- Физиологическая (функциональная) экология

Содержание
- 2. Основатели: Джордж Бартоломью, Кнут Шмидт-Ниельсен, Пер Шоландер, Лоренс Ирвинг, Н.И. Калабухов, А.Д. Слоним, И.А. Шилов.
- 3. George A. Bartholomew (1919-2006) Knut Schmidt-Nielsen (1915–2007) Per Fredrik Scholander (1905-1980)
- 4. Абрам Донович Слоним (1903-1986) Игорь Александрович Шилов (1921-2001) Николай Иванович Калабухов (1908–1991)
- 5. В самостоятельную дисциплину функциональная экология выделилась в конце 80-х годов XX века. 1987 год: появление книги
- 6. Энергетика Расход энергии животным отражает цену его жизнедеятельности. Изучение энергетики на организменном, популяционном и видовом уровнях
- 7. Изменение веществ в процессе их биогенного круговорота происходит с затратой энергии Главный источник энергии – энергия
- 8. Биоэнергетика
- 9. 40-60 кДж/ моль 13,8 кДж/моль, АТФ - В клетке около 1 млрд молекул АТФ. - Каждая
- 10. БОЛЬШАЯ ЭНЕРГИЯ, GEI экскреторная энергия, EE метаболизированная энергия, ME энергия существования, EMR продуктивная энергия, PE энергия
- 11. Усвоение энергии из пищи (MEC) MEC = ME/GEI, но GEI = ME + EE, MEC =
- 12. Несмотря на то, что MEC не может быть в строгом смысле характеристикой пищи, накопленные данные по
- 13. 20 15 10 5 Семена ложноцветных Семена крестоцветных Рыбы Личинки насекомых Звери Имаго насекомых Пауки Пресмыкающиеся
- 14. Состав пищи (жиры/белки/углеводы) Биоэнергетика
- 15. Питательное вещество Углевод Жир Белок (→мочевина) Белок (→мочевая кислота) (а) Ккал/г (б) л О2/г (в) ккал/л
- 16. Углеводы (глюкоза): C6H12O6 + 6O2 ??? 6CO2 + 6H2O + Energy RQ = 6CO2 / 6O2
- 17. БОЛЬШАЯ ЭНЕРГИЯ, GEI экскреторная энергия, EE метаболизированная энергия, ME энергия существования, EMR продуктивная энергия, PE энергия
- 18. Энергия, накопленная в тканях гетеротрофа = вторичная продукция экосистемы. НО энергия, доступная для потребления другими гетеротрофами
- 20. var P/ME > x10 var PE/ME
- 22. Организм - Та Диапазон Та воздуха: +50°С ………..-50°С = = Фундаментальная характеристика биосферы. С-В Евразия ………
- 23. Q10 = K t+10 / K t = 2-3 для большинства абиотических реакций. У многих животных
- 24. 10 30 20 Q10=3 Q10=2 Т°С Потребление О2 y=b ax Организм – Та. Q10 Скорость процесса
- 25. R2=R1 Q10 (Т2 - Т1)/10 Организм – Та. Q10 log y = log b + x
- 26. (Т2 - Т1)/10 log R 2 Организм – Та. Q10 y=b ax R2=R1 Q10 (Т2 -
- 27. 10 20 15 Т°С Log O2, мкл/г * ч Организм – Та. Q10 y=b ax R2=R1
- 28. Организм – Та. Ta Lim Температурные пороги жизни Изменчивость диапазона Та Изменение Та-толерантности в течение жизни:
- 29. Организм – Та. Ta Lim Верхний температурный предел Ни одно животное не может пройти весь жизненный
- 30. Организм – Та. Ta Lim Обезвоживание ? повышение устойчивости к Та (цисты, споры, семена): Личинки мухи
- 31. Т°С % выживших Организм – Та. Ta Lim LT50 100 50 Определение летальных температур Летальная Та
- 32. LT50 и причины гибели при перегреве Организм – Та. Ta Lim У многих сложно-организованных животных Та
- 33. Организм – Та. Ta Lim Причины тепловой смерти: Денатурация белков (45-55°С для ряда белков, но это
- 34. Организм – Та. Ta Lim Нижний температурный предел Рассогласование реакций, имеющих разные Q10 Инактивация ферментов (у
- 35. Организм – Та. Ta Lim Структурные изменения клеток и тканей 2 механизма у животных из холодных
- 36. Организм – Та. Ta Lim Переохлаждение При отсутствии чужеродных частиц чистую воду можно охладить до -20°С.
- 37. Организм – Та. Ta Lim АНТИФРИЗЫ А Б Глицерин – есть у многих насекомых. Защищает ткани
- 38. -1.0 -1.0 -1.5 -1.73° -1.73° -1.73° + 5.0° T°С Глубина, м Северный Лабрадор, Костистые рыбы С=400
- 39. Вещество со свойствами антифриза, выделенное из антарктической рыбы Trematomus, в водном растворе с необыкновенной эффективностью препятствует
- 40. Гликопротеид Trematomus (2 аминокислоты и дисахарид): АЛАНИН 23% ТРЕОНИН 16% ДИСАХАРИД Организм – Та. Ta Lim
- 41. Физиологические адаптации к перемене температуры Организм – Та. Ta Lim Воздействие сублетальной Та адаптация к летальной
- 42. Скорость акклимации Организм – Та. Ta Lim Другие факторы: возраст, размеры, качество воды, ФОТОПЕРИОД…
- 43. Сплошная линия очерчивает полный диапазон Та-толерантности золотой рыбки (Fry et al., 1942). У рыбки, содержавшейся при
- 44. Организм – Та. Ta Lim
- 45. Та-компенсация и интенсивность метаболизма. Правило компенсации Организм – Та. Ta Lim
- 46. Организм – Та. Ta Lim Варианты протекания процессов компенсации при изменении Та от Т1 до Т2
- 47. Организм – Та. Ta Lim Температурная устойчивость животных Генетические различия (~связанные с их географическим распространением) Сезонные
- 49. Research Summary last updated by G. A. Bartholomew in 1989 and slightly updated in 2006 by
- 50. Knut Schmidt-Nielsen Born in TrondheimBorn in Trondheim, NorwayBorn in Trondheim, Norway. He was educated in OsloBorn
- 51. Scholander, Per Fredrik (Thorkelsson) (Norway- United States 1905-1980) physiological ecology Physiologist Scholander became well known for
- 52. СЛОНИМ Абрам Донович (1903, Петровск-ПортСЛОНИМ Абрам Донович (1903, Петровск-Порт Дагестанской обл. – 1973, Фрунзе) физиолог. Д-р
- 54. Скачать презентацию

Основатели: Джордж Бартоломью, Кнут Шмидт-Ниельсен, Пер Шоландер, Лоренс Ирвинг, Н.И. Калабухов,
Основатели: Джордж Бартоломью, Кнут Шмидт-Ниельсен, Пер Шоландер, Лоренс Ирвинг, Н.И. Калабухов,

George A. Bartholomew
(1919-2006)
Knut Schmidt-Nielsen
(1915–2007)
Per Fredrik Scholander
(1905-1980)
George A. Bartholomew
(1919-2006)
Knut Schmidt-Nielsen
(1915–2007)
Per Fredrik Scholander
(1905-1980)

Абрам Донович Слоним
(1903-1986)
Игорь Александрович Шилов
(1921-2001)
Николай Иванович Калабухов
(1908–1991)
Абрам Донович Слоним
(1903-1986)
Игорь Александрович Шилов
(1921-2001)
Николай Иванович Калабухов
(1908–1991)

В самостоятельную дисциплину функциональная экология выделилась в конце 80-х годов
В самостоятельную дисциплину функциональная экология выделилась в конце 80-х годов

Энергетика
Расход энергии животным отражает цену его жизнедеятельности.
Изучение энергетики на
Энергетика
Расход энергии животным отражает цену его жизнедеятельности.
Изучение энергетики на
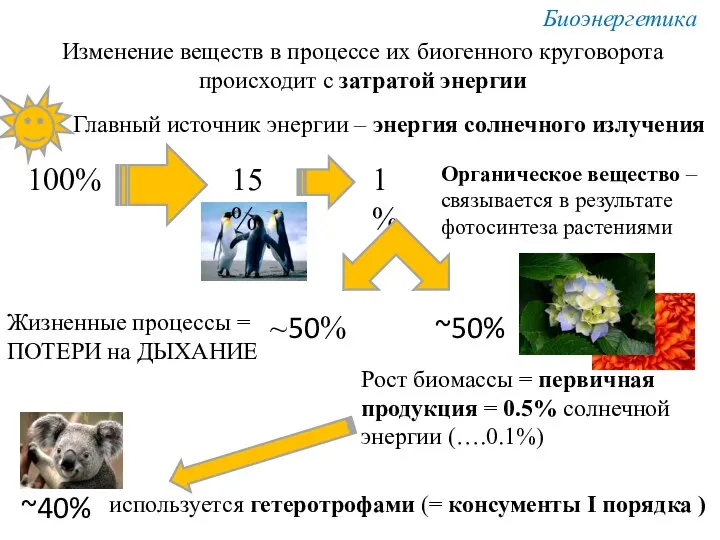
Изменение веществ в процессе их биогенного круговорота
происходит с затратой энергии
Главный
Изменение веществ в процессе их биогенного круговорота
происходит с затратой энергии
Главный
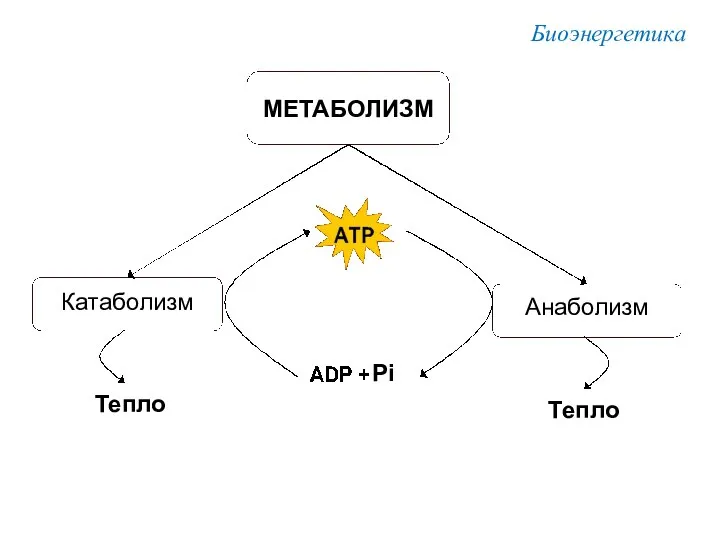
Биоэнергетика
Биоэнергетика
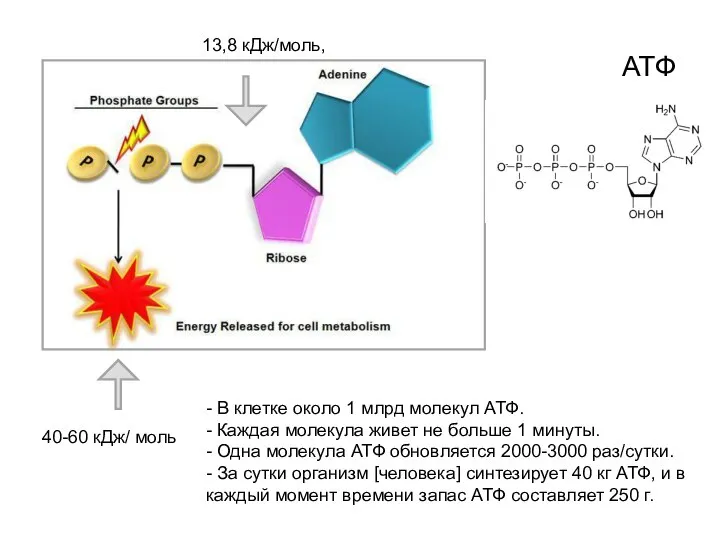
40-60 кДж/ моль
13,8 кДж/моль,
АТФ
- В клетке около 1 млрд молекул АТФ.
40-60 кДж/ моль
13,8 кДж/моль,
АТФ
- В клетке около 1 млрд молекул АТФ.
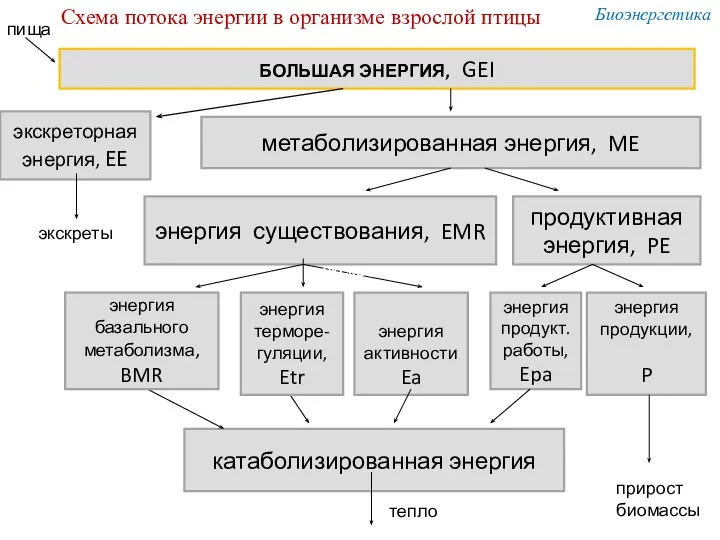
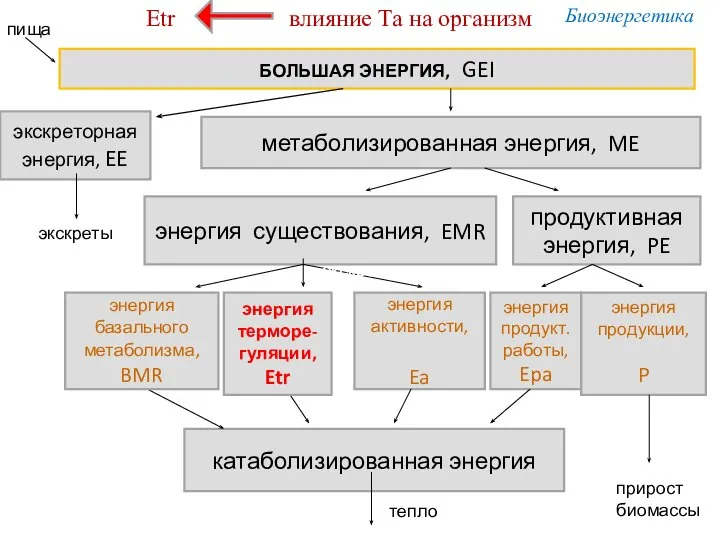
БОЛЬШАЯ ЭНЕРГИЯ, GEI
экскреторная
энергия, EE
метаболизированная энергия, ME
энергия существования, EMR
продуктивная энергия, PE
энергия
БОЛЬШАЯ ЭНЕРГИЯ, GEI
экскреторная
энергия, EE
метаболизированная энергия, ME
энергия существования, EMR
продуктивная энергия, PE
энергия

Усвоение энергии из пищи (MEC) < 100%
MEC = ME/GEI,
но GEI
Усвоение энергии из пищи (MEC) < 100%
MEC = ME/GEI,
но GEI

Несмотря на то, что MEC не может быть в строгом смысле
Несмотря на то, что MEC не может быть в строгом смысле

20
15
10
5
Семена ложноцветных
Семена крестоцветных
Рыбы
Личинки насекомых
Звери
Имаго насекомых
Пауки
Пресмыкающиеся
Земноводные
Почки цветковых
Хвоя
Разнотравье
Цветы
Семена хвойных
Листья
Ягоды
Пыльники хвойных
Черви
Моллюски
Семена злаков
Ракообразные
Семена березы
Яйца птиц
Яйца
20
15
10
5
Семена ложноцветных
Семена крестоцветных
Рыбы
Личинки насекомых
Звери
Имаго насекомых
Пауки
Пресмыкающиеся
Земноводные
Почки цветковых
Хвоя
Разнотравье
Цветы
Семена хвойных
Листья
Ягоды
Пыльники хвойных
Черви
Моллюски
Семена злаков
Ракообразные
Семена березы
Яйца птиц
Яйца
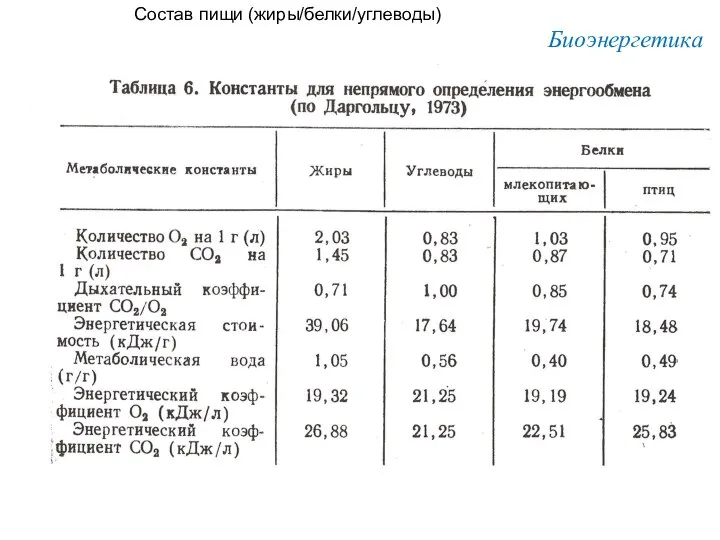
Состав пищи (жиры/белки/углеводы)
Биоэнергетика
Состав пищи (жиры/белки/углеводы)
Биоэнергетика
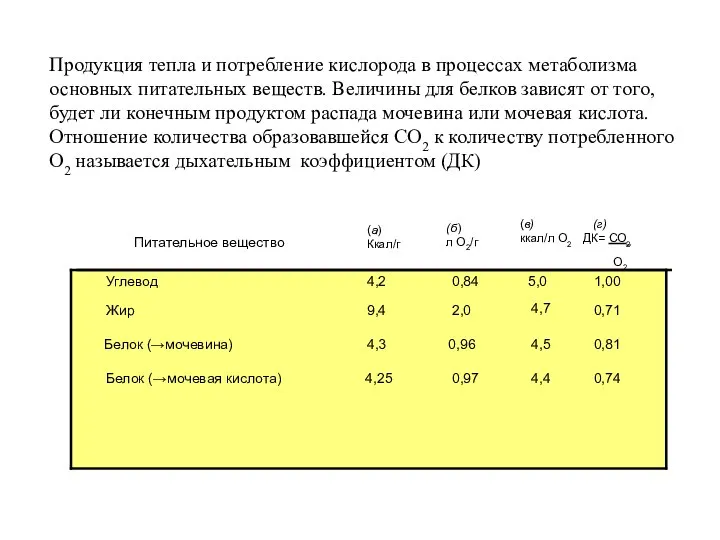
Питательное вещество
Углевод
Жир
Белок (→мочевина)
Белок (→мочевая кислота)
(а)
Ккал/г
(б)
л О2/г
(в)
ккал/л О2
(г)
ДК= СО2
О2
4,2
9,4
4,3
4,25
0,84
Питательное вещество
Углевод
Жир
Белок (→мочевина)
Белок (→мочевая кислота)
(а)
Ккал/г
(б)
л О2/г
(в)
ккал/л О2
(г)
ДК= СО2
О2
4,2
9,4
4,3
4,25
0,84
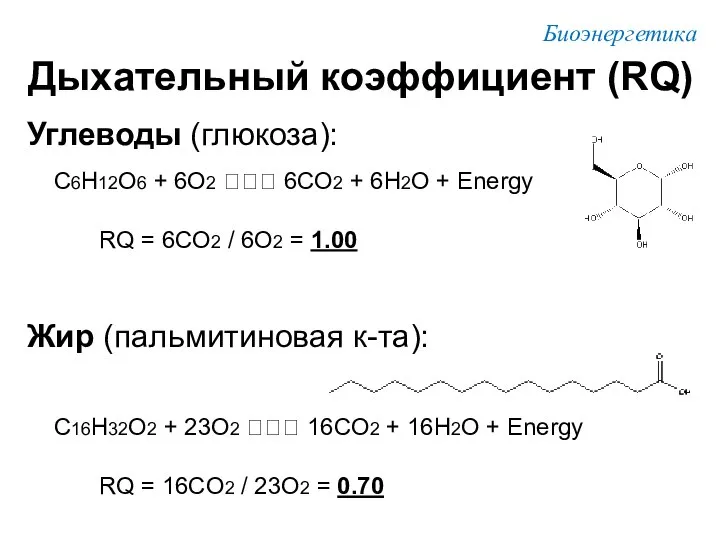
Углеводы (глюкоза):
C6H12O6 + 6O2 ??? 6CO2 + 6H2O + Energy
RQ =
Углеводы (глюкоза):
C6H12O6 + 6O2 ??? 6CO2 + 6H2O + Energy
RQ =
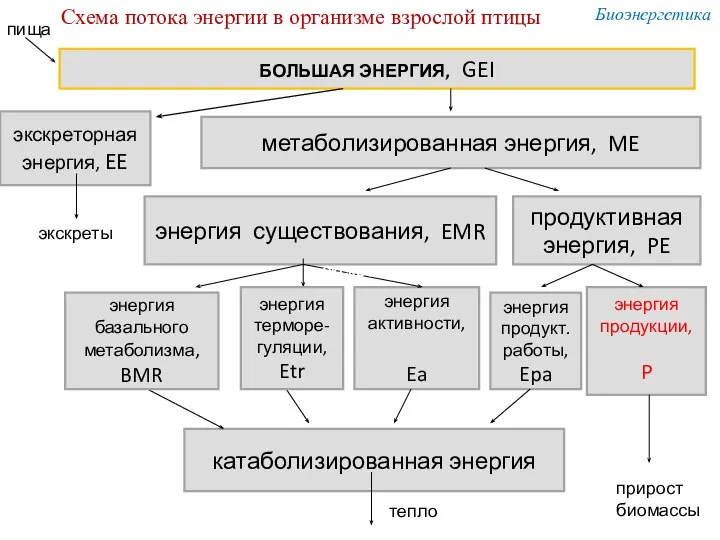
БОЛЬШАЯ ЭНЕРГИЯ, GEI
экскреторная
энергия, EE
метаболизированная энергия, ME
энергия существования, EMR
продуктивная энергия, PE
энергия
БОЛЬШАЯ ЭНЕРГИЯ, GEI
экскреторная
энергия, EE
метаболизированная энергия, ME
энергия существования, EMR
продуктивная энергия, PE
энергия

Энергия, накопленная в тканях гетеротрофа = вторичная
продукция экосистемы.
НО энергия, доступная
Энергия, накопленная в тканях гетеротрофа = вторичная
продукция экосистемы.
НО энергия, доступная


var P/ME > x10
var PE/ME <=x2
var P/ME > x10
var PE/ME <=x2

Организм - Та
Диапазон Та воздуха: +50°С ………..-50°С =
= Фундаментальная характеристика
Организм - Та
Диапазон Та воздуха: +50°С ………..-50°С =
= Фундаментальная характеристика

Q10 = K t+10 / K t = 2-3 для большинства
Q10 = K t+10 / K t = 2-3 для большинства
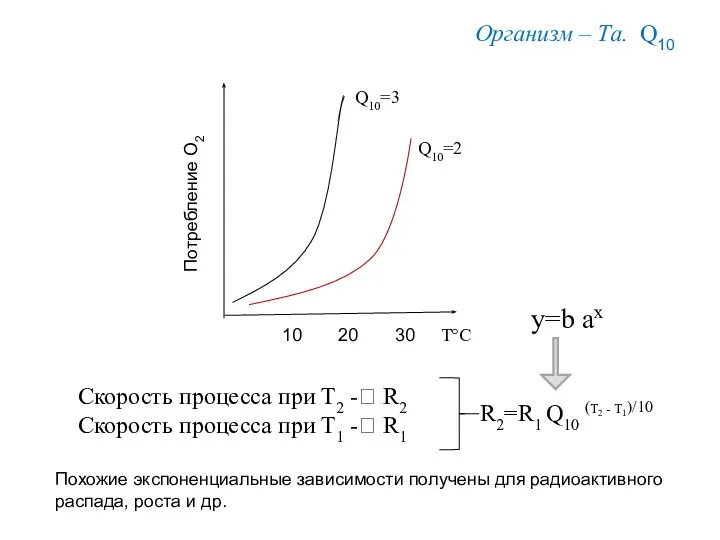
10
30
20
Q10=3
Q10=2
Т°С
Потребление О2
y=b ax
Организм – Та. Q10
Скорость процесса при Т2 -? R2
10
30
20
Q10=3
Q10=2
Т°С
Потребление О2
y=b ax
Организм – Та. Q10
Скорость процесса при Т2 -? R2

R2=R1 Q10 (Т2 - Т1)/10
Организм – Та. Q10
log y = log
R2=R1 Q10 (Т2 - Т1)/10
Организм – Та. Q10
log y = log
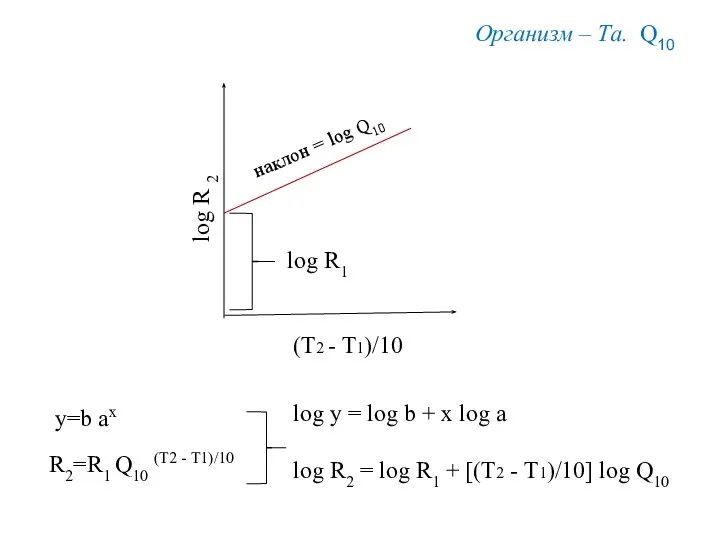
(Т2 - Т1)/10
log R 2
Организм – Та. Q10
y=b ax
R2=R1 Q10 (Т2
(Т2 - Т1)/10
log R 2
Организм – Та. Q10
y=b ax
R2=R1 Q10 (Т2
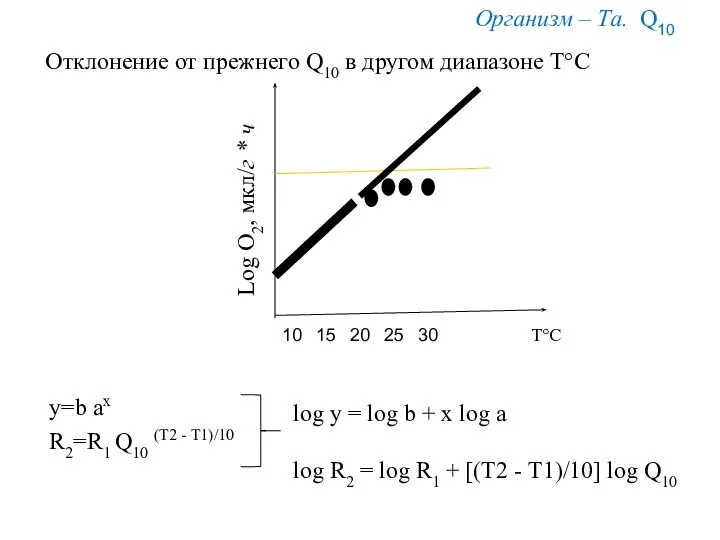
10
20
15
Т°С
Log O2, мкл/г * ч
Организм – Та. Q10
y=b ax
R2=R1 Q10 (Т2
10
20
15
Т°С
Log O2, мкл/г * ч
Организм – Та. Q10
y=b ax
R2=R1 Q10 (Т2

Организм – Та. Ta Lim
Температурные пороги жизни
Изменчивость диапазона Та
Изменение Та-толерантности
Организм – Та. Ta Lim
Температурные пороги жизни
Изменчивость диапазона Та
Изменение Та-толерантности

Организм – Та. Ta Lim
Верхний температурный предел
Ни одно животное не
Организм – Та. Ta Lim
Верхний температурный предел
Ни одно животное не

Организм – Та. Ta Lim
Обезвоживание ? повышение устойчивости к Та
Организм – Та. Ta Lim
Обезвоживание ? повышение устойчивости к Та
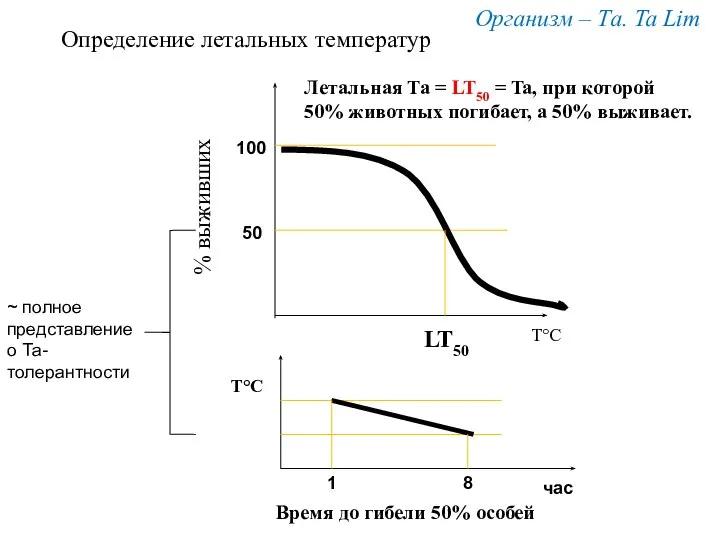
Т°С
% выживших
Организм – Та. Ta Lim
LT50
100
50
Определение летальных температур
Летальная Та =
Т°С
% выживших
Организм – Та. Ta Lim
LT50
100
50
Определение летальных температур
Летальная Та =

LT50 и причины гибели при перегреве
Организм – Та. Ta Lim
У
LT50 и причины гибели при перегреве
Организм – Та. Ta Lim
У

Организм – Та. Ta Lim
Причины тепловой смерти:
Денатурация белков (45-55°С для
Организм – Та. Ta Lim
Причины тепловой смерти:
Денатурация белков (45-55°С для

Организм – Та. Ta Lim
Нижний температурный предел
Рассогласование реакций, имеющих разные
Организм – Та. Ta Lim
Нижний температурный предел
Рассогласование реакций, имеющих разные
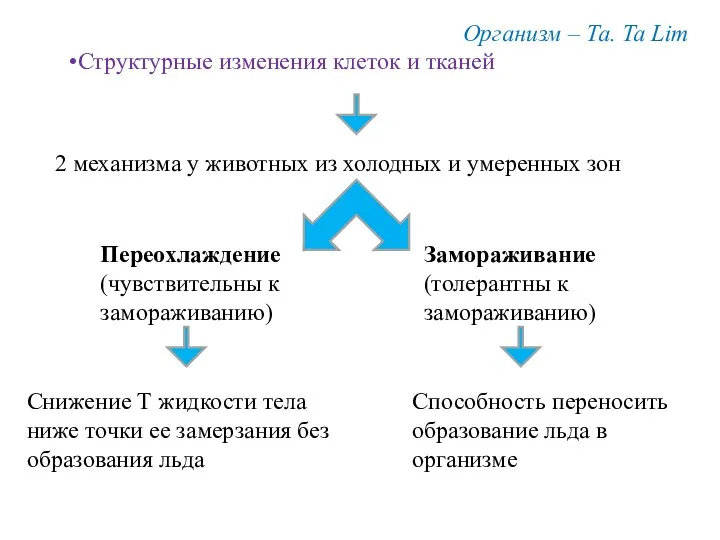
Организм – Та. Ta Lim
Структурные изменения клеток и тканей
2 механизма
Организм – Та. Ta Lim
Структурные изменения клеток и тканей
2 механизма

Организм – Та. Ta Lim
Переохлаждение
При отсутствии чужеродных частиц чистую воду
Организм – Та. Ta Lim
Переохлаждение
При отсутствии чужеродных частиц чистую воду

Организм – Та. Ta Lim
АНТИФРИЗЫ
А
Б
Глицерин – есть у многих насекомых.
Организм – Та. Ta Lim
АНТИФРИЗЫ
А
Б
Глицерин – есть у многих насекомых.
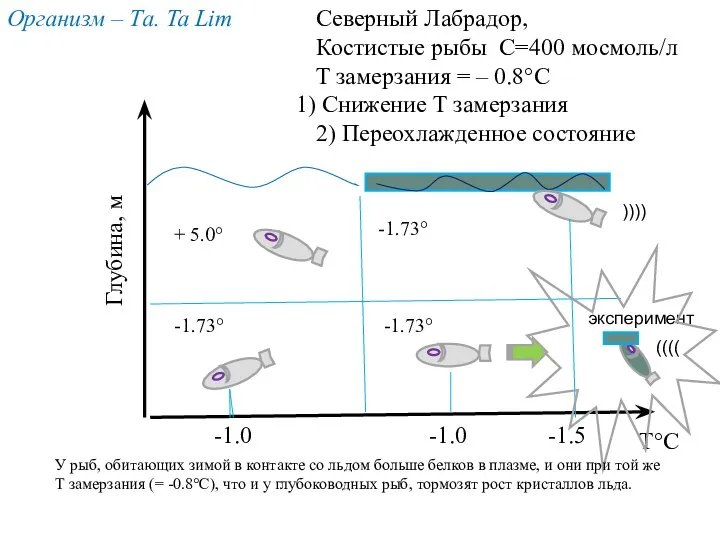
-1.0
-1.0
-1.5
-1.73°
-1.73°
-1.73°
+ 5.0°
T°С
Глубина, м
Северный Лабрадор,
Костистые рыбы С=400 мосмоль/л
T замерзания = –
-1.0
-1.0
-1.5
-1.73°
-1.73°
-1.73°
+ 5.0°
T°С
Глубина, м
Северный Лабрадор,
Костистые рыбы С=400 мосмоль/л
T замерзания = –

Вещество со свойствами антифриза, выделенное из антарктической рыбы
Trematomus, в водном растворе
Вещество со свойствами антифриза, выделенное из антарктической рыбы
Trematomus, в водном растворе

Гликопротеид Trematomus
(2 аминокислоты и дисахарид):
АЛАНИН 23%
ТРЕОНИН 16%
ДИСАХАРИД
Организм – Та. Ta
Гликопротеид Trematomus
(2 аминокислоты и дисахарид):
АЛАНИН 23%
ТРЕОНИН 16%
ДИСАХАРИД
Организм – Та. Ta

Физиологические адаптации к перемене температуры
Организм – Та. Ta Lim
Воздействие сублетальной
Физиологические адаптации к перемене температуры
Организм – Та. Ta Lim
Воздействие сублетальной
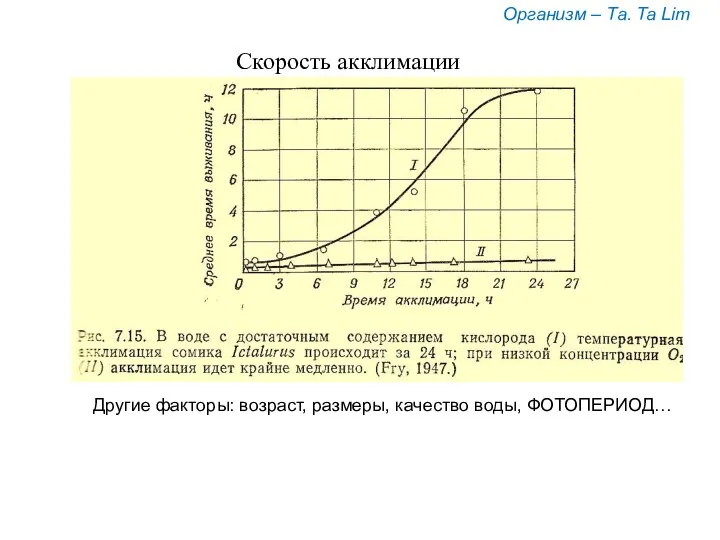
Скорость акклимации
Организм – Та. Ta Lim
Другие факторы: возраст, размеры, качество воды,
Скорость акклимации
Организм – Та. Ta Lim
Другие факторы: возраст, размеры, качество воды,

Сплошная линия очерчивает
полный диапазон Та-толерантности
золотой рыбки (Fry et al., 1942).
У
Сплошная линия очерчивает
полный диапазон Та-толерантности
золотой рыбки (Fry et al., 1942).
У

Организм – Та. Ta Lim
Организм – Та. Ta Lim
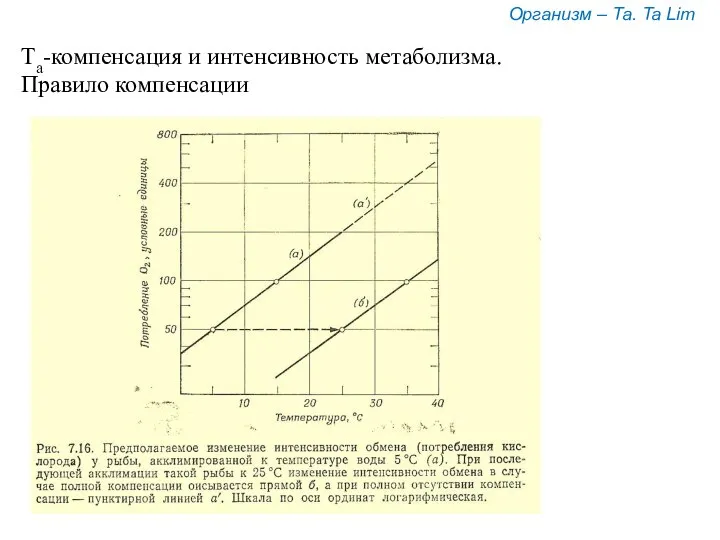
Та-компенсация и интенсивность метаболизма.
Правило компенсации
Организм – Та. Ta Lim
Та-компенсация и интенсивность метаболизма.
Правило компенсации
Организм – Та. Ta Lim
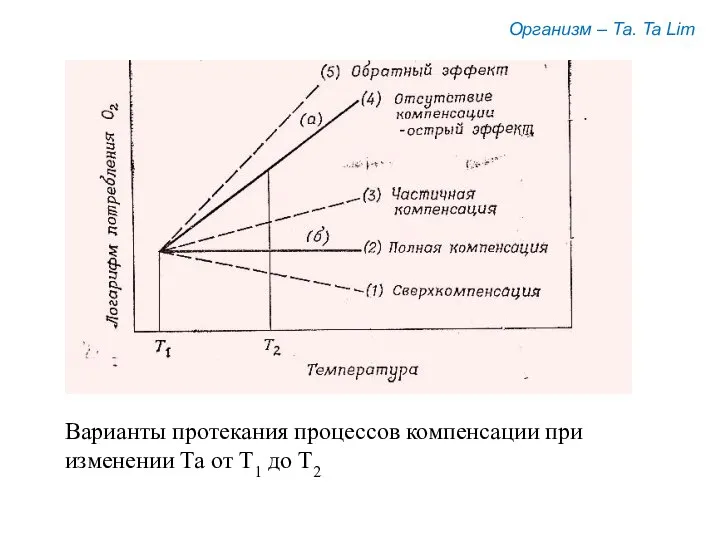
Организм – Та. Ta Lim
Варианты протекания процессов компенсации при
изменении Та
Организм – Та. Ta Lim
Варианты протекания процессов компенсации при
изменении Та
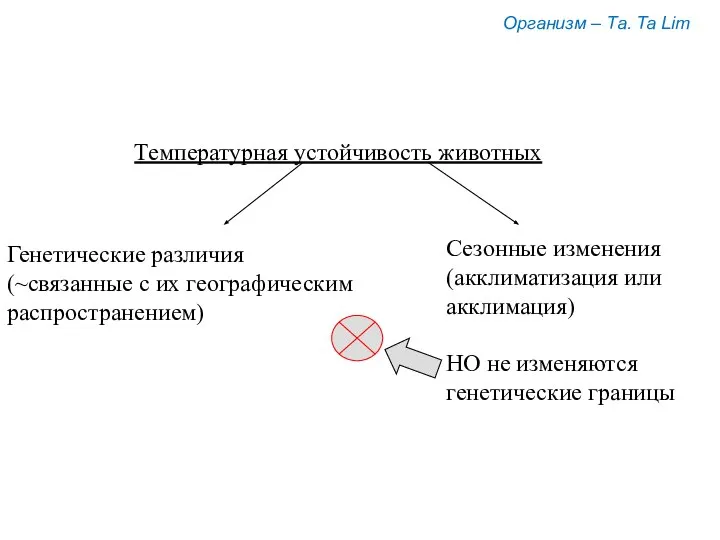
Организм – Та. Ta Lim
Температурная устойчивость животных
Генетические различия
(~связанные с их
Организм – Та. Ta Lim
Температурная устойчивость животных
Генетические различия
(~связанные с их


Research Summary
last updated by G. A. Bartholomew in 1989 and slightly
Research Summary last updated by G. A. Bartholomew in 1989 and slightly

Knut Schmidt-Nielsen
Born in TrondheimBorn in Trondheim, NorwayBorn in Trondheim, Norway. He
Knut Schmidt-Nielsen
Born in TrondheimBorn in Trondheim, NorwayBorn in Trondheim, Norway. He

Scholander, Per Fredrik (Thorkelsson) (Norway- United States 1905-1980)
physiological ecology
Physiologist Scholander became
Scholander, Per Fredrik (Thorkelsson) (Norway- United States 1905-1980)
physiological ecology
Physiologist Scholander became

СЛОНИМ Абрам Донович (1903, Петровск-ПортСЛОНИМ Абрам Донович (1903, Петровск-Порт Дагестанской обл.
СЛОНИМ Абрам Донович (1903, Петровск-ПортСЛОНИМ Абрам Донович (1903, Петровск-Порт Дагестанской обл.
 ЭКОволонтёрство
ЭКОволонтёрство Красная книга НСО
Красная книга НСО Разделы экологии
Разделы экологии Экология и промышленность
Экология и промышленность Презентация на тему Животные -космонавты
Презентация на тему Животные -космонавты  Материальные потоки веществ в лесных экосистемах
Материальные потоки веществ в лесных экосистемах Способы утилизации ТБО
Способы утилизации ТБО Вода и здоровье
Вода и здоровье Животное из красной книги. Манул
Животное из красной книги. Манул Электронный учебник по разделу экологии – популяция животных
Электронный учебник по разделу экологии – популяция животных Экологические проблемы озера Байкал
Экологические проблемы озера Байкал Классификация рисков
Классификация рисков Осознание и поиск способов преодоления мирового экологического кризиса
Осознание и поиск способов преодоления мирового экологического кризиса Экология жилища
Экология жилища Чорнобильська катастрофа
Чорнобильська катастрофа Экологическое нормирование воздействий на атмосферу
Экологическое нормирование воздействий на атмосферу Северное море
Северное море Вплив пилу на здоров’я людини
Вплив пилу на здоров’я людини ЮНЕСКО. Список особо охраняемых природных территорий
ЮНЕСКО. Список особо охраняемых природных территорий Презентация на тему Такие разные праздники
Презентация на тему Такие разные праздники  Going green: in danger
Going green: in danger Химические свойства атмосферного воздуха, влияние на здоровье. Тесты. (Лекция 2)
Химические свойства атмосферного воздуха, влияние на здоровье. Тесты. (Лекция 2) Конференция "Планета в пластиковой упаковке"
Конференция "Планета в пластиковой упаковке" Пасивне будівництво
Пасивне будівництво Сохраним и преумножим богатство нашего бора
Сохраним и преумножим богатство нашего бора Презентация Рысь
Презентация Рысь  Экологиялық факторлар
Экологиялық факторлар Экологическая политика ЕС
Экологическая политика ЕС