- Палеоген. Палеогеновый период

Содержание
- 2. Палеоге́новый пери́од— геологический период; первый период кайнозоя. Начался 66,0 млн лет назад, закончился 23,03 млн лет
- 3. В начале палеогена в Северном полушарии выделяются два крупных материка, соединявшихся в районе Берингова пролива -
- 4. В течение палеогенового периода образования располагались в континентальных условиях. Исключением были их окраинные части. Они испытывали
- 5. Она находилась несколько в иных условиях, чем Восточно-Европейская. На протяжении кайнозойской эры Сибирская платформа была представлена
- 6. Палеоклиматы суши палеогена были разнообразны Климатические особенности Рисунок - Схематическая глобальная палеоклиматическая карта эоценовой эпохи На
- 7. Начало кайнозойской эры связано с великим альпийским горообразованием (все высочайшие горные системы мира возникли приблизительно в
- 8. Необходимо акцентировать внимание на новых качествах, появившиеся у млекопитающих, которые могут считаться более прогрессивными, предопределившими их
- 9. Морская фауна
- 10. Моллюски в палеогене
- 11. Брюхоногие моллюски Брюхоногие – самая многообразная и распространенная группа моллюсков. Брюхоногих около 90 тыс. современных видов,
- 13. Костистые рыбы
- 14. 3 подотрядах китообразных Древние киты - Archeoceti; Усатые киты - Mysticeti; Зубатые киты - Odontoceti.
- 15. Древние киты - Archeoceti
- 16. Усатые киты - Mysticeti
- 17. Зубатые киты - Odontoceti
- 18. Акулы в палеогене
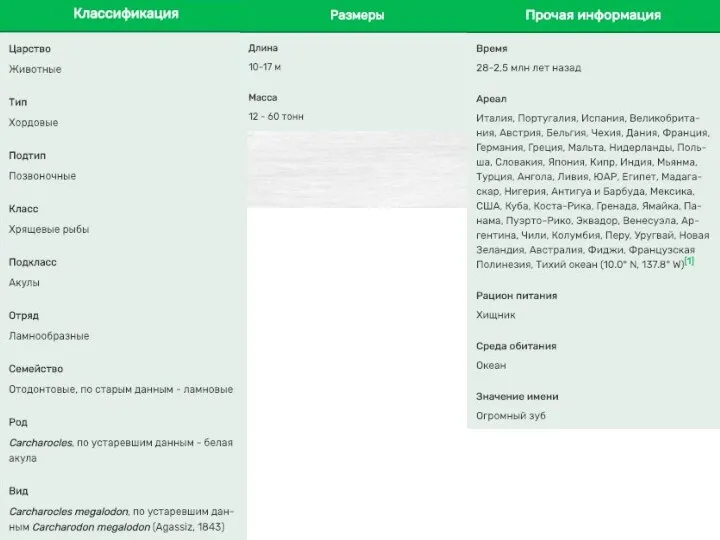
- 19. Мегалодон (лат. Carcharocles megalodon или Otodus megalodon, от греч. μέγας и ὀδούς — большой зуб) —
- 21. Anomotodon (лат.) – Аномотодоны — род вымерших акул из семейства скапаноринховых (Mitsukurinidae). Ископаемые остатки находят в
- 22. Кретоксирина (Cretoxyrhina) принадлежит к вымершему семейству Cretoxyrhinidae, входящему в современный отряд ламноидных акул (Lamniformes).
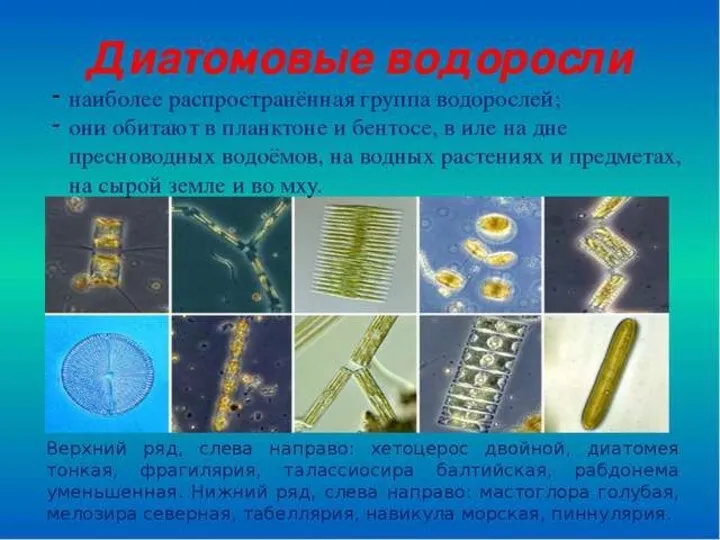
- 23. Водоросли в палеогене КОККОЛИТОФОРИДЫ (Coccolithophorea), класс одноклеточных, морских, планктонных гаптофитовых водорослей. Около 200 современных видов; наибольшего
- 24. Cretoxyrhinidae (лат.) — семейство вымерших акул отряда ламнообразных, живших во время барремского — лангского веков (130,0—13,82
- 26. Кораллы в палеогене СКЛЕРАКТИ́НИИ, мадрепоровые кораллы (Scleractinia, Madreporaria), отряд морских беспозвоночных подкласса шестилучевых кораллов. Наиболее разнообразны
- 28. Высота птицы 2,5 метра. Голова достигала 40 см в длину. Имелся массивный загнутый клюв. Птица обладала
- 29. Достигали в высоту 1,75 метра. Весили 100 кг. Имели массивный клюв. Длинные ноги, приспособленные для бега.
- 30. Высота птицы 1-3 метра. Были хищниками. На конце рудиментарных крыльев имели крюки. Имели мощный клюв, которым
- 31. Имели длинные ноги и шею. Строение черепа и клюва у них было уткоподобное. Предпочитали жить колониями
- 32. Oligocolius psittacocephalon. Череп птицы был похож на череп попугая. Предположительно был зоб. Pelagornis sandersi Размах крыльев
- 33. Kumimanu biceae Высота 170 см. Вес 100 кг. Примобукко Размером со скворца. Имела округлое тело и
- 34. Palaeotis weigelti Имела длинные ноги. Имела маленькие крылья, не предназначенные для полетов. Была высотой до 1
- 35. Сумчатый саблезубый тигр Достигал размеров ягуара, тело длинное, ноги мощные и довольно короткие, полустопоходящие. Шея мощная,
- 36. Наиболее известным представителем диноцерат является уинтатерий Диноцераты в переводе означает «ужасные рога» и имеет в виду
- 37. Аминодонты Размером и телосложением они напоминали гиппопотама. Их конечности были короткими и мощными (на передних по
- 38. Пантолямбда По размеру был сопоставим с овцой, внешне напоминал животных семейства кошачьих, однако был травоядным, а
- 39. Разница в анатомии между гиракотериями и орогиппусами относительно небольшая. Они были приблизительно равны по размерам, 24—45
- 40. Тениодонты довольно быстро эволюционировали в узкоспециализированную группу роющих животных, однако внутри группы наблюдалась значительная вариация размеров
- 41. С изменением климатических условий менялись и растения палеогена. В палеогеновом периоде подавляющее количество растительных форм стало
- 42. На рубежах аридных поясов климат становился влажнее, что способствовало разрастанию растительности смешанных типов. По берегам водоемов
- 44. Скачать презентацию

Палеоге́новый пери́од— геологический период; первый период кайнозоя. Начался 66,0 млн лет
Палеоге́новый пери́од— геологический период; первый период кайнозоя. Начался 66,0 млн лет

В начале палеогена в Северном полушарии выделяются два крупных материка, соединявшихся

В течение палеогенового периода образования располагались в континентальных условиях. Исключением были
В течение палеогенового периода образования располагались в континентальных условиях. Исключением были

Она находилась несколько в иных условиях, чем Восточно-Европейская. На протяжении кайнозойской
Она находилась несколько в иных условиях, чем Восточно-Европейская. На протяжении кайнозойской

Палеоклиматы суши палеогена были разнообразны
Климатические особенности
Рисунок - Схематическая глобальная палеоклиматическая карта
Палеоклиматы суши палеогена были разнообразны
Климатические особенности
Рисунок - Схематическая глобальная палеоклиматическая карта

Начало кайнозойской эры связано с великим альпийским горообразованием (все высочайшие горные
Начало кайнозойской эры связано с великим альпийским горообразованием (все высочайшие горные

Необходимо акцентировать внимание на новых качествах, появившиеся у млекопитающих, которые могут
Необходимо акцентировать внимание на новых качествах, появившиеся у млекопитающих, которые могут

Морская фауна
Морская фауна

Моллюски в палеогене
Моллюски в палеогене

Брюхоногие моллюски
Брюхоногие – самая многообразная и распространенная группа моллюсков.
Брюхоногих около
Брюхоногие моллюски
Брюхоногие – самая многообразная и распространенная группа моллюсков.
Брюхоногих около


Костистые рыбы
Костистые рыбы

3 подотрядах китообразных
Древние киты - Archeoceti;
Усатые киты - Mysticeti;
Зубатые киты - Odontoceti.
3 подотрядах китообразных
Древние киты - Archeoceti;
Усатые киты - Mysticeti;
Зубатые киты - Odontoceti.

Древние киты - Archeoceti
Древние киты - Archeoceti

Усатые киты - Mysticeti
Усатые киты - Mysticeti

Зубатые киты - Odontoceti
Зубатые киты - Odontoceti

Акулы в палеогене
Акулы в палеогене

Мегалодон (лат. Carcharocles megalodon или Otodus megalodon, от греч. μέγας и
Мегалодон (лат. Carcharocles megalodon или Otodus megalodon, от греч. μέγας и
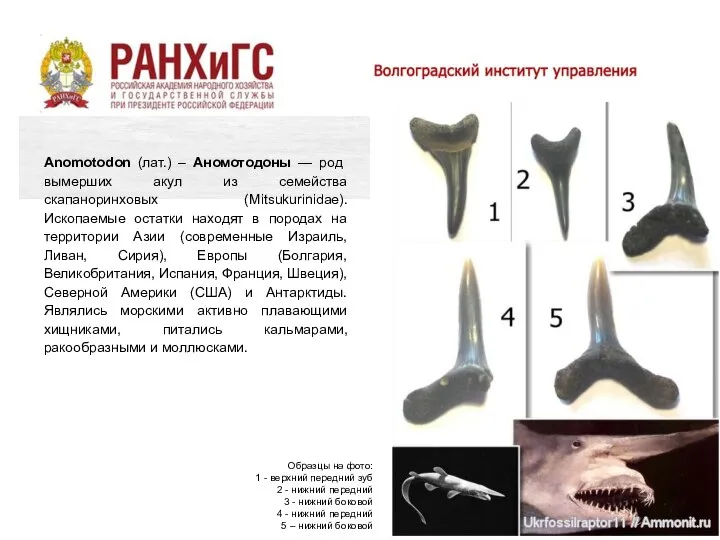
Anomotodon (лат.) – Аномотодоны — род вымерших акул из семейства скапаноринховых
Anomotodon (лат.) – Аномотодоны — род вымерших акул из семейства скапаноринховых

Кретоксирина (Cretoxyrhina) принадлежит к вымершему семейству Cretoxyrhinidae, входящему в современный отряд
Кретоксирина (Cretoxyrhina) принадлежит к вымершему семейству Cretoxyrhinidae, входящему в современный отряд
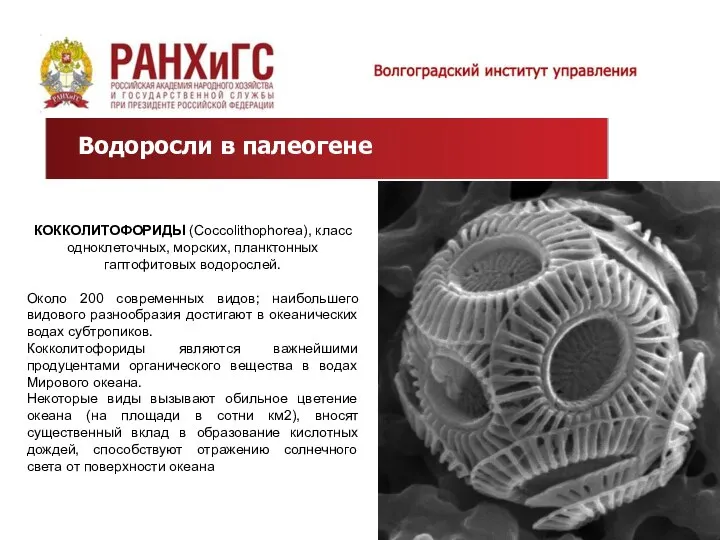
Водоросли в палеогене
КОККОЛИТОФОРИДЫ (Coccolithophorea), класс одноклеточных, морских, планктонных гаптофитовых
Водоросли в палеогене
КОККОЛИТОФОРИДЫ (Coccolithophorea), класс одноклеточных, морских, планктонных гаптофитовых
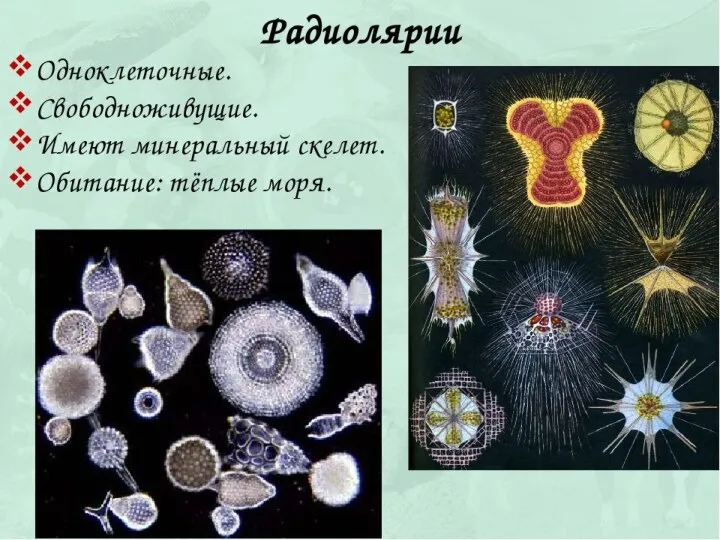
Cretoxyrhinidae (лат.) — семейство вымерших акул отряда ламнообразных, живших во время
Cretoxyrhinidae (лат.) — семейство вымерших акул отряда ламнообразных, живших во время

Кораллы в палеогене
СКЛЕРАКТИ́НИИ, мадрепоровые кораллы (Scleractinia, Madreporaria), отряд морских беспозвоночных
Кораллы в палеогене
СКЛЕРАКТИ́НИИ, мадрепоровые кораллы (Scleractinia, Madreporaria), отряд морских беспозвоночных


Высота птицы 2,5 метра.
Голова достигала 40 см в длину.
Имелся массивный загнутый
Высота птицы 2,5 метра.
Голова достигала 40 см в длину.
Имелся массивный загнутый

Достигали в высоту 1,75 метра.
Весили 100 кг.
Имели массивный клюв.
Длинные ноги,
Достигали в высоту 1,75 метра.
Весили 100 кг.
Имели массивный клюв.
Длинные ноги,

Высота птицы 1-3 метра.
Были хищниками.
На конце рудиментарных крыльев имели крюки.
Имели мощный
Высота птицы 1-3 метра.
Были хищниками.
На конце рудиментарных крыльев имели крюки.
Имели мощный

Имели длинные ноги и шею.
Строение черепа и клюва у них было
Имели длинные ноги и шею.
Строение черепа и клюва у них было

Oligocolius psittacocephalon.
Череп птицы был похож на череп попугая.
Предположительно был зоб.
Pelagornis sandersi
Размах
Oligocolius psittacocephalon.
Череп птицы был похож на череп попугая.
Предположительно был зоб.
Pelagornis sandersi
Размах

Kumimanu biceae
Высота 170 см.
Вес 100 кг.
Примобукко
Размером со скворца.
Имела округлое тело
Kumimanu biceae
Высота 170 см.
Вес 100 кг.
Примобукко
Размером со скворца.
Имела округлое тело

Palaeotis weigelti
Имела длинные ноги.
Имела маленькие крылья, не предназначенные для полетов.
Была высотой
Palaeotis weigelti
Имела длинные ноги.
Имела маленькие крылья, не предназначенные для полетов.
Была высотой

Сумчатый саблезубый тигр
Достигал размеров ягуара, тело длинное, ноги мощные
и довольно короткие,
Сумчатый саблезубый тигр
Достигал размеров ягуара, тело длинное, ноги мощные
и довольно короткие,

Наиболее известным представителем диноцерат является уинтатерий
Диноцераты в переводе означает «ужасные рога»
Наиболее известным представителем диноцерат является уинтатерий
Диноцераты в переводе означает «ужасные рога»

Аминодонты
Размером и телосложением они напоминали гиппопотама. Их конечности были короткими и
Аминодонты
Размером и телосложением они напоминали гиппопотама. Их конечности были короткими и

Пантолямбда
По размеру был сопоставим с овцой, внешне напоминал животных семейства кошачьих,
Пантолямбда
По размеру был сопоставим с овцой, внешне напоминал животных семейства кошачьих,

Разница в анатомии между гиракотериями и орогиппусами относительно небольшая. Они были
Разница в анатомии между гиракотериями и орогиппусами относительно небольшая. Они были

Тениодонты довольно быстро эволюционировали в узкоспециализированную группу роющих животных, однако внутри
Тениодонты довольно быстро эволюционировали в узкоспециализированную группу роющих животных, однако внутри

С изменением климатических условий менялись и растения палеогена. В палеогеновом периоде подавляющее
С изменением климатических условий менялись и растения палеогена. В палеогеновом периоде подавляющее

На рубежах аридных поясов климат становился влажнее, что способствовало разрастанию растительности
На рубежах аридных поясов климат становился влажнее, что способствовало разрастанию растительности
 Эпоха Великих географических открытий
Эпоха Великих географических открытий Кавказские Минеральные Воды - презентация к уроку Географии
Кавказские Минеральные Воды - презентация к уроку Географии Презентация на тему: Природная зона – тундра
Презентация на тему: Природная зона – тундра Природные зоны – арктическая пустыня
Природные зоны – арктическая пустыня Tourists & Homes
Tourists & Homes Украина. Общая характеристика страны - презентация к уроку Географии
Украина. Общая характеристика страны - презентация к уроку Географии 10 klase. Pasaules politiskā karte. Iedzīvotāji
10 klase. Pasaules politiskā karte. Iedzīvotāji 03.2020 Вулканизм
03.2020 Вулканизм Канада. Загальні відомості про країну
Канада. Загальні відомості про країну Географическая номенклатура. 8 класс
Географическая номенклатура. 8 класс Серые лесные, аллювиальные и болотные почвы
Серые лесные, аллювиальные и болотные почвы Россия - наша Родина
Россия - наша Родина Город Бремен (Германия)
Город Бремен (Германия) Индия
Индия Практическая работа «Определение расстояний, направлений и географических координат» составила: Кудрявцева Н.В. Учитель географ
Практическая работа «Определение расстояний, направлений и географических координат» составила: Кудрявцева Н.В. Учитель географ Пособие гражданам, имеющим детей
Пособие гражданам, имеющим детей NA ESIA Updates AZE_updated
NA ESIA Updates AZE_updated Развитие форм рельефа 8 класс
Развитие форм рельефа 8 класс Процессы урбанизации в современном мире
Процессы урбанизации в современном мире Викторина по географиии
Викторина по географиии Тектоника литосферных плит
Тектоника литосферных плит Презентация на тему Волга матушка
Презентация на тему Волга матушка Полезные ископаемые
Полезные ископаемые Сертификат Турция
Сертификат Турция Албания С чёрным орлом
Албания С чёрным орлом Рельеф Земли. Горы и равнины
Рельеф Земли. Горы и равнины Ледники. Современное наземное оледенение земли
Ледники. Современное наземное оледенение земли Мировой океан
Мировой океан