- Биоразнообразие. Biodiversity

Содержание
- 2. КАК ИЗМЕРЯТЬ ВИДОВОЕ РАЗНООБРАЗИЕ (БИОРАЗНООБРАЗИЕ BIODIVERSITY) Видовое богатство (число видов без случайных и временных вселенцев) Гетерогенность
- 3. Широтные градиенты биоразнообразия 620 видов деревьев
- 4. ШИРОТНЫЕ ГРАДИЕНТЫ
- 5. Какие факторы могут определять градиенты биоразнообразия Время в эволюционном масштабе То есть в старых сообщества больше
- 6. ЭВОЛЮЦИОННЫЙ МАСШТАБ ВРЕМЕНИ (МИЛЛИОНЫ ЛЕТ)
- 7. Гипотетическая схема повышения биоразнообразия со временем. Проверка истории, как фактора трудна. поэтому сначала на- до отбросить
- 8. Пространственная гетерогенность МакАртур (1965) ввел понятия: Внутрибиотопическое (альфа) р-зие и Межбиотопическое (бета) р-зие Вариант А в
- 9. Простая гипотеза разнообразие животных зависит от разнообразия деревьев, проходит только для амфибий
- 10. Увеличение биоразнообразия в тропиках показано и для водных экосистем, где нет гетерогенности. Таким образом, увеличение пространственной
- 11. Конкуренция Предполагают: Конкуренция играет большую роль в тропиках, тогда как в высоких широтах численность ограничена физическими
- 12. Хищничество Предполагают: Хищники держат популяции жертв на таком низком уровне, что они не конкурируют. Это позволяет
- 13. Модель Янсена-Конелла (Janzen-Connel) о роли хищничества. Конкуренция поддерживает разнообразие хищников и паразитов, а хищничество продуцентов и
- 14. ХИЩНИКИ
- 15. Климат и его флуктуации Более стабильный и мягкий климат – больше видовое богатство (Sanders, 1968). Сообщества
- 16. Видовое богатство для деревьев и позвоночных С. Америки и его связь с доступной энергией (Brown, 1981)
- 17. Продуктивность Гипотеза, чем выше продуктивность сообщества, тем выше разнообразие не проходит. Самые богатые растительные сообщества не
- 18. ОБИЛИЕ РЕСУРСОВ (ПРОДУКТИВНОСТЬ)
- 19. Модификация гипотезы влияния продуктивности. Более длительный сезон вегетации – более дробное временное деление ресурсов. Комбинация стабильности
- 20. ЧИСЛО ЗВЕНЬЕВ В ЦЕПЯХ ПИТАНИЯ x – число трофических уровней y – число сообществ
- 21. Нарушения В равновесии доминируют самые конкурентные. Хищничество, пастьба, изменения погоды, катастрофы (пожары) нарушают равновесие. Часто плохо
- 22. Оценка роли нарушений зависит от пространственной шкалы. Для небольших S – разнообразие стадий сукцессии. Но на
- 23. Гипотеза «среднего уровня нарушений» работает в одних биотопах (а) и не работает в других (b)
- 24. Примеры сообществ, где гипотеза не работает Донные с-ва ручьев Южного о-ва Новой Зеландии (температура, течение, дно)
- 25. Локальное и региональное разнообразие Локальное разнообразие определяется шкалой при которой виды сообщества могут взаимодействовать. Шкала регионального
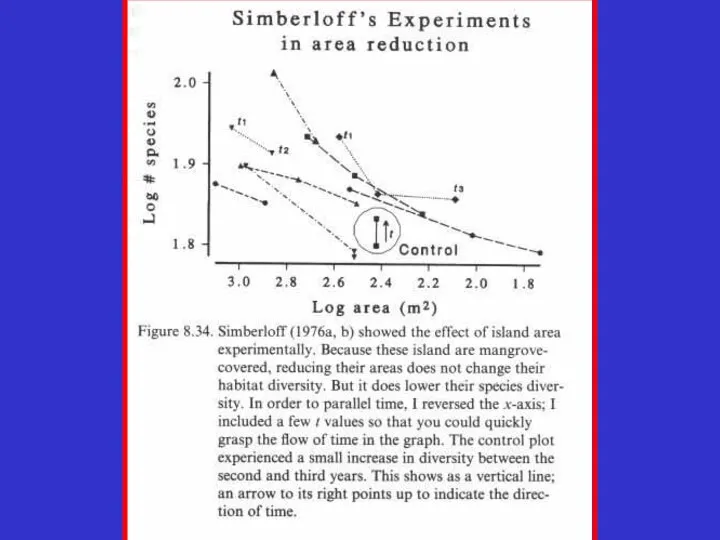
- 26. СВЯЗЬ ЧИСЛА ВИДОВ С ПЛОЩАДЬЮ ИЗОЛЯТА S=CAz, где S число видов, С -постоянная для данного таксона
- 27. S=CAz, где S число видов, С -постоянная для данного таксона и местности, А - площадь изолята,
- 31. БОГАТСТВО ОСТРОВНОЙ ФАУНЫ (MacArthur, Wilson, 1963)
- 32. Гипотетическое соотношение локального и регионального видового богатства в зависимости от насыщенности сообщества
- 33. Экспериментальная проверка 1. Птицы 15 га – 200-700 , ч. набл. (Pearson, 1977) 2. Разные сообщества
- 35. ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯ ШЕННОНА-УИВЕРА H=-Σ PilogPi
- 38. John Terborg, 1973. Тропики богаче видами потому что их территории больше
- 39. ЭКОЛОГИЧЕСКИЙ МАСШТАБ ВРЕМЕНИ
- 40. УСТОЙЧИВОСТЬ СРЕДЫ
- 41. ГЕТЕРОГЕННОСТЬ СРЕДЫ
- 42. КОНКУРЕНЦИЯ
- 43. ГИПОТЕЗЫ, ОБЪЯСНЯЮЩИЕ РАЗНООБРАЗИЕ ВИДОВ В СООБЩСТВАХ 1. РАЗМЕР ТЕРРИТОРИИ 3. ВРЕМЯ. ЭВОЛЮЦИОННЫЙ МАСШТАБ 4. ВРЕМЯ. ЭКОЛОГИЧЕСКИЙ
- 45. ЭКОЛОГИЧЕСКИЙ МАСШТАБ ВРЕМЕНИ (ГОДЫ) Динамика структуры сообщества грызунов Черных земель, Калмыкия
- 46. КОНКУРЕНЦИЯ
- 49. Скачать презентацию

КАК ИЗМЕРЯТЬ ВИДОВОЕ РАЗНООБРАЗИЕ (БИОРАЗНООБРАЗИЕ BIODIVERSITY)
Видовое богатство (число видов без случайных
КАК ИЗМЕРЯТЬ ВИДОВОЕ РАЗНООБРАЗИЕ (БИОРАЗНООБРАЗИЕ BIODIVERSITY)
Видовое богатство (число видов без случайных
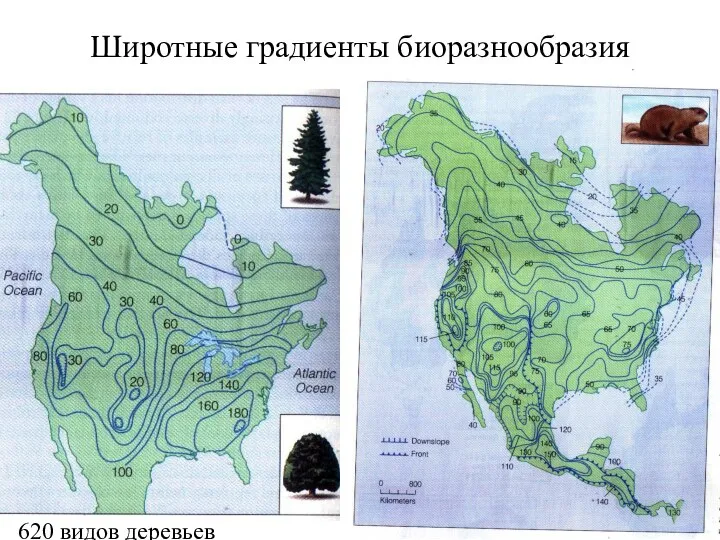
Широтные градиенты биоразнообразия
620 видов деревьев
Широтные градиенты биоразнообразия
620 видов деревьев
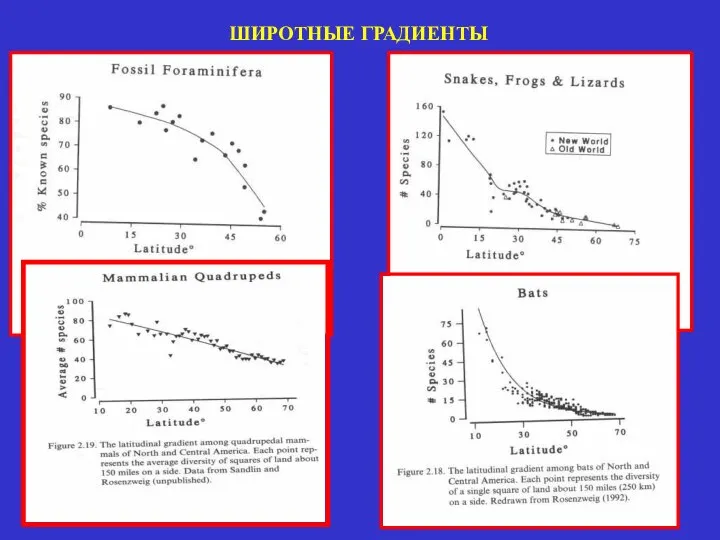
ШИРОТНЫЕ ГРАДИЕНТЫ
ШИРОТНЫЕ ГРАДИЕНТЫ

Какие факторы могут определять градиенты биоразнообразия
Время в эволюционном масштабе
То есть в
Какие факторы могут определять градиенты биоразнообразия
Время в эволюционном масштабе
То есть в
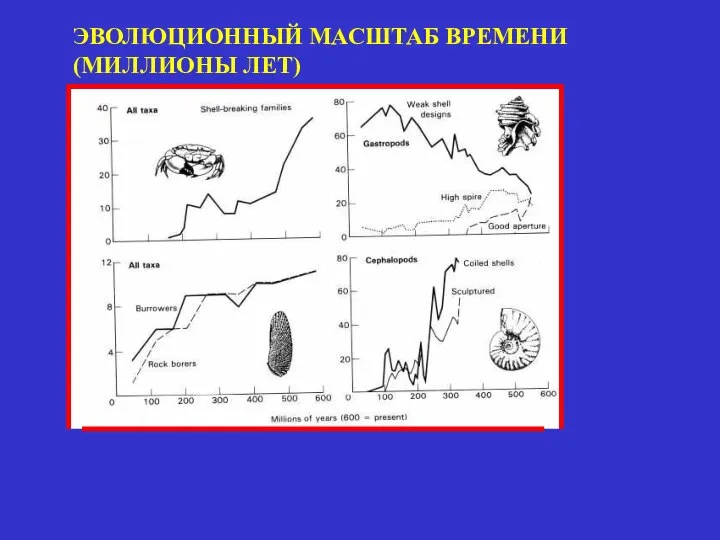
ЭВОЛЮЦИОННЫЙ МАСШТАБ ВРЕМЕНИ
(МИЛЛИОНЫ ЛЕТ)
ЭВОЛЮЦИОННЫЙ МАСШТАБ ВРЕМЕНИ
(МИЛЛИОНЫ ЛЕТ)
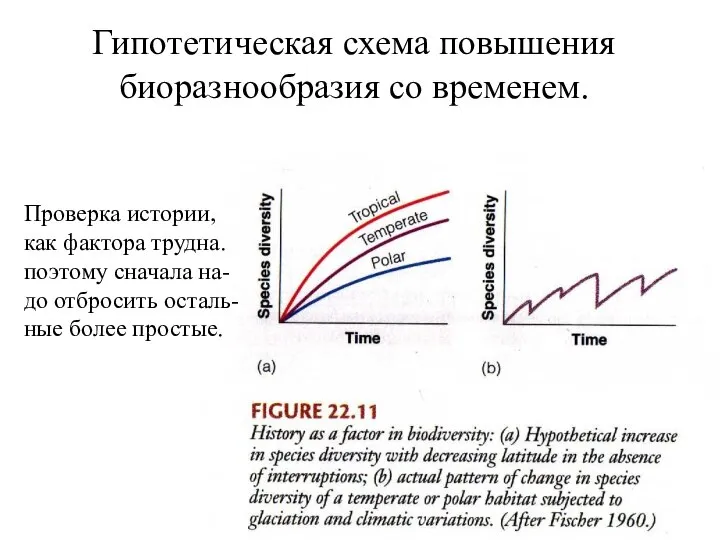
Гипотетическая схема повышения биоразнообразия со временем.
Проверка истории,
как фактора трудна.
поэтому сначала на-
до
Гипотетическая схема повышения биоразнообразия со временем.
Проверка истории,
как фактора трудна.
поэтому сначала на-
до

Пространственная гетерогенность
МакАртур (1965) ввел понятия: Внутрибиотопическое (альфа) р-зие и Межбиотопическое (бета)
Пространственная гетерогенность
МакАртур (1965) ввел понятия: Внутрибиотопическое (альфа) р-зие и Межбиотопическое (бета)
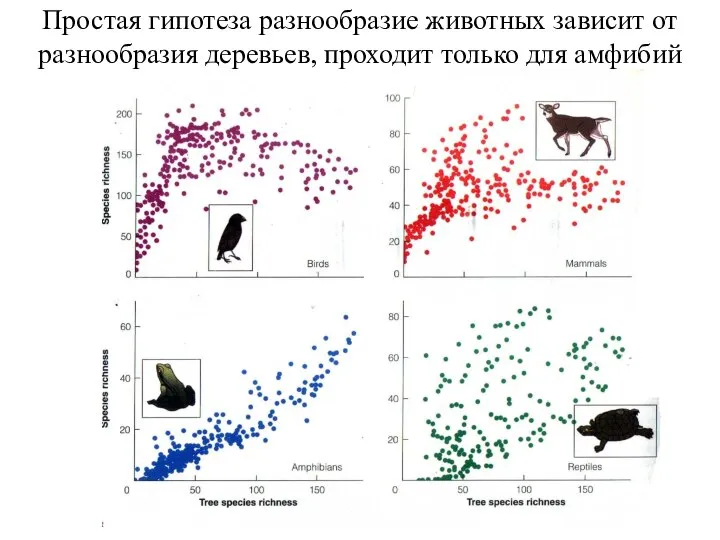
Простая гипотеза разнообразие животных зависит от разнообразия деревьев, проходит только для
Простая гипотеза разнообразие животных зависит от разнообразия деревьев, проходит только для

Увеличение биоразнообразия в тропиках показано и для водных экосистем, где нет
Увеличение биоразнообразия в тропиках показано и для водных экосистем, где нет

Конкуренция
Предполагают: Конкуренция играет большую роль в тропиках, тогда как в высоких
Конкуренция
Предполагают: Конкуренция играет большую роль в тропиках, тогда как в высоких

Хищничество
Предполагают: Хищники держат популяции жертв на таком низком уровне, что они
Хищничество
Предполагают: Хищники держат популяции жертв на таком низком уровне, что они

Модель Янсена-Конелла
(Janzen-Connel) о роли
хищничества.
Конкуренция поддерживает
разнообразие хищников и
Модель Янсена-Конелла
(Janzen-Connel) о роли
хищничества.
Конкуренция поддерживает
разнообразие хищников и
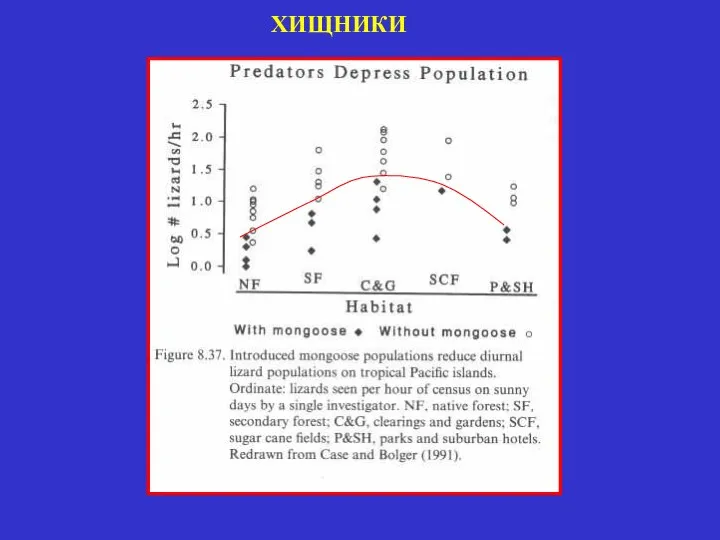
ХИЩНИКИ
ХИЩНИКИ

Климат и его флуктуации
Более стабильный и мягкий климат – больше видовое
Климат и его флуктуации
Более стабильный и мягкий климат – больше видовое
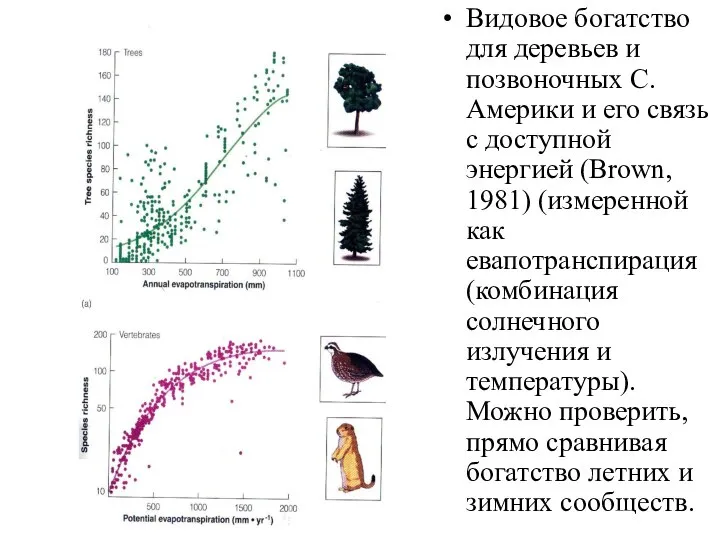
Видовое богатство для деревьев и позвоночных С. Америки и его связь
Видовое богатство для деревьев и позвоночных С. Америки и его связь

Продуктивность
Гипотеза, чем выше продуктивность сообщества, тем выше разнообразие не проходит.
Самые богатые
Продуктивность
Гипотеза, чем выше продуктивность сообщества, тем выше разнообразие не проходит.
Самые богатые
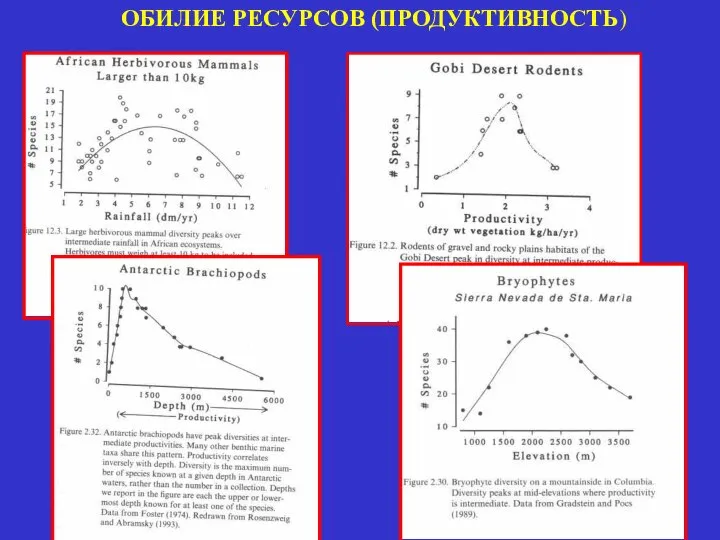
ОБИЛИЕ РЕСУРСОВ (ПРОДУКТИВНОСТЬ)
ОБИЛИЕ РЕСУРСОВ (ПРОДУКТИВНОСТЬ)

Модификация гипотезы влияния продуктивности.
Более длительный сезон вегетации – более дробное временное
Модификация гипотезы влияния продуктивности.
Более длительный сезон вегетации – более дробное временное
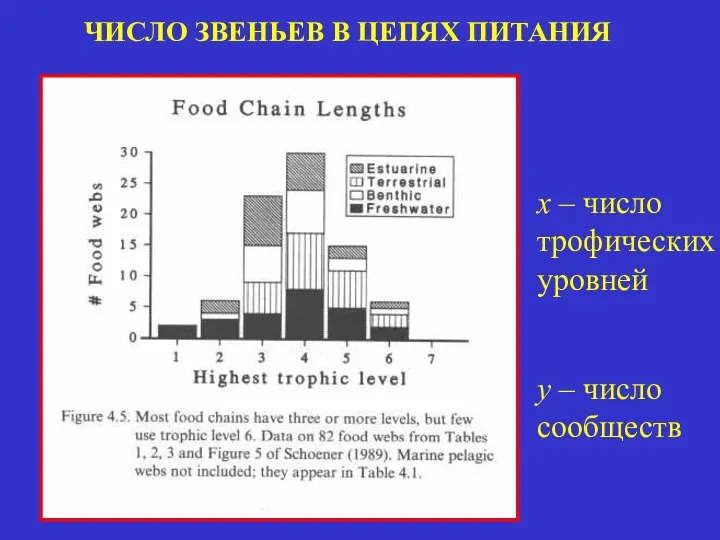
ЧИСЛО ЗВЕНЬЕВ В ЦЕПЯХ ПИТАНИЯ
x – число трофических уровней
y – число
ЧИСЛО ЗВЕНЬЕВ В ЦЕПЯХ ПИТАНИЯ
x – число трофических уровней
y – число

Нарушения
В равновесии доминируют самые конкурентные.
Хищничество, пастьба, изменения погоды, катастрофы (пожары) нарушают
Нарушения
В равновесии доминируют самые конкурентные.
Хищничество, пастьба, изменения погоды, катастрофы (пожары) нарушают

Оценка роли нарушений зависит от пространственной шкалы. Для небольших S –
Оценка роли нарушений зависит от пространственной шкалы. Для небольших S –
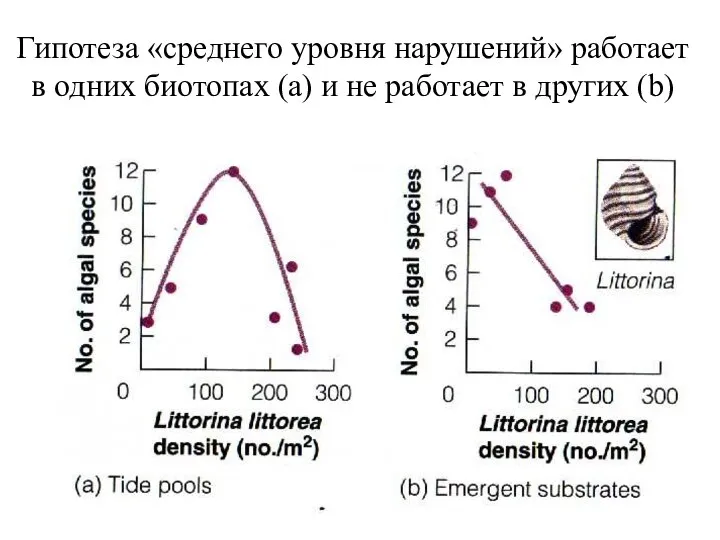
Гипотеза «среднего уровня нарушений» работает в одних биотопах (а) и не
Гипотеза «среднего уровня нарушений» работает в одних биотопах (а) и не
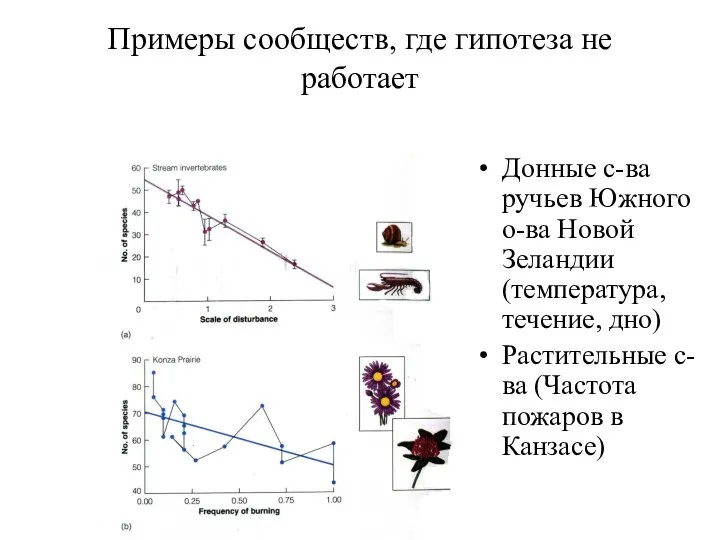
Примеры сообществ, где гипотеза не работает
Донные с-ва ручьев Южного о-ва Новой
Примеры сообществ, где гипотеза не работает
Донные с-ва ручьев Южного о-ва Новой

Локальное и региональное разнообразие
Локальное разнообразие определяется шкалой при которой виды сообщества
Локальное и региональное разнообразие
Локальное разнообразие определяется шкалой при которой виды сообщества

СВЯЗЬ ЧИСЛА ВИДОВ С ПЛОЩАДЬЮ ИЗОЛЯТА
S=CAz,
где S число
СВЯЗЬ ЧИСЛА ВИДОВ С ПЛОЩАДЬЮ ИЗОЛЯТА
S=CAz,
где S число
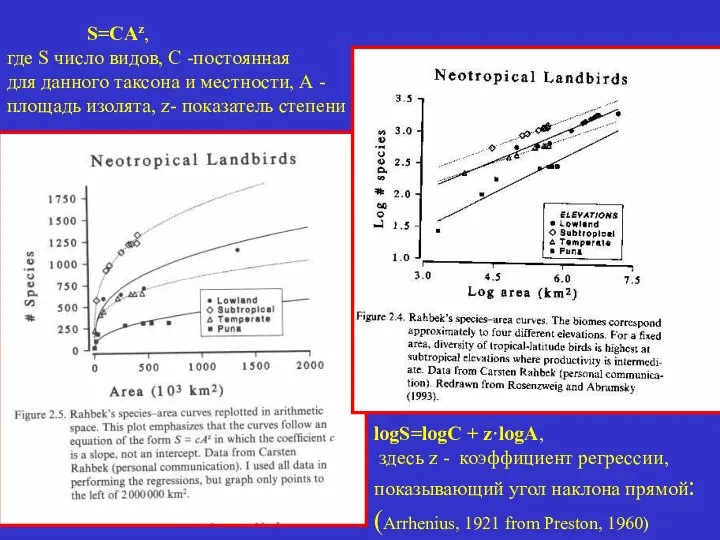
S=CAz,
где S число видов, С -постоянная
для данного таксона и
S=CAz,
где S число видов, С -постоянная
для данного таксона и


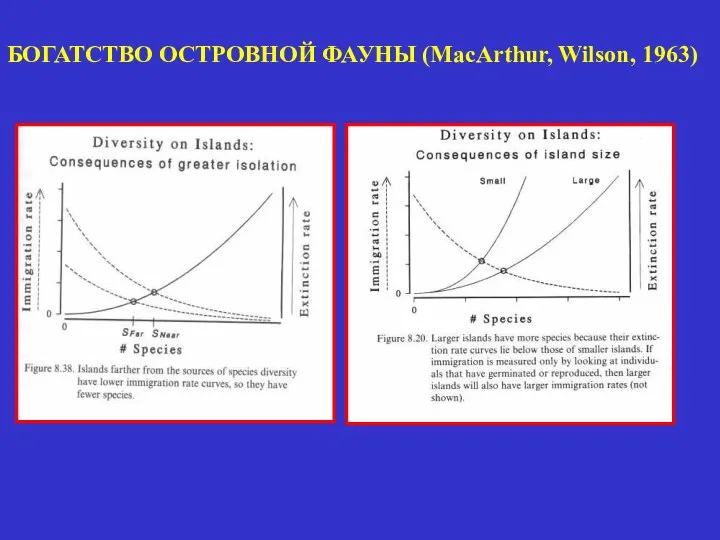
БОГАТСТВО ОСТРОВНОЙ ФАУНЫ (MacArthur, Wilson, 1963)
БОГАТСТВО ОСТРОВНОЙ ФАУНЫ (MacArthur, Wilson, 1963)
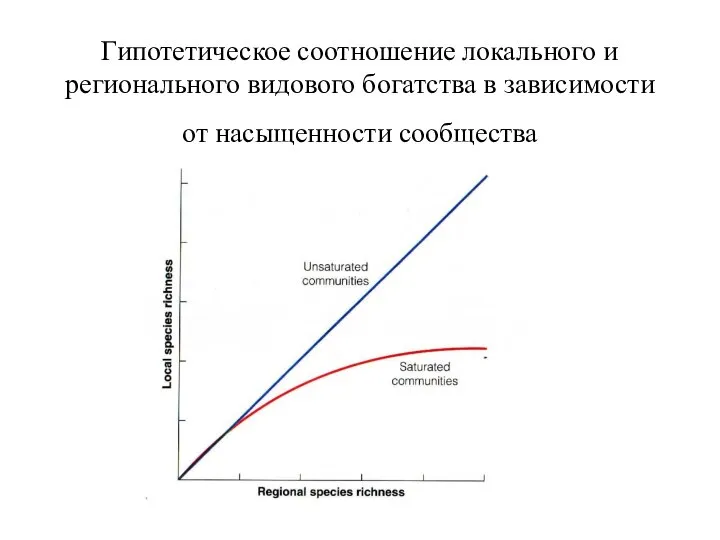
Гипотетическое соотношение локального и регионального видового богатства в зависимости от насыщенности
Гипотетическое соотношение локального и регионального видового богатства в зависимости от насыщенности
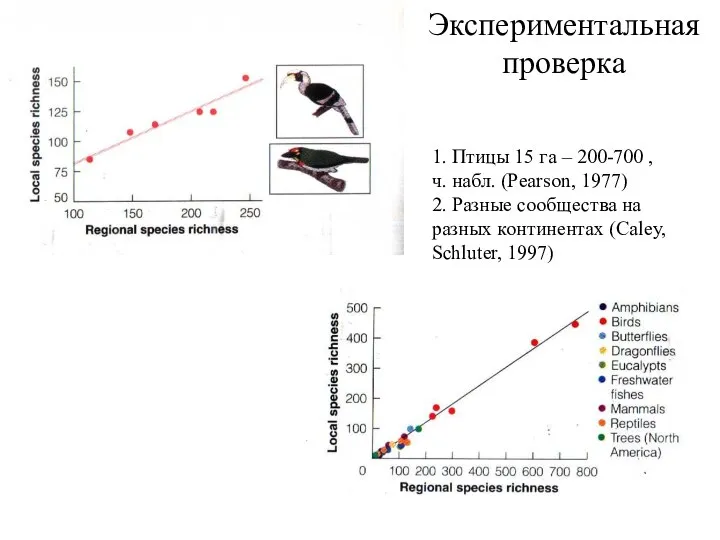
Экспериментальная проверка
1. Птицы 15 га – 200-700 , ч. набл. (Pearson,
Экспериментальная проверка
1. Птицы 15 га – 200-700 , ч. набл. (Pearson,


ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯ
ШЕННОНА-УИВЕРА
H=-Σ PilogPi
ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯ
ШЕННОНА-УИВЕРА
H=-Σ PilogPi


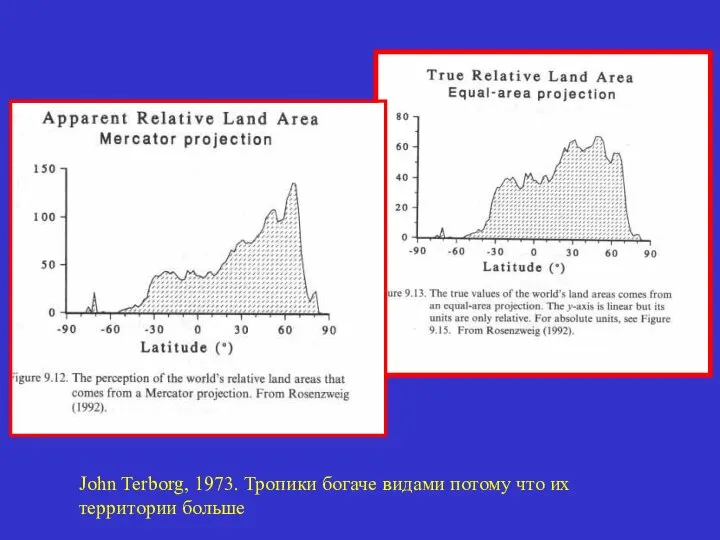
John Terborg, 1973. Тропики богаче видами потому что их
территории больше
John Terborg, 1973. Тропики богаче видами потому что их
территории больше

ЭКОЛОГИЧЕСКИЙ МАСШТАБ ВРЕМЕНИ
ЭКОЛОГИЧЕСКИЙ МАСШТАБ ВРЕМЕНИ
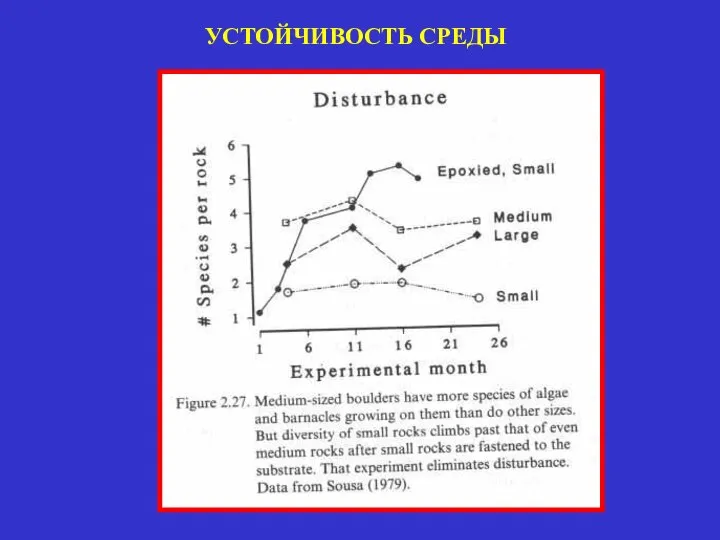
УСТОЙЧИВОСТЬ СРЕДЫ
УСТОЙЧИВОСТЬ СРЕДЫ
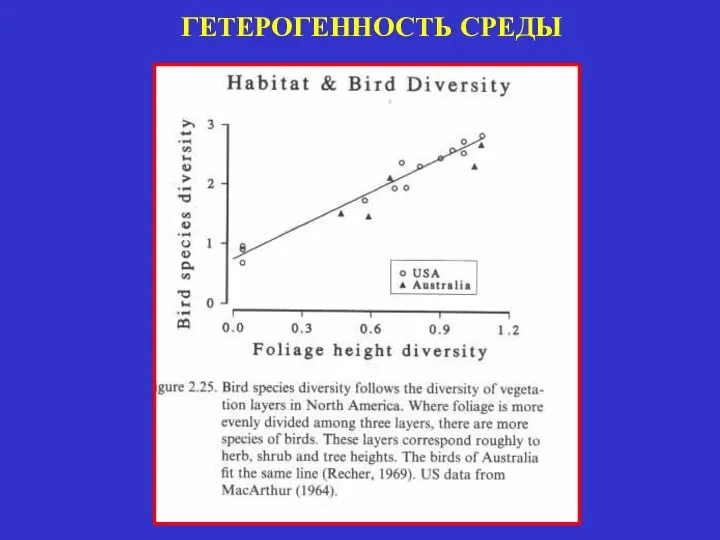
ГЕТЕРОГЕННОСТЬ СРЕДЫ
ГЕТЕРОГЕННОСТЬ СРЕДЫ

КОНКУРЕНЦИЯ
КОНКУРЕНЦИЯ

ГИПОТЕЗЫ, ОБЪЯСНЯЮЩИЕ РАЗНООБРАЗИЕ
ВИДОВ В СООБЩСТВАХ
1. РАЗМЕР ТЕРРИТОРИИ
3. ВРЕМЯ. ЭВОЛЮЦИОННЫЙ МАСШТАБ
4.
ГИПОТЕЗЫ, ОБЪЯСНЯЮЩИЕ РАЗНООБРАЗИЕ
ВИДОВ В СООБЩСТВАХ
1. РАЗМЕР ТЕРРИТОРИИ
3. ВРЕМЯ. ЭВОЛЮЦИОННЫЙ МАСШТАБ
4.


ЭКОЛОГИЧЕСКИЙ МАСШТАБ
ВРЕМЕНИ (ГОДЫ)
Динамика структуры сообщества
грызунов Черных земель, Калмыкия
ЭКОЛОГИЧЕСКИЙ МАСШТАБ
ВРЕМЕНИ (ГОДЫ)
Динамика структуры сообщества
грызунов Черных земель, Калмыкия

КОНКУРЕНЦИЯ
КОНКУРЕНЦИЯ

 How are you feeling?
How are you feeling? Гидросфера. Экологические проблемы
Гидросфера. Экологические проблемы Загрязнение пресных вод
Загрязнение пресных вод Альтернативная энергетика
Альтернативная энергетика Экология атмосферы
Экология атмосферы Рациональное природопользование и охрана окружающей среды
Рациональное природопользование и охрана окружающей среды Шымкент қаласының экологиясы
Шымкент қаласының экологиясы Мой дом – экологически безопасное жилище
Мой дом – экологически безопасное жилище Утилизация отходов
Утилизация отходов Определение запыленности воздуха по снегу
Определение запыленности воздуха по снегу Природоохранные лимиты
Природоохранные лимиты Мировая проблема окончания ресурсов
Мировая проблема окончания ресурсов Понятие экологического аудита
Понятие экологического аудита Презентация Рельеф России
Презентация Рельеф России Здровье человека и современная окружающая среда
Здровье человека и современная окружающая среда Влияние жилищных условий на здоровье населения. Гигиенические и экологические проблемы. Застройки населенных пунктов
Влияние жилищных условий на здоровье населения. Гигиенические и экологические проблемы. Застройки населенных пунктов Нормирование выбросов на АО «НИИ Атмосфера». Законодательные, нормативные и методические документы
Нормирование выбросов на АО «НИИ Атмосфера». Законодательные, нормативные и методические документы Загрязнение Мирового океана
Загрязнение Мирового океана Аттестационная работа. Моя экологическая грамотность
Аттестационная работа. Моя экологическая грамотность Ағызынды сулардың экологиялық жағдайы
Ағызынды сулардың экологиялық жағдайы Красная и чёрная книга
Красная и чёрная книга Зачем нам нужен лес?
Зачем нам нужен лес? Мусор на наших улицах
Мусор на наших улицах Ауыз судың химиялық тұрғыдан зиянсыздығын сипаттаушы көрсеткіштер
Ауыз судың химиялық тұрғыдан зиянсыздығын сипаттаушы көрсеткіштер Работа международных экологических организаций России
Работа международных экологических организаций России Экологически значимые свойства воздуха. Погода, климат, микроклимат. (Лекция 3)
Экологически значимые свойства воздуха. Погода, климат, микроклимат. (Лекция 3) Збереження довкілля
Збереження довкілля Проблемы реки Ушайка, Томской области
Проблемы реки Ушайка, Томской области