMACULATUS Опыт с 40
- 21. Опыт с 252 апельсинами, из которых только каждый двадцатый был доступен для клещей. КОЛЕБАНИЯ ЧИСЛЕННОСТИ ХИЩНОГО
- 22. Было получено 112 поколений 14 полных циклов КОЛЕБАНИЯ ЧИСЛЕННОСТИ ПАРАЗИТОЙДА HETEROSPILUS PROSOPIDYS И ЕГО ЖЕРТВЫ –
- 23. КОЭВОЛЮЦИЯ ХИЩНИКА И ЖЕРТВЫ Миллионы лет Увеличение разнообразия таксонов хищников, способных разрушать толстостенные раковины молюсков, происходило
- 24. КРИПТИЧЕСКИЕ ЖЕРТВЫ Обыкновенная игуана Iguana iguana
- 25. Лентохвостый дипло- дактилюс (Diplodactylus taenicauda) из гекконов Восточная Австралия
- 26. Земляной (шишкохвостый) Nephrurus wheeleri (Западная Австралия)
- 27. Листохвостый геккон Phyllurus platurus (Юго-восточная Австралия)
- 28. РАСЧЛЕНЯЮЩАЯ ОКРАСКА Капская игольная змея (Mehelya capensis)
- 29. FURCIFER MINOR
- 30. Изменчивость количества и размера пятен у гуппи (Poecilia reticulata) в зависимости от количества хищников на трансекте
- 31. КРИПТИЧЕСКИЕ ХИЩНИКИ
- 32. РОГАТАЯ ГАДЮКА АВИЦЕНЫ (CERASTES VIPERA)
- 33. Жерлянкоподобная лягушка ADELOTUS BREVIS (Побережье Восточной Австралии)
- 34. ВРЕМЯ Вероятость обнаружения КРИПТИЧЕСКАЯ ЖЕРТВА ХИЩНИК: ЗАТРАЧИВАЕМОЕ НА ПОИСК КРИПТИЧЕСКИ ОКРАШЕННОЙ ЖЕРТВЫ БОЛЬШЕ ВРЕМЕНИ МЕХАНИЗМЫ: а)
- 35. АПОСТАТИЧЕСКОЕ ХИЩНИЧЕСТВО Хищник предпочитает диспропорционально чаще многочисленную жертву малочисленной. Цыпленок выбирает из смешанной популяции зеленых и
- 36. CRINIA SINGIFERA (Юго-восток Австралии) Внутрипопуляционный полиморфизм окраски
- 37. АПОСЕМАТИЧЕСКАЯ ОКРАСКА
- 38. NOTADEN BENETTII (ЮГО-ВОСТОК АВСТРАЛИИ)
- 39. PSEUDOPHRYNE CORROBOREA (ЮГО-ВОСТОК АВСТРАЛИИ)
- 40. CHLAMIDOSAURUS KINGII ПЛАЩЕНОСНАЯ ЯЩЕРИЦА Северная Австралия
- 42. Phrynocephalus mystaceus
- 43. АПОСЕМАТИЧЕСКАЯ ОКРАСКА Предупреждающая (апосематическая) окраска встречается у несъедобных видов: 1. Апосематическая окраска вторична Несъедобность редка. 2.
- 44. ГИПОТЕЗЫ ЭВОЛЮЦИИ АПОСЕМАТИЧЕСКОЙ ОКРАСКИ СТРАТЕГИЯ ЖЕРТВЫ: 1. Поскольку жертвы часто ассоциированы и представлены родственными особями (гусеницы)
- 45. Механизмы, обуслословливающие избегание несъедобной жертвы хищником 1. Научение хишника всегда имеет важное значение в распознавании жертвы.
- 46. ПРЕИМУЩЕСТВО АГГРЕГИРОВАННОГО РАСПРЕДЕЛЕНИЯ ПИЩЕВЫХ ОБЪЕКТОВ С АПОСЕМАТИЧЕСКОЙ ОКРАСКОЙ Цыпленок быстрее научается избегать несъедобную “жертву”, если та
- 47. МИМИКРИЯ МЮЛЛЕРА - ОБА ВИДА ЖЕРТВ НЕСЪЕДОБНЫ (один более, чем другой) И ВЫИГРЫВАЮТ ОТ КОНВЕРГЕНЦИИ. ВАРИАНТ
- 48. МОДЕЛИ P. mephalion P. chamissonia P. perrhebus МОДЕЛЬ МИМИКРИРУЮЩИЕ ФОРМЫ PAPILIO LYSITHOUS Из Punnett 1915, Poulton,
- 49. Бэйтсова мимикрия у булавоусых Восточной Африки Ядовитые данаиды Самец не мимикр., Самки мимикр 1-й вид самец
- 50. H. sapho H. cydno H. erato H. melpomene Мюллерова мимикрия у бабочек Heliconius в Южной Америке.
- 51. ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (1) Число дней ПО ПЕРЕМЕЩЕНИЮ МИМИКРИРУЮЩЕЙ ФОРМЫ В ПЕРУ (Mallet, Barton,
- 52. ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (2) H petiverana В' Heliconius favorinus H petiverana М H. emma
- 53. Ядовитая модель из рода Micrurus – коралловые аспиды справа, имитаторы Pliocercus из ужовых Е – одновременная
- 55. Скачать презентацию

Lepus arcticus
Lepus arcticus


Lepus americanus
1860
1860
Lepus americanus
1860
1860

Bergerud A.T. 1983 Prey switching in simple ecosystem. Scientific American, 249,
Bergerud A.T. 1983 Prey switching in simple ecosystem. Scientific American, 249,

Виды
Тип взаимодействия А B
Конкуренция - -
Аменсализм 0 -
Нейтрализм 0 0
Хищничество + -
Паразитизм + -
Мимикрия Бейтса + -
Растительноядность + -
Комменсализм + 0
Протокооперация + +
(факультативный мутуализм)
Мимикрия Мюллера + +
Мутуализм + +
Виды
Тип взаимодействия А B
Конкуренция - -
Аменсализм 0 -
Нейтрализм 0 0
Хищничество + -
Паразитизм + -
Мимикрия Бейтса + -
Растительноядность + -
Комменсализм + 0
Протокооперация + +
(факультативный мутуализм)
Мимикрия Мюллера + +
Мутуализм + +

Питты
Кат-Тьена
Pitta ellioti
полосатобрюхая
Pitta soror
синепоясничная
Pitta moluccensis
молуккская
Питты
Кат-Тьена
Pitta ellioti
полосатобрюхая
Pitta soror
синепоясничная
Pitta moluccensis
молуккская



\
\



Коллективная охота стаи волков на лося на
острове Айл Роял
Коллективная охота стаи волков на лося на
острове Айл Роял

![dN/dt=rN[(K-N)/K], где r=r max- врожденная скорость роста популяции К- предельная плотность](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/516077/slide-14.jpg)
dN/dt=rN[(K-N)/K], где
r=r max- врожденная скорость
роста популяции К- предельная
плотность насыщения,
dN/dt=rN[(K-N)/K], где
r=r max- врожденная скорость
роста популяции К- предельная
плотность насыщения,

УРАВНЕНИЯ ХИЩНИЧЕСТВА (Lotka, 1925; Volterra, 1926, 1931)
dN1/dt=r1N1-p1N1N2,
dN2/dt=p2N1N2-d2N2,
где N1 - плотность популяции
УРАВНЕНИЯ ХИЩНИЧЕСТВА (Lotka, 1925; Volterra, 1926, 1931)
dN1/dt=r1N1-p1N1N2,
dN2/dt=p2N1N2-d2N2,
где N1 - плотность популяции

ГИПОТЕТИЧЕСКАЯ ИЗОКЛИНА ЖЕРТВЫ
(dN/dt=0). Внутри заштрихованной области
плотность жертвы возрастает, а снаружи
падает
При
(dN/dt=0). Внутри заштрихованной области
плотность жертвы возрастает, а снаружи
падает
При
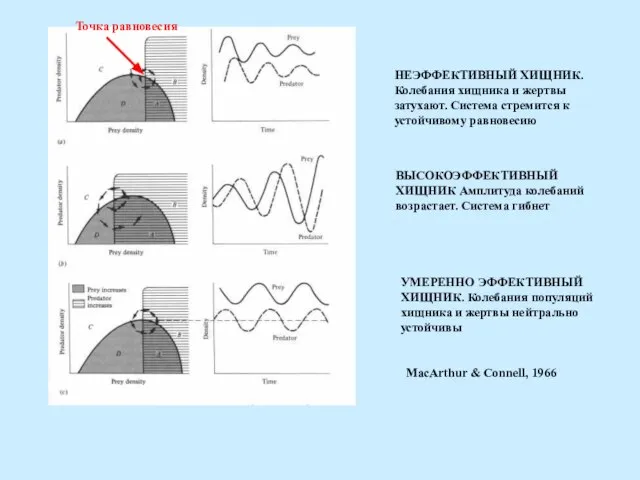
НЕЭФФЕКТИВНЫЙ ХИЩНИК.
Колебания хищника и жертвы
затухают. Система стремится к
устойчивому равновесию
Точка равновесия
ВЫСОКОЭФФЕКТИВНЫЙ
ХИЩНИК Амплитуда
НЕЭФФЕКТИВНЫЙ ХИЩНИК.
Колебания хищника и жертвы
затухают. Система стремится к
устойчивому равновесию
Точка равновесия
ВЫСОКОЭФФЕКТИВНЫЙ
ХИЩНИК Амплитуда
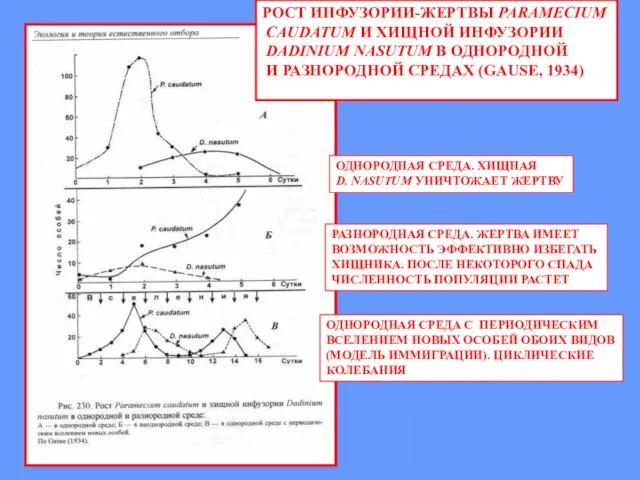
РОСТ ИНФУЗОРИИ-ЖЕРТВЫ PARAMECIUM
CAUDATUM И ХИЩНОЙ ИНФУЗОРИИ
DADINIUM NASUTUM В ОДНОРОДНОЙ
РОСТ ИНФУЗОРИИ-ЖЕРТВЫ PARAMECIUM
CAUDATUM И ХИЩНОЙ ИНФУЗОРИИ
DADINIUM NASUTUM В ОДНОРОДНОЙ
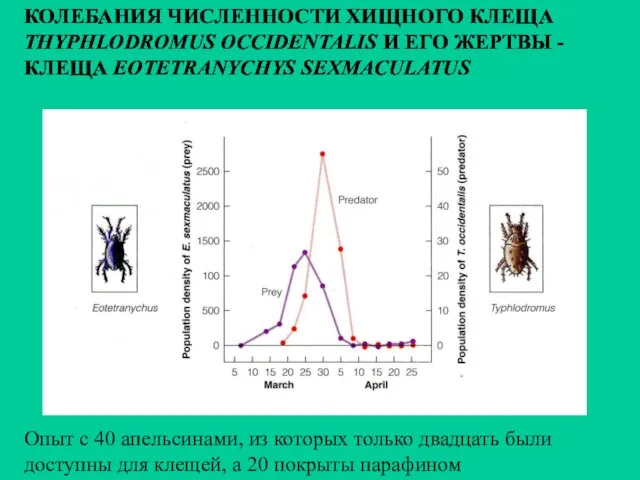
КОЛЕБАНИЯ ЧИСЛЕННОСТИ ХИЩНОГО КЛЕЩА
THYPHLODROMUS OCCIDENTALIS И ЕГО ЖЕРТВЫ -
КЛЕЩА EOTETRANYCHYS SEXMACULATUS
Опыт
КОЛЕБАНИЯ ЧИСЛЕННОСТИ ХИЩНОГО КЛЕЩА
THYPHLODROMUS OCCIDENTALIS И ЕГО ЖЕРТВЫ -
КЛЕЩА EOTETRANYCHYS SEXMACULATUS
Опыт
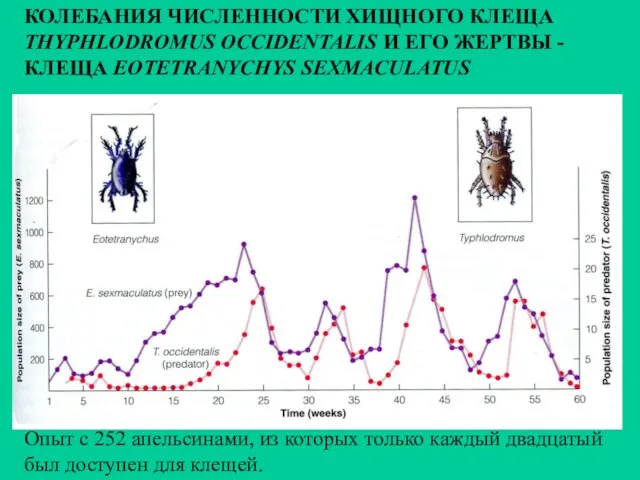
Опыт с 252 апельсинами, из которых только каждый двадцатый был доступен
Опыт с 252 апельсинами, из которых только каждый двадцатый был доступен
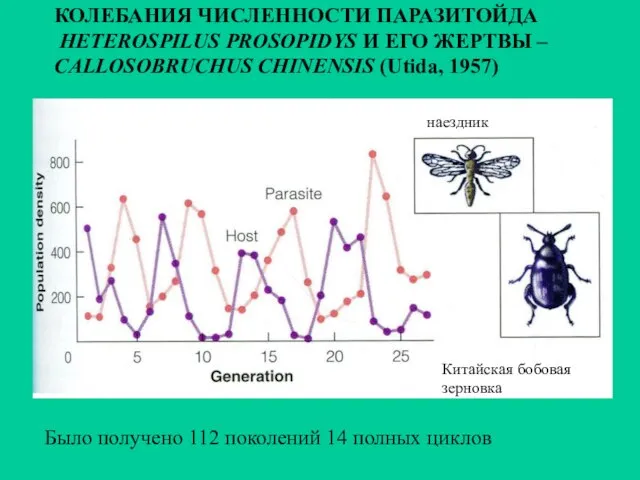
Было получено 112 поколений 14 полных циклов
КОЛЕБАНИЯ ЧИСЛЕННОСТИ ПАРАЗИТОЙДА
HETEROSPILUS PROSOPIDYS
Было получено 112 поколений 14 полных циклов
КОЛЕБАНИЯ ЧИСЛЕННОСТИ ПАРАЗИТОЙДА HETEROSPILUS PROSOPIDYS
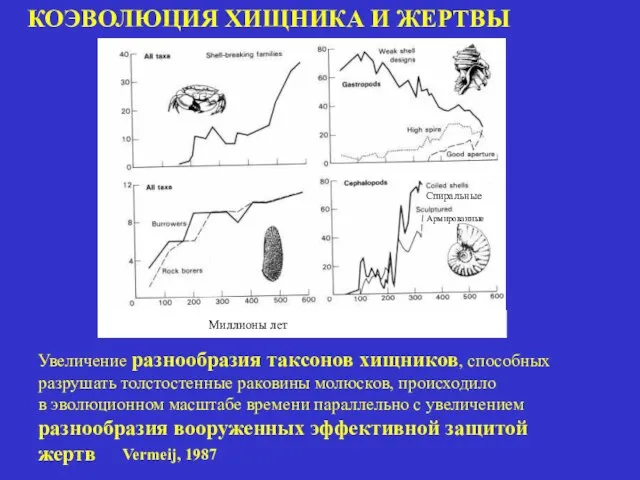
КОЭВОЛЮЦИЯ ХИЩНИКА И ЖЕРТВЫ
Миллионы лет
Увеличение разнообразия таксонов хищников, способных
разрушать толстостенные
КОЭВОЛЮЦИЯ ХИЩНИКА И ЖЕРТВЫ
Миллионы лет
Увеличение разнообразия таксонов хищников, способных
разрушать толстостенные

КРИПТИЧЕСКИЕ ЖЕРТВЫ
Обыкновенная игуана
Iguana iguana
КРИПТИЧЕСКИЕ ЖЕРТВЫ
Обыкновенная игуана
Iguana iguana

Лентохвостый дипло-
дактилюс
(Diplodactylus
taenicauda) из гекконов
Восточная Австралия
Лентохвостый дипло-
дактилюс
(Diplodactylus
taenicauda) из гекконов
Восточная Австралия

Земляной (шишкохвостый) Nephrurus wheeleri (Западная Австралия)
Земляной (шишкохвостый) Nephrurus wheeleri (Западная Австралия)

Листохвостый геккон Phyllurus platurus (Юго-восточная Австралия)
Листохвостый геккон Phyllurus platurus (Юго-восточная Австралия)

РАСЧЛЕНЯЮЩАЯ ОКРАСКА
Капская игольная змея (Mehelya capensis)
РАСЧЛЕНЯЮЩАЯ ОКРАСКА
Капская игольная змея (Mehelya capensis)

FURCIFER MINOR
FURCIFER MINOR

Изменчивость количества и размера
пятен у гуппи (Poecilia reticulata) в
зависимости от
Изменчивость количества и размера
пятен у гуппи (Poecilia reticulata) в
зависимости от

КРИПТИЧЕСКИЕ ХИЩНИКИ
КРИПТИЧЕСКИЕ ХИЩНИКИ

РОГАТАЯ ГАДЮКА АВИЦЕНЫ (CERASTES VIPERA)
РОГАТАЯ ГАДЮКА АВИЦЕНЫ (CERASTES VIPERA)

Жерлянкоподобная лягушка ADELOTUS BREVIS
(Побережье Восточной Австралии)
Жерлянкоподобная лягушка ADELOTUS BREVIS
(Побережье Восточной Австралии)
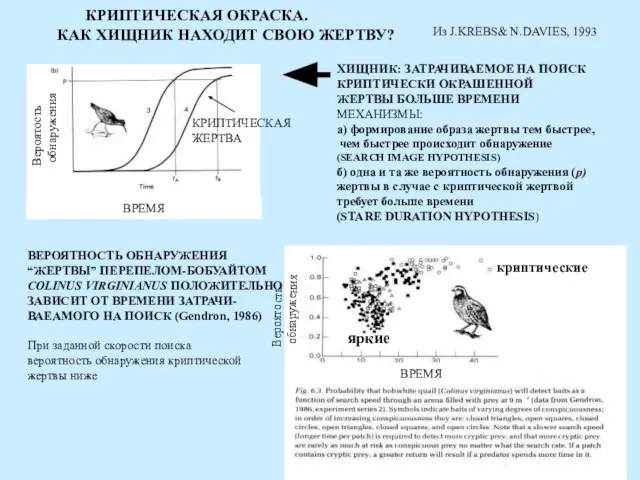
ВРЕМЯ
Вероятость
обнаружения
КРИПТИЧЕСКАЯ
ЖЕРТВА
ХИЩНИК: ЗАТРАЧИВАЕМОЕ НА ПОИСК
КРИПТИЧЕСКИ ОКРАШЕННОЙ
ЖЕРТВЫ БОЛЬШЕ ВРЕМЕНИ
МЕХАНИЗМЫ:
а) формирование
ВРЕМЯ
Вероятость
обнаружения
КРИПТИЧЕСКАЯ
ЖЕРТВА
ХИЩНИК: ЗАТРАЧИВАЕМОЕ НА ПОИСК
КРИПТИЧЕСКИ ОКРАШЕННОЙ
ЖЕРТВЫ БОЛЬШЕ ВРЕМЕНИ
МЕХАНИЗМЫ:
а) формирование
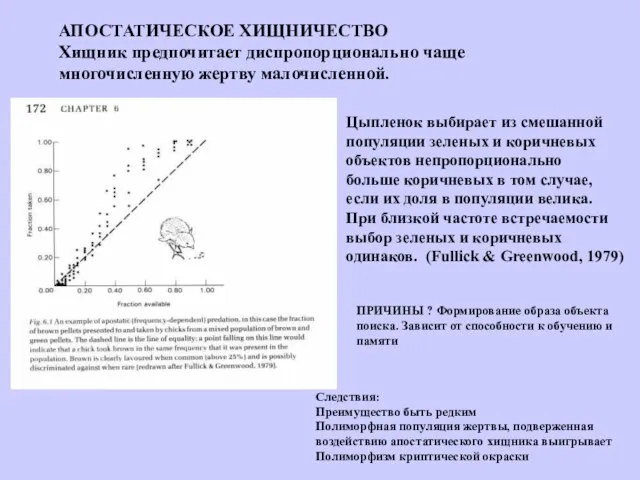
АПОСТАТИЧЕСКОЕ ХИЩНИЧЕСТВО
Хищник предпочитает диспропорционально чаще
многочисленную жертву малочисленной.
Цыпленок выбирает из смешанной
популяции зеленых
АПОСТАТИЧЕСКОЕ ХИЩНИЧЕСТВО
Хищник предпочитает диспропорционально чаще
многочисленную жертву малочисленной.
Цыпленок выбирает из смешанной
популяции зеленых

CRINIA SINGIFERA (Юго-восток Австралии)
Внутрипопуляционный полиморфизм окраски
CRINIA SINGIFERA (Юго-восток Австралии)
Внутрипопуляционный полиморфизм окраски

АПОСЕМАТИЧЕСКАЯ ОКРАСКА
АПОСЕМАТИЧЕСКАЯ ОКРАСКА

NOTADEN BENETTII (ЮГО-ВОСТОК АВСТРАЛИИ)
NOTADEN BENETTII (ЮГО-ВОСТОК АВСТРАЛИИ)

PSEUDOPHRYNE CORROBOREA (ЮГО-ВОСТОК АВСТРАЛИИ)
PSEUDOPHRYNE CORROBOREA (ЮГО-ВОСТОК АВСТРАЛИИ)

CHLAMIDOSAURUS KINGII
ПЛАЩЕНОСНАЯ ЯЩЕРИЦА
Северная Австралия
CHLAMIDOSAURUS KINGII
ПЛАЩЕНОСНАЯ ЯЩЕРИЦА
Северная Австралия


Phrynocephalus mystaceus
Phrynocephalus mystaceus

АПОСЕМАТИЧЕСКАЯ ОКРАСКА
Предупреждающая (апосематическая) окраска
встречается у несъедобных видов:
1. Апосематическая окраска вторична
АПОСЕМАТИЧЕСКАЯ ОКРАСКА
Предупреждающая (апосематическая) окраска
встречается у несъедобных видов:
1. Апосематическая окраска вторична

ГИПОТЕЗЫ ЭВОЛЮЦИИ АПОСЕМАТИЧЕСКОЙ
ОКРАСКИ
СТРАТЕГИЯ ЖЕРТВЫ:
1. Поскольку жертвы часто ассоциированы
ГИПОТЕЗЫ ЭВОЛЮЦИИ АПОСЕМАТИЧЕСКОЙ
ОКРАСКИ
СТРАТЕГИЯ ЖЕРТВЫ:
1. Поскольку жертвы часто ассоциированы

Механизмы, обуслословливающие избегание
несъедобной жертвы хищником
1. Научение хишника всегда имеет
Механизмы, обуслословливающие избегание
несъедобной жертвы хищником
1. Научение хишника всегда имеет
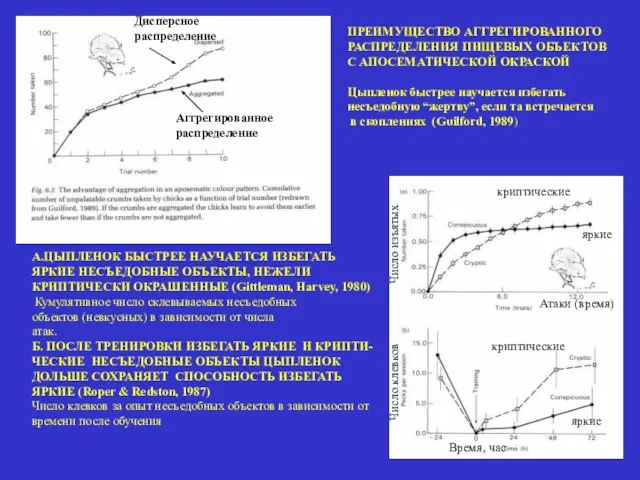
ПРЕИМУЩЕСТВО АГГРЕГИРОВАННОГО
РАСПРЕДЕЛЕНИЯ ПИЩЕВЫХ ОБЪЕКТОВ
С АПОСЕМАТИЧЕСКОЙ ОКРАСКОЙ
Цыпленок быстрее научается избегать
ПРЕИМУЩЕСТВО АГГРЕГИРОВАННОГО
РАСПРЕДЕЛЕНИЯ ПИЩЕВЫХ ОБЪЕКТОВ
С АПОСЕМАТИЧЕСКОЙ ОКРАСКОЙ
Цыпленок быстрее научается избегать

МИМИКРИЯ МЮЛЛЕРА - ОБА ВИДА ЖЕРТВ НЕСЪЕДОБНЫ (один более, чем
другой)
МИМИКРИЯ МЮЛЛЕРА - ОБА ВИДА ЖЕРТВ НЕСЪЕДОБНЫ (один более, чем
другой)
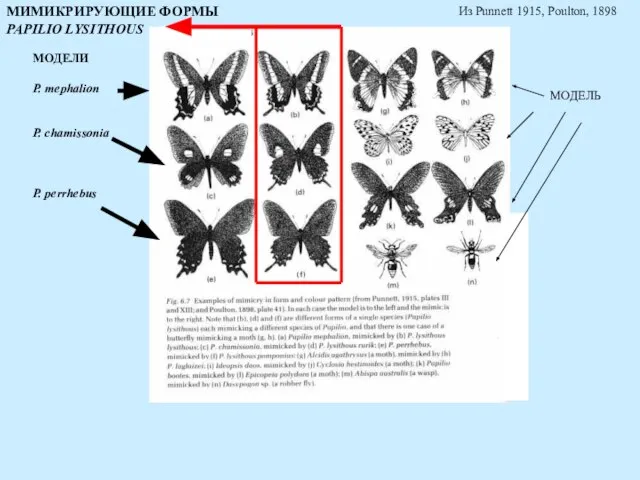
МОДЕЛИ
P. mephalion
P. chamissonia
P. perrhebus
МОДЕЛЬ
МИМИКРИРУЮЩИЕ ФОРМЫ
PAPILIO LYSITHOUS
Из Punnett 1915, Poulton, 1898
МОДЕЛИ
P. mephalion
P. chamissonia
P. perrhebus
МОДЕЛЬ
МИМИКРИРУЮЩИЕ ФОРМЫ
PAPILIO LYSITHOUS
Из Punnett 1915, Poulton, 1898

Бэйтсова мимикрия у булавоусых Восточной Африки
Ядовитые данаиды
Самец
не мимикр.,
Самки
мимикр
1-й вид
самец
не
Бэйтсова мимикрия у булавоусых Восточной Африки
Ядовитые данаиды
Самец
не мимикр.,
Самки
мимикр
1-й вид
самец
не
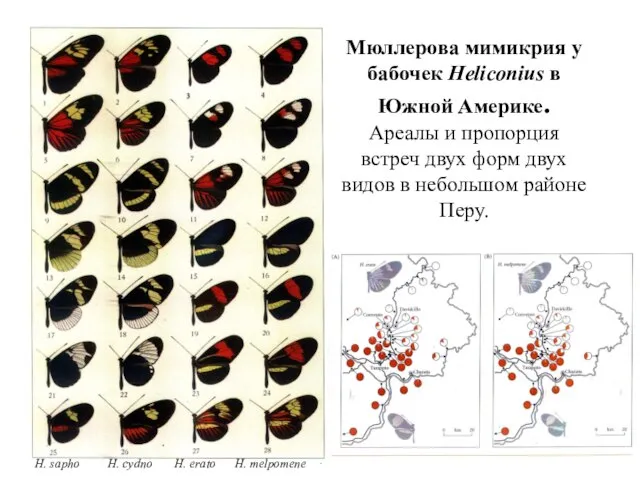
H. sapho H. cydno H. erato H. melpomene
Мюллерова мимикрия у
H. sapho H. cydno H. erato H. melpomene
Мюллерова мимикрия у

ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (1)
Число дней
ПО ПЕРЕМЕЩЕНИЮ МИМИКРИРУЮЩЕЙ ФОРМЫ В
ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (1)
Число дней
ПО ПЕРЕМЕЩЕНИЮ МИМИКРИРУЮЩЕЙ ФОРМЫ В

ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (2)
H petiverana
В'
Heliconius
favorinus
H petiverana
М
H. emma
Число дней
С
ЭКСПЕРИМЕНТ ПО ВЫЖИВАЕМОСТИ ГЕЛИКОНОВЫХ БАБОЧЕК (2)
H petiverana
В'
Heliconius
favorinus
H petiverana
М
H. emma
Число дней
С
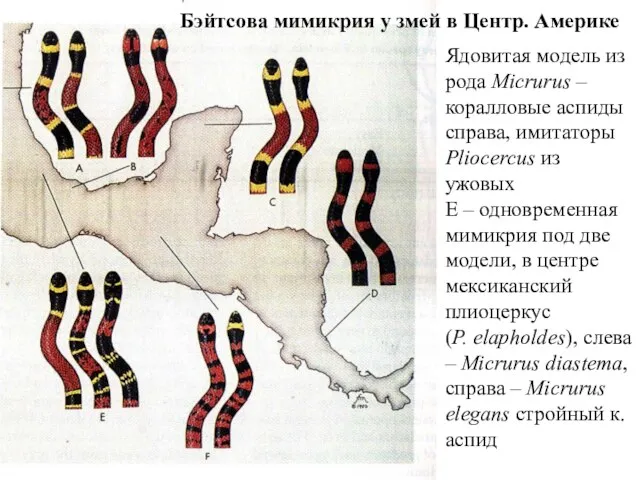
Ядовитая модель из рода Micrurus – коралловые аспиды
справа, имитаторы Pliocercus
Ядовитая модель из рода Micrurus – коралловые аспиды
справа, имитаторы Pliocercus
 Онто-филогенетические процессы при формировании скелета, мышц и кожи человека. ВПР костно-мышечной системы, кожи и ее придатков
Онто-филогенетические процессы при формировании скелета, мышц и кожи человека. ВПР костно-мышечной системы, кожи и ее придатков Учительский проект по биологии для 7 класса «Простейшие или Вторжение в тайны невидимок» Автор: Меделяев Дмитрий Борисович, уч
Учительский проект по биологии для 7 класса «Простейшие или Вторжение в тайны невидимок» Автор: Меделяев Дмитрий Борисович, уч Тело человека. Скелет. Мышцы
Тело человека. Скелет. Мышцы Фізіологія, біохімія, генетика рослин та ландшафтний фітодизайн
Фізіологія, біохімія, генетика рослин та ландшафтний фітодизайн Презентация на тему "Биологические особенности раннецветущих растений" - скачать презентации по Биологии
Презентация на тему "Биологические особенности раннецветущих растений" - скачать презентации по Биологии Самые опасные животные для человека Человек не так силен, как сам о себе думает. В природе множество животных, способных нанест
Самые опасные животные для человека Человек не так силен, как сам о себе думает. В природе множество животных, способных нанест Грибы. История открытия грибов
Грибы. История открытия грибов Эндокринная система. Функции желез внутренней секреции
Эндокринная система. Функции желез внутренней секреции Пресмыкающиеся, или Рептилии (Reptilia)
Пресмыкающиеся, или Рептилии (Reptilia) Пернатый мир. Большая синица
Пернатый мир. Большая синица Презентация на тему "Животные живого уголка" - скачать бесплатно презентации по Биологии
Презентация на тему "Животные живого уголка" - скачать бесплатно презентации по Биологии Рослини-хижаки
Рослини-хижаки Фото нарцисса
Фото нарцисса Что же такое онтогенез?
Что же такое онтогенез? Helicobacter
Helicobacter Символ нашего сострадания, поддержки и надежды на будущее - будущее без СПИДа.
Символ нашего сострадания, поддержки и надежды на будущее - будущее без СПИДа. Природа и наша безопасность. Тест №40
Природа и наша безопасность. Тест №40 Пресноводные брюхоногие моллюски каналов окрестностей г. Тирасполя
Пресноводные брюхоногие моллюски каналов окрестностей г. Тирасполя Дыхательная система
Дыхательная система Они живут рядом. Растительный и животный мир Владимирской области
Они живут рядом. Растительный и животный мир Владимирской области Презентация на тему "Биология – наука о живом мире. Общие свойства живых организмов" - скачать презентации по Биологии
Презентация на тему "Биология – наука о живом мире. Общие свойства живых организмов" - скачать презентации по Биологии Презентация на тему "Первая медицинская помощь при кровотечениях" - скачать презентации по Биологии
Презентация на тему "Первая медицинская помощь при кровотечениях" - скачать презентации по Биологии The nervous system
The nervous system Общие принципы организации тканей. Эпителиальные ткани
Общие принципы организации тканей. Эпителиальные ткани Отдел Покрытосеменные (тема 31)
Отдел Покрытосеменные (тема 31) Кровеносная система
Кровеносная система  Семейство крестоцветные
Семейство крестоцветные Биология как наука
Биология как наука