- Важное свойство мембран

Содержание
- 2. Если сигнал воспринимается мембранными рецепторами, то схему передачи информации можно представить так: взаимодействие рецептора с сигнальной
- 3. Рецепторы По локализации различают мембранные, цитоплазматические и ядерные рецепторы. По другой классификации все рецепторы можно разделить
- 4. Рис. 5-29. Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные с ионными каналами, например
- 5. Рис. 5-30. Мембранная организация β2-адренорецептора. 1 - фрагмент рецептора, участвующий в связывании Gs-белка; 2, 3 -
- 6. Рецепторы адреналина - адренорецепторы Адренорецепторы различают по распределению в организме - центральные и периферические. Центральные адренорецепторы,
- 7. β-Адренорецепторы встречаются практически во всех тканях организма. Количество β-адренорецепторов, приходящееся на клетку, варьирует от 300 до
- 8. Рецепторы с тирозинкиназной активностью Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки по тирозину, подразделяют на 2
- 9. Каталитический центр тирозиновой протеинкиназы находится на внутриклеточных доменах β-субъединиц. В отсутствие гормона инсулиновые рецепторы не проявляют
- 10. РИС. 5-31. АКТИВАЦИЯ РЕЦЕПТОРА ИНСУЛИНА - ТИРОЗИНОВОЙ ПРОТЕИНКИНАЗЫ.
- 11. Рецепторы с гуанилатциклазной активностью Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из важных посредников внутриклеточной передачи
- 12. Рис. 5-32. Образование 3',5'-циклического ГМФ (цГМФ).
- 13. РИС. 5-33. РЕГУЛЯЦИЯ АКТИВНОСТИ МЕМБРАННОЙ (1) И ЦИТОЗОЛЬНОЙ (2) ГУАНИЛАТЦИКЛАЗЫ.
- 14. Мембраннсювязанная гуанилатциклаза - трансмембранный гликопротеин. Внутриклеточный домен гуанилатциклазы проявляет каталитическую активность, внеклеточный домен служит рецептором. Присоединение
- 15. Передача сигнала с помощью внутриклеточных рецепторов Передача сигнала липидорастворимых стероидных гормонов и тироксина возможна только при
- 17. Последовательность событий, приводящих к активации транскрипции: гормон проходит через двойной липидный слой клеточной мембраны. взаимодействие гормона
- 18. Цитокины — небольшие пептидные информационные молекулы. Цитокины имеют молекулярную массу, не превышающую 30 кD. Цитокин выделяется
- 19. Группировка цитокинов по механизму действия позволяет разделить цитокины на следующие группы: провоспалительные, обеспечивающие мобилизацию воспалительного ответа
- 20. Факторы роста В поддержании жизни высших организмов ключевую роль играет контроль пролиферации ,дифференцировки и направленного движения
- 22. Скачать презентацию

Если сигнал воспринимается мембранными рецепторами, то схему передачи информации можно представить
Если сигнал воспринимается мембранными рецепторами, то схему передачи информации можно представить

Рецепторы
По локализации различают мембранные, цитоплазматические и ядерные рецепторы. По другой
Рецепторы
По локализации различают мембранные, цитоплазматические и ядерные рецепторы. По другой

Рис. 5-29. Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные
Рис. 5-29. Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные

Рис. 5-30. Мембранная организация β2-адренорецептора. 1 - фрагмент рецептора, участвующий в связывании Gs-белка;
Рис. 5-30. Мембранная организация β2-адренорецептора. 1 - фрагмент рецептора, участвующий в связывании Gs-белка;

Рецепторы адреналина - адренорецепторы
Адренорецепторы различают по распределению в организме - центральные
Рецепторы адреналина - адренорецепторы
Адренорецепторы различают по распределению в организме - центральные

β-Адренорецепторы встречаются практически во всех тканях организма. Количество β-адренорецепторов, приходящееся на клетку,
β-Адренорецепторы встречаются практически во всех тканях организма. Количество β-адренорецепторов, приходящееся на клетку,

Рецепторы с тирозинкиназной активностью
Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки по
Рецепторы с тирозинкиназной активностью
Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки по

Каталитический центр тирозиновой протеинкиназы находится на внутриклеточных доменах β-субъединиц. В отсутствие
Каталитический центр тирозиновой протеинкиназы находится на внутриклеточных доменах β-субъединиц. В отсутствие
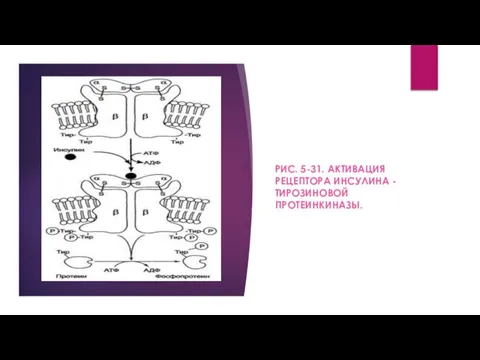
РИС. 5-31. АКТИВАЦИЯ РЕЦЕПТОРА ИНСУЛИНА - ТИРОЗИНОВОЙ ПРОТЕИНКИНАЗЫ.
РИС. 5-31. АКТИВАЦИЯ РЕЦЕПТОРА ИНСУЛИНА - ТИРОЗИНОВОЙ ПРОТЕИНКИНАЗЫ.

Рецепторы с гуанилатциклазной активностью
Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из
Рецепторы с гуанилатциклазной активностью
Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из
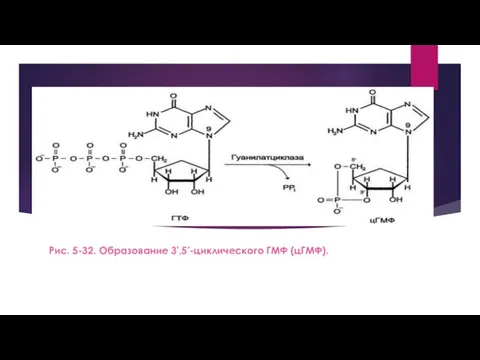
Рис. 5-32. Образование 3',5'-циклического ГМФ (цГМФ).
Рис. 5-32. Образование 3',5'-циклического ГМФ (цГМФ).
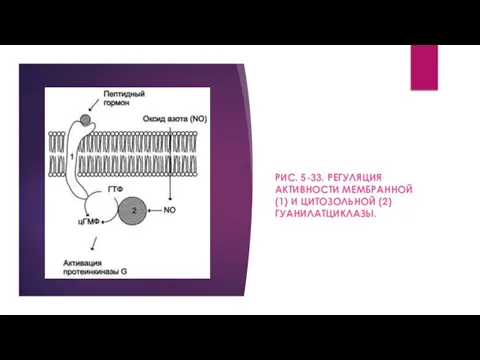
РИС. 5-33. РЕГУЛЯЦИЯ АКТИВНОСТИ МЕМБРАННОЙ (1) И ЦИТОЗОЛЬНОЙ (2) ГУАНИЛАТЦИКЛАЗЫ.
РИС. 5-33. РЕГУЛЯЦИЯ АКТИВНОСТИ МЕМБРАННОЙ (1) И ЦИТОЗОЛЬНОЙ (2) ГУАНИЛАТЦИКЛАЗЫ.

Мембраннсювязанная гуанилатциклаза - трансмембранный гликопротеин. Внутриклеточный домен гуанилатциклазы проявляет каталитическую активность,
Мембраннсювязанная гуанилатциклаза - трансмембранный гликопротеин. Внутриклеточный домен гуанилатциклазы проявляет каталитическую активность,

Передача сигнала с помощью внутриклеточных рецепторов
Передача сигнала липидорастворимых стероидных гормонов и
Передача сигнала с помощью внутриклеточных рецепторов
Передача сигнала липидорастворимых стероидных гормонов и
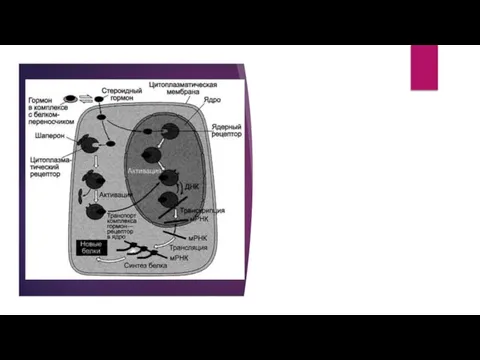

Последовательность событий, приводящих к активации транскрипции:
гормон проходит через двойной липидный слой
Последовательность событий, приводящих к активации транскрипции:
гормон проходит через двойной липидный слой

Цитокины — небольшие пептидные информационные молекулы. Цитокины имеют молекулярную массу, не превышающую 30 кD.
Цитокин
Цитокины — небольшие пептидные информационные молекулы. Цитокины имеют молекулярную массу, не превышающую 30 кD.
Цитокин

Группировка цитокинов по механизму действия позволяет разделить цитокины на следующие группы:
провоспалительные,
Группировка цитокинов по механизму действия позволяет разделить цитокины на следующие группы:
провоспалительные,

Факторы роста
В поддержании жизни высших организмов ключевую роль играет контроль пролиферации ,дифференцировки и направленного движения
Факторы роста
В поддержании жизни высших организмов ключевую роль играет контроль пролиферации ,дифференцировки и направленного движения
 Очень медленно идет, Дом она с собой несет. Испугается чего-то, В панцирь с головой уйдет
Очень медленно идет, Дом она с собой несет. Испугается чего-то, В панцирь с головой уйдет ВОЗНИКНОВЕНИЕ ЖИЗНИ
ВОЗНИКНОВЕНИЕ ЖИЗНИ  Физиология пищеварения
Физиология пищеварения Изоляция – эволюционный фактор
Изоляция – эволюционный фактор Основные понятия генетики. Моногибридное скрещивание
Основные понятия генетики. Моногибридное скрещивание Следы жизнедеятельности животных
Следы жизнедеятельности животных Решение задач по генетике Памятка для учащихся 9 – 10 классов Составила: Белова Римма Васильевна, учитель биологии ГБОУ СОШ с. Чёр
Решение задач по генетике Памятка для учащихся 9 – 10 классов Составила: Белова Римма Васильевна, учитель биологии ГБОУ СОШ с. Чёр Ихтиофауна памятника природы Долина реки Издревая
Ихтиофауна памятника природы Долина реки Издревая Тип Кишечнополостные
Тип Кишечнополостные Растения и животные луга УЧИТЕЛЬ Макарова Светлана Павловна средняя школа № 65 г.Кирова
Растения и животные луга УЧИТЕЛЬ Макарова Светлана Павловна средняя школа № 65 г.Кирова Биоразнообразие природы Московской области
Биоразнообразие природы Московской области Galápagos tortoise
Galápagos tortoise Мыши - семейство млекопитающих отряда грызунов
Мыши - семейство млекопитающих отряда грызунов Сельское хозяйство. Растениеводство.
Сельское хозяйство. Растениеводство. Пингвины
Пингвины Ұлулар
Ұлулар Кости и органы. Самостоятельная работа. Вариант 2
Кости и органы. Самостоятельная работа. Вариант 2 Презентация на тему "Дикие - дикие кошки" - скачать презентации по Биологии
Презентация на тему "Дикие - дикие кошки" - скачать презентации по Биологии Конечный мозг. Большие полушария. Кора
Конечный мозг. Большие полушария. Кора Презентация на тему "БЕЛКИ, КАК БИОПОЛИМЕРЫ, ИХ СОСТАВ" - скачать бесплатно презентации по Биологии
Презентация на тему "БЕЛКИ, КАК БИОПОЛИМЕРЫ, ИХ СОСТАВ" - скачать бесплатно презентации по Биологии Способы размножения растений
Способы размножения растений Ярмарка знаний. Цветок, плод, семя. 6 класс
Ярмарка знаний. Цветок, плод, семя. 6 класс Химические реакции с участием моносахаридов. Структура и функции ди-, олиго- и полисахаридов
Химические реакции с участием моносахаридов. Структура и функции ди-, олиго- и полисахаридов Предмет физиологии центральной нервной системы
Предмет физиологии центральной нервной системы Обитатели воды
Обитатели воды Чайный гриб
Чайный гриб Ткани растений
Ткани растений Завдання для екзамену з дисципліни “Ветеринарна мікробіологія”
Завдання для екзамену з дисципліни “Ветеринарна мікробіологія”