- БИОХИМИЯ ПЕЧЕНИ

Содержание
- 2. План лекции Строение и функции печени Обмен веществ в печени Обмен веществ абсобтивный и постабсобтивний периоды
- 3. Печень – самый крупный орган в организме человека, состоит примерно из 300 млрд клеток, 80% из
- 4. Между эндотелием и гепатоцитами расположено перисинусоидальное пространство – пространство Диссе. Вдоль синусоид располагаются клетки ретикуло- эндотелиальной
- 5. Печень перекрещивает метаболизм углеводов, липидов и белков Печень, являясь центральным органом метаболизма, участвует в поддержании метаболического
- 6. глицерол-3-фосфат, образуемый из диоксиацетонфосфата гликолиза, направляется для синтеза триацилглицеролов. Также для этой цели может быть использован
- 7. ФУНКЦИИ ПЕЧЕНИ метаболическая, депонирующая, барьерная, экскреторная и гомеостатическая
- 8. ФУНКЦИИ ПЕЧЕНИ Метаболическая. Продукты расщепления питательных веществ поступают в печень из пищеварительного тракта через воротную вену.
- 9. Обмен веществ в печени Печень принимает участие в метаболизме почти всех классов веществ
- 10. КОМПЕНСАТОРНЫЕ функции печени Ткани высших организмов нуждаются в постоянном притоке богатых энергией веществ и предшественников для
- 11. ФАЗА АБСОРБЦИИ Фаза абсорбции (утилизации и депонирования) начинается непосредственно с приема пищи и длится примерно 2–4
- 12. Поджелудочная железа отвечает на это изменением выброса гормонов: увеличением секреции инсулина и уменьшением секреции глюкагона. Увеличение
- 13. Фаза постабсорбции При прекращении поступления пищи вскоре начинается фаза постабсорбции. Эта стадия начинается с изменения секреции
- 14. В печени происходит мобилизация гликогена (гликогенолиз). Полученная глюкоза используется для обеспечения других тканей, прежде всего мозга,
- 15. Метаболизм углеводов Глюкоза наряду с жирными кислотами и кетоновыми телами является важнейшим источником энергии. Уровень глюкозы
- 16. Метаболизм липидов Если во время приема пищи в печень поступает избыток глюкозы, который не используется для
- 17. Белковый обмен Больше половины синтезируемого за сутки в организме белка приходится на печень. Скорость обновления всех
- 18. Тесное взаимодействие синтеза мочевины и ЦТК
- 19. Пигментный обмен Участие печени в пигментном обмене заключается в превращении гидрофобного билирубина в гидрофильную форму и
- 20. Оценка метаболической функции В клинической практике существуют приемы оценки той или иной функции: Участие в углеводном
- 21. Механизмы обезвреживания токсических веществ в печени
- 22. Механизмы обезвреживания токсических веществ в печени
- 23. Инактивация чужеродных веществ в организме Чужеродные вещества, попадающие в организм из ЖКТ, через кожу и лёгкие
- 24. Метаболизм и выведение ксенобиотиков из организма: R — радикал, используемый при конъюгации (глутатион, глюкуронил и др.);
- 25. В мембранах эндоплазматического ретикулума (ЭР) практически всех тканей локализована система микросомального (монооксигеназного) окисления (МСО), отвечающая за
- 26. Система микросомального окисления веществ и реакции коньюгации Процесс состоит из одного или двух этапов и сводится
- 27. Положение белков монооксигеназной системы в мембране
- 28. В результате 1этапа - гидроксилирования возможны: повышение растворимости гидрофобного соединения, потеря молекулой ее биологической активности или
- 29. 2 этап инактивации – реакция коньюгации модифицированных на 1 этапе или содержащих полярные группы веществ. Они
- 30. Схема реакций коньюгации RОН –ксенобиотик, образующийся в результате микросомального окисления GSН- глутатион
- 31. ФАФС – донор остатка серной кислоты ( и для образования солей сульфатов аммония в печени), образуется
- 32. Активная форма метионина S-аденозилметионин – донор метильных групп в различных реакциях метаболизма и обезвреживания ксенобиотиков различного
- 33. Особое место среди ферментов, участвующих в обезвреживании ксенобиотиков, нормальных метаболитов и лекарств, занимают глутатионтрансферазы. Известно множество
- 34. • конъюгацией остатка субстрата R с глутатионом (GSH): R + GSH → GSRH; • нуклеофильными замещениями:
- 35. Глутатион и димер глутатиона или восстановленный и окисленный глутатион
- 36. ↑ Димер глутатиона или глутатион окисленный Восстановление осуществляет глутатион редуктаза коферментом которой является НАДФН+Н
- 37. Обезвреживание продуктов жизнедеятельности микрофлоры кишечника Образование токсичных продуктов из аминокислот под действием микроорганизмов кишечника называют гниением
- 38. Реакции коньюгации индола и скатола
- 39. Биотрансформация лекарств печени Лекарства выводятся из организма в неизменном виде или подвергаются химической модификации. Последняя может
- 40. Реакция инактивации аспирина Инактивация сульфаниламидов
- 41. Парацетамол (ацетаминофен), входит в состав многих обезболивающих препаратов: гриппостат, фервекс, пенталгин, может сразу коньюгировать с УДФ-глюкуроновой
- 42. Парацетамол (ацетаминофен), входит в состав многих обезболивающих препаратов: гриппостат, фервекс, пенталгин; При обезвреживании может сразу коньюгировать
- 43. Лидокаин (ацетанилид), применяется в стоматологии для местного обезболивания, сначала гидролизуется с образованием уксусной кислоты и аминобензола.
- 44. Важную роль в выведении из клеток гидрофобных ксенобиотиков играет транспортная АТФ-аза, которую называют Р-гликопротеин. Этот интегральный
- 45. Опять кровь
- 46. Основные механизмы фагоцитоза От микрооорганизмов поступающих в организм человека, клетки защищаются фагоцитозом. Основную роль в этом
- 47. Образование активных форм кислорода активированными макрофагами, нейтрофилами и эозинофилами в процессе респираторного взрыва
- 48. Активирующийся при фагоцитозе ферментный комплекс мембран фагосом НАДФН-оксидаза используя кислород, катализирует образование супероксидного аниона: 2О2 +
- 49. Благодарю за внимание
- 51. Скачать презентацию

План лекции
Строение и функции печени
Обмен веществ в печени
Обмен веществ абсобтивный и
План лекции
Строение и функции печени
Обмен веществ в печени
Обмен веществ абсобтивный и

Печень – самый крупный орган в организме человека, состоит
примерно из
Печень – самый крупный орган в организме человека, состоит
примерно из
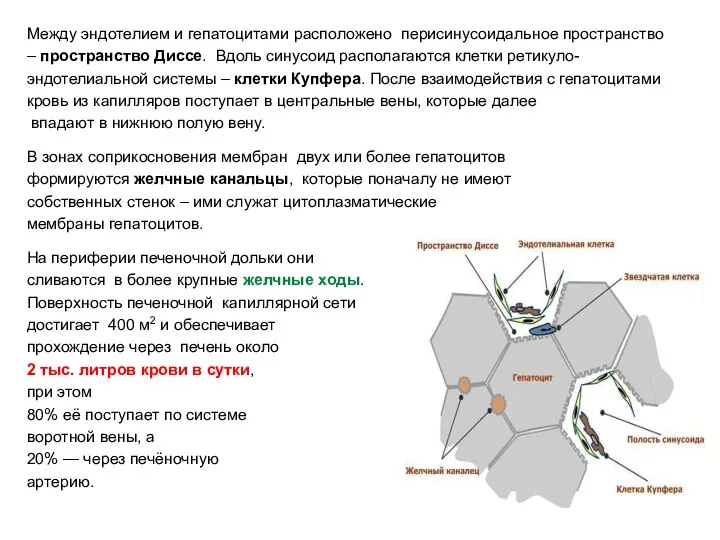
Между эндотелием и гепатоцитами расположено перисинусоидальное пространство
– пространство Диссе. Вдоль синусоид располагаются
Между эндотелием и гепатоцитами расположено перисинусоидальное пространство
– пространство Диссе. Вдоль синусоид располагаются

Печень перекрещивает метаболизм
углеводов, липидов и белков
Печень, являясь центральным органом метаболизма,
Печень перекрещивает метаболизм
углеводов, липидов и белков
Печень, являясь центральным органом метаболизма,
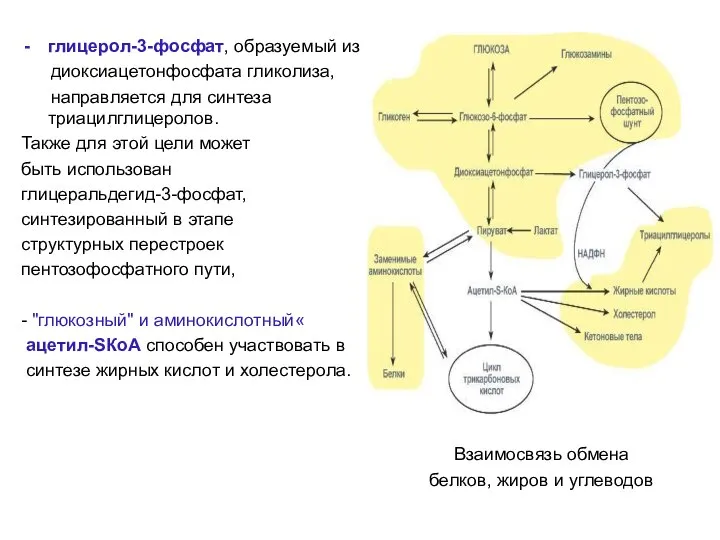
глицерол-3-фосфат, образуемый из
диоксиацетонфосфата гликолиза,
направляется для синтеза триацилглицеролов.
глицерол-3-фосфат, образуемый из
диоксиацетонфосфата гликолиза,
направляется для синтеза триацилглицеролов.
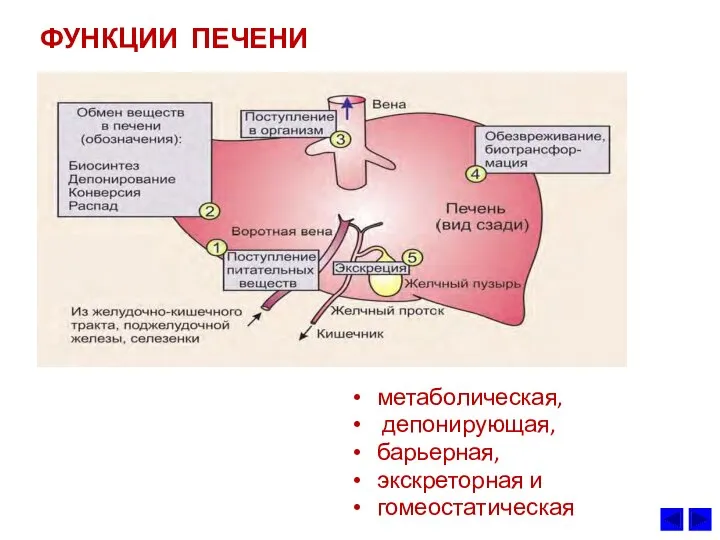
ФУНКЦИИ ПЕЧЕНИ
метаболическая,
депонирующая,
барьерная,
экскреторная и
гомеостатическая
ФУНКЦИИ ПЕЧЕНИ
метаболическая,
депонирующая,
барьерная,
экскреторная и
гомеостатическая

ФУНКЦИИ ПЕЧЕНИ
Метаболическая. Продукты расщепления питательных веществ поступают в
печень из пищеварительного тракта
ФУНКЦИИ ПЕЧЕНИ
Метаболическая. Продукты расщепления питательных веществ поступают в
печень из пищеварительного тракта

Обмен веществ в печени
Печень принимает участие в метаболизме почти всех классов
Обмен веществ в печени
Печень принимает участие в метаболизме почти всех классов

КОМПЕНСАТОРНЫЕ функции печени
Ткани высших организмов нуждаются в постоянном притоке
богатых энергией
КОМПЕНСАТОРНЫЕ функции печени
Ткани высших организмов нуждаются в постоянном притоке
богатых энергией
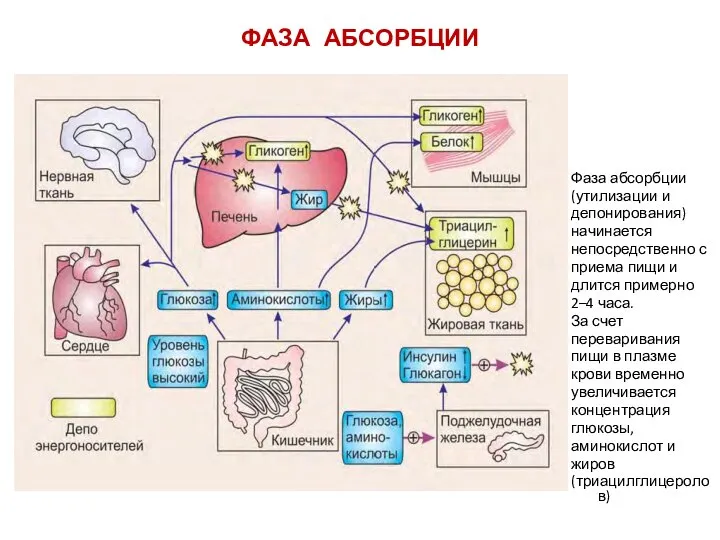
ФАЗА АБСОРБЦИИ
Фаза абсорбции
(утилизации и
депонирования)
начинается
непосредственно с
приема пищи
ФАЗА АБСОРБЦИИ
Фаза абсорбции
(утилизации и
депонирования)
начинается
непосредственно с
приема пищи

Поджелудочная железа отвечает на это изменением выброса гормонов:
увеличением секреции инсулина
Поджелудочная железа отвечает на это изменением выброса гормонов:
увеличением секреции инсулина
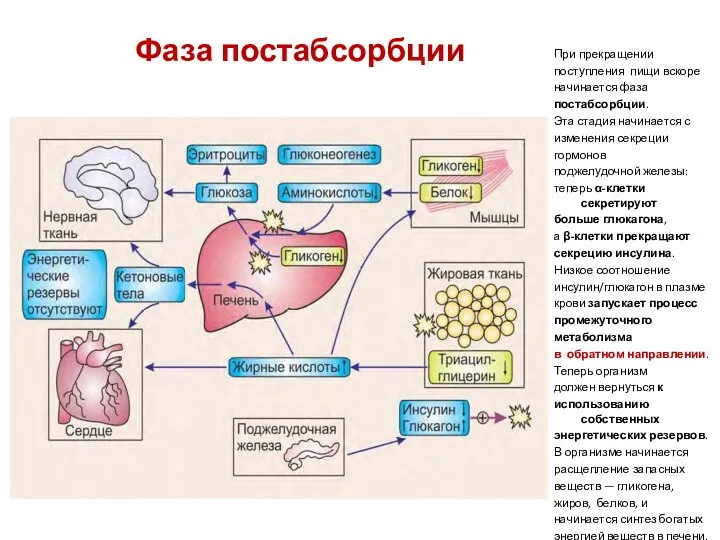
Фаза постабсорбции
При прекращении
поступления пищи вскоре
начинается фаза
постабсорбции.
Эта стадия начинается
Фаза постабсорбции
При прекращении
поступления пищи вскоре
начинается фаза
постабсорбции.
Эта стадия начинается

В печени происходит мобилизация гликогена (гликогенолиз).
Полученная глюкоза используется для обеспечения
В печени происходит мобилизация гликогена (гликогенолиз).
Полученная глюкоза используется для обеспечения

Метаболизм углеводов
Глюкоза наряду с жирными кислотами и кетоновыми телами является важнейшим
Метаболизм углеводов
Глюкоза наряду с жирными кислотами и кетоновыми телами является важнейшим

Метаболизм липидов
Если во время приема пищи в печень поступает избыток глюкозы,
Метаболизм липидов
Если во время приема пищи в печень поступает избыток глюкозы,

Белковый обмен
Больше половины синтезируемого за сутки в организме белка
приходится на
Белковый обмен
Больше половины синтезируемого за сутки в организме белка
приходится на
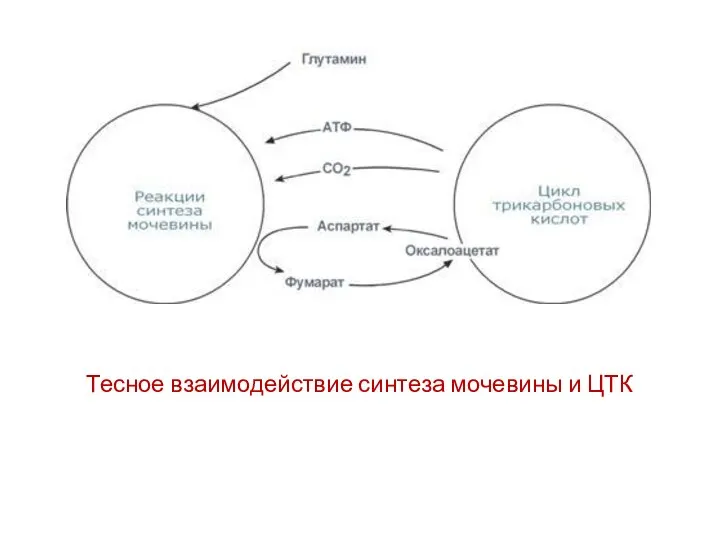
Тесное взаимодействие синтеза мочевины и ЦТК
Тесное взаимодействие синтеза мочевины и ЦТК

Пигментный обмен
Участие печени в пигментном обмене заключается в
превращении гидрофобного билирубина
Пигментный обмен
Участие печени в пигментном обмене заключается в
превращении гидрофобного билирубина

Оценка метаболической функции
В клинической практике существуют приемы оценки той или иной
Оценка метаболической функции
В клинической практике существуют приемы оценки той или иной

Механизмы обезвреживания токсических веществ в печени
Механизмы обезвреживания токсических веществ в печени
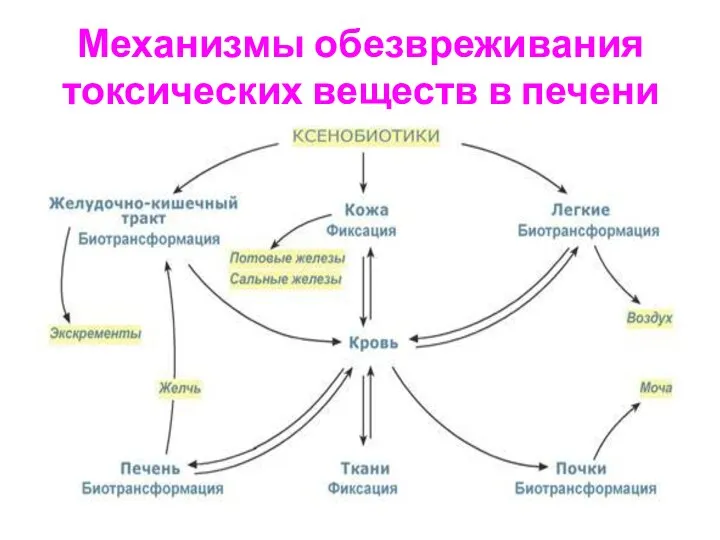
Механизмы обезвреживания токсических веществ в печени
Механизмы обезвреживания токсических веществ в печени

Инактивация чужеродных веществ в организме
Чужеродные вещества, попадающие в организм из ЖКТ,
Инактивация чужеродных веществ в организме
Чужеродные вещества, попадающие в организм из ЖКТ,
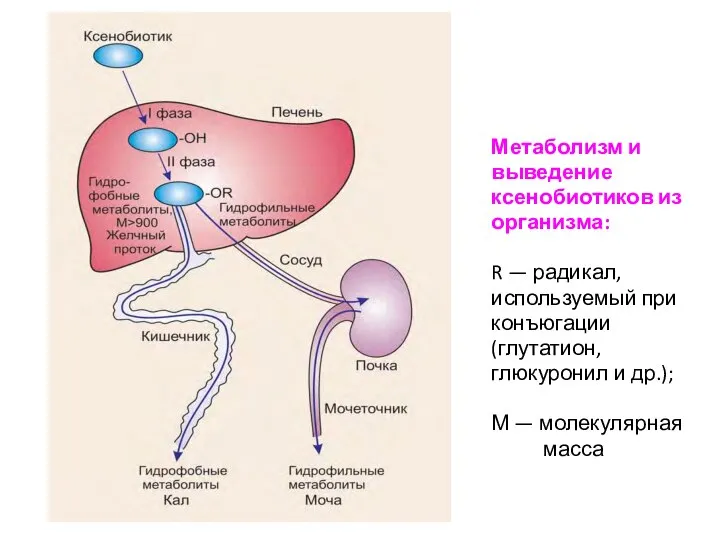
Метаболизм и
выведение
ксенобиотиков из
организма:
R — радикал,
используемый при
конъюгации
Метаболизм и
выведение
ксенобиотиков из
организма:
R — радикал,
используемый при
конъюгации

В мембранах эндоплазматического ретикулума (ЭР) практически всех тканей
локализована система микросомального
В мембранах эндоплазматического ретикулума (ЭР) практически всех тканей
локализована система микросомального
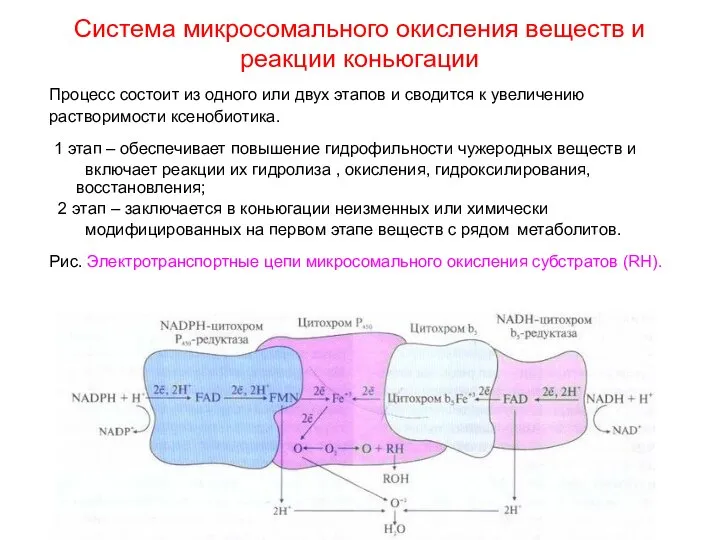
Система микросомального окисления веществ и реакции коньюгации
Процесс состоит из одного или
Система микросомального окисления веществ и реакции коньюгации
Процесс состоит из одного или
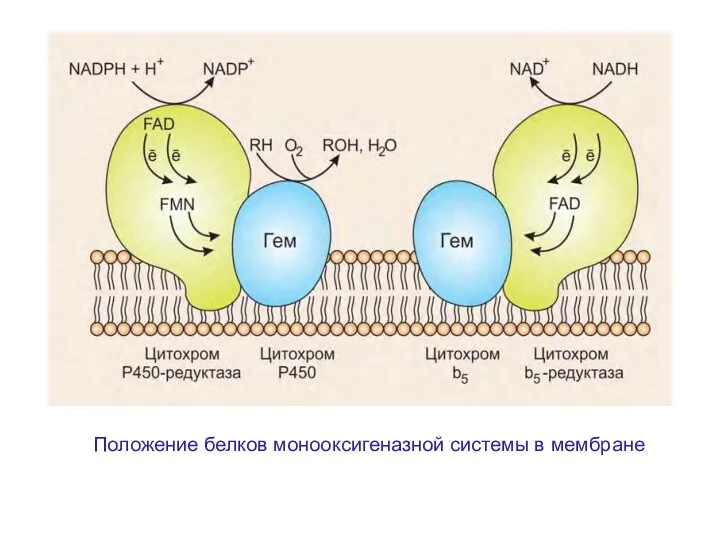
Положение белков монооксигеназной системы в мембране
Положение белков монооксигеназной системы в мембране

В результате 1этапа - гидроксилирования возможны:
повышение растворимости гидрофобного соединения,
потеря молекулой
В результате 1этапа - гидроксилирования возможны:
повышение растворимости гидрофобного соединения,
потеря молекулой

2 этап инактивации – реакция коньюгации модифицированных на
1 этапе или содержащих
2 этап инактивации – реакция коньюгации модифицированных на
1 этапе или содержащих
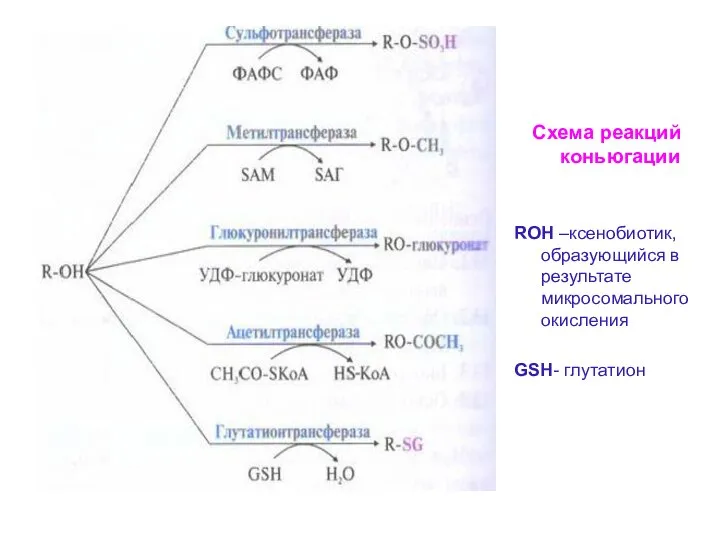
Схема реакций коньюгации
RОН –ксенобиотик, образующийся в результате микросомального окисления
GSН- глутатион
Схема реакций коньюгации
RОН –ксенобиотик, образующийся в результате микросомального окисления
GSН- глутатион
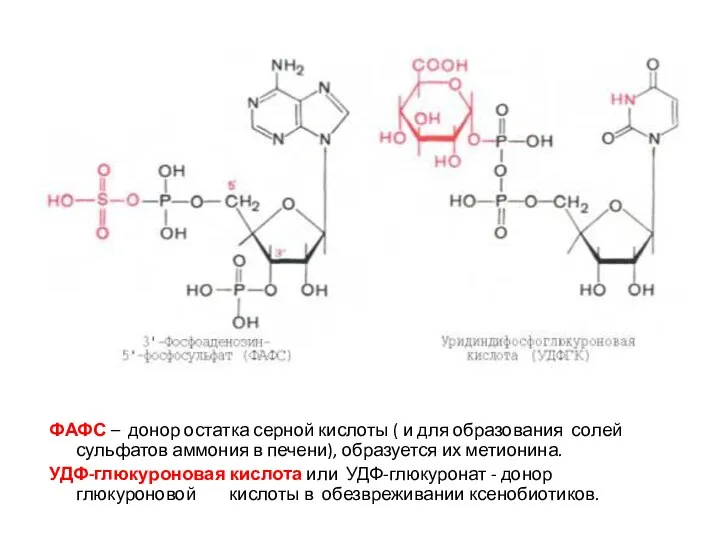
ФАФС – донор остатка серной кислоты ( и для образования солей
ФАФС – донор остатка серной кислоты ( и для образования солей
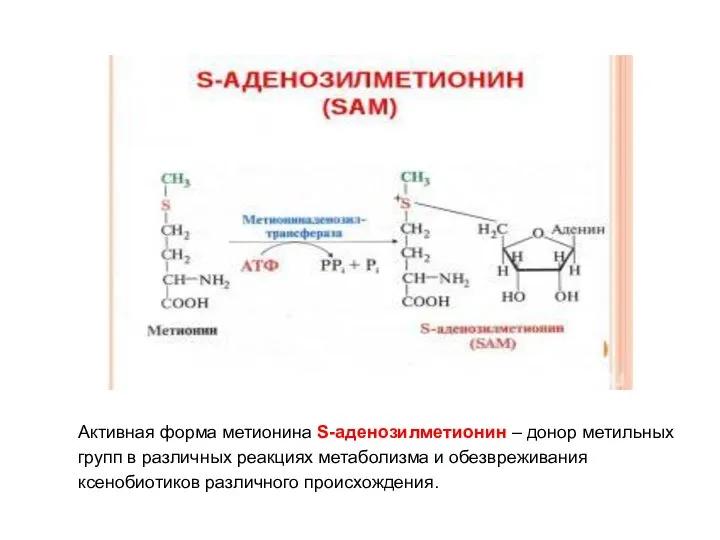
Активная форма метионина S-аденозилметионин – донор метильных
групп в различных реакциях метаболизма
Активная форма метионина S-аденозилметионин – донор метильных
групп в различных реакциях метаболизма

Особое место среди ферментов, участвующих в обезвреживании
ксенобиотиков, нормальных метаболитов и
Особое место среди ферментов, участвующих в обезвреживании
ксенобиотиков, нормальных метаболитов и
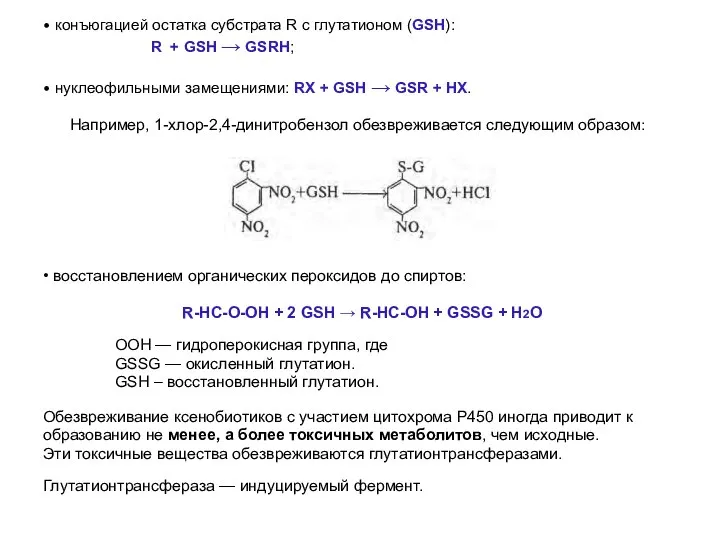
• конъюгацией остатка субстрата R с глутатионом (GSH):
R + GSH →
• конъюгацией остатка субстрата R с глутатионом (GSH):
R + GSH →
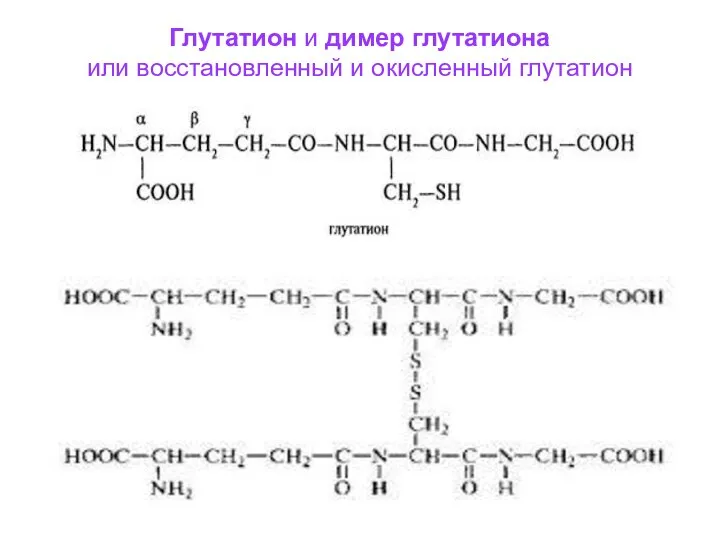
Глутатион и димер глутатиона
или восстановленный и окисленный глутатион
Глутатион и димер глутатиона
или восстановленный и окисленный глутатион
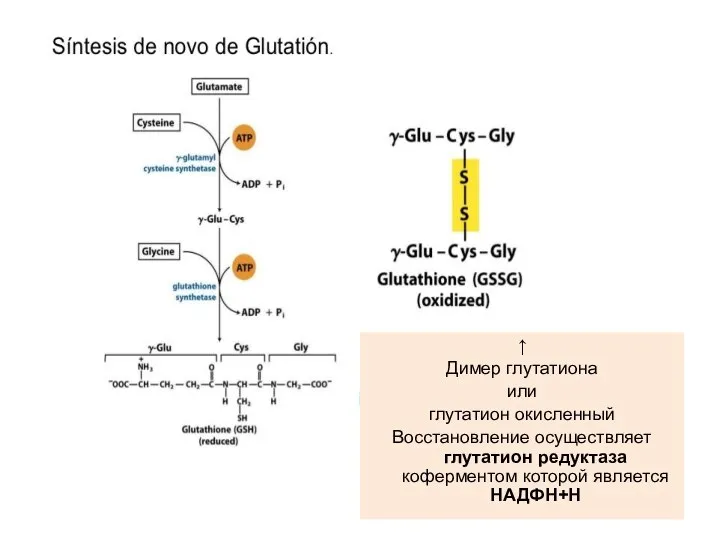
↑
Димер глутатиона
или
глутатион окисленный
Восстановление осуществляет глутатион редуктаза коферментом которой является
↑
Димер глутатиона
или
глутатион окисленный
Восстановление осуществляет глутатион редуктаза коферментом которой является
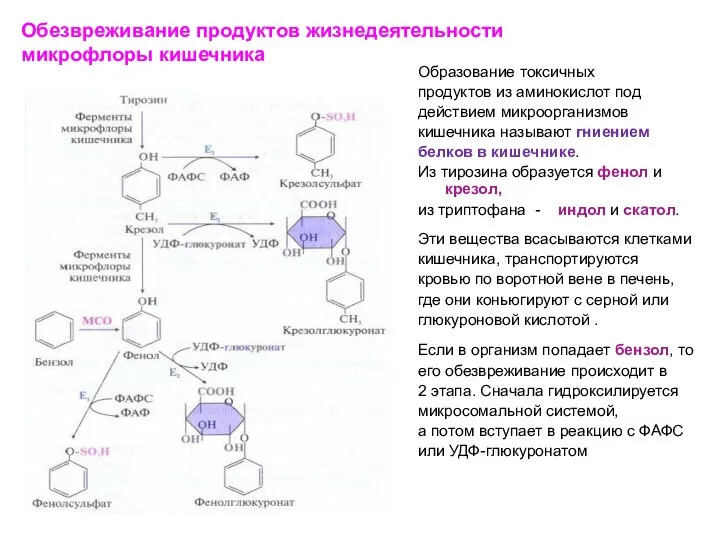
Обезвреживание продуктов жизнедеятельности
микрофлоры кишечника
Образование токсичных
продуктов из аминокислот под
действием микроорганизмов
Обезвреживание продуктов жизнедеятельности
микрофлоры кишечника
Образование токсичных
продуктов из аминокислот под
действием микроорганизмов
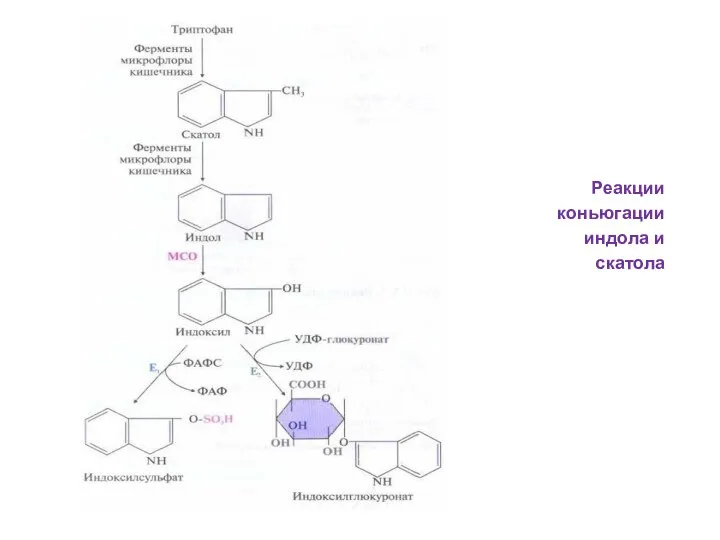
Реакции
коньюгации
индола и
скатола
Реакции
коньюгации
индола и
скатола

Биотрансформация лекарств печени
Лекарства выводятся из организма в неизменном виде или подвергаются
химической
Биотрансформация лекарств печени
Лекарства выводятся из организма в неизменном виде или подвергаются
химической
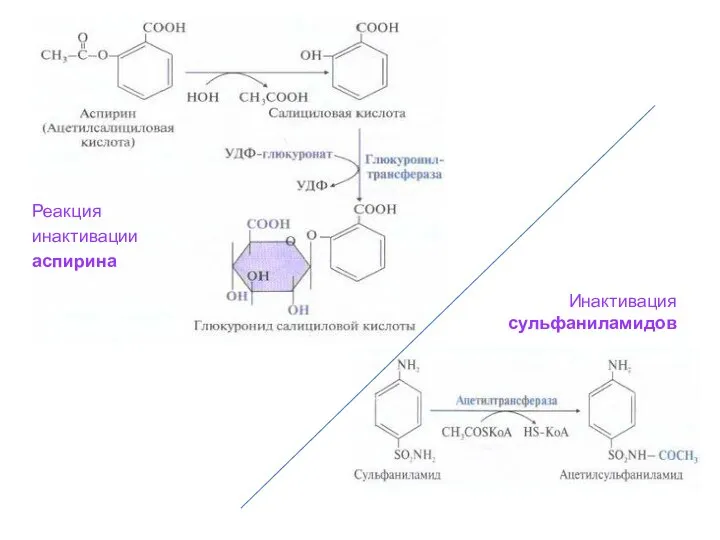
Реакция
инактивации
аспирина
Инактивация сульфаниламидов
Реакция
инактивации
аспирина
Инактивация сульфаниламидов

Парацетамол (ацетаминофен), входит в состав многих
обезболивающих препаратов: гриппостат,
фервекс,
пенталгин,
может сразу коньюгировать
Парацетамол (ацетаминофен), входит в состав многих
обезболивающих препаратов: гриппостат,
фервекс,
пенталгин,
может сразу коньюгировать
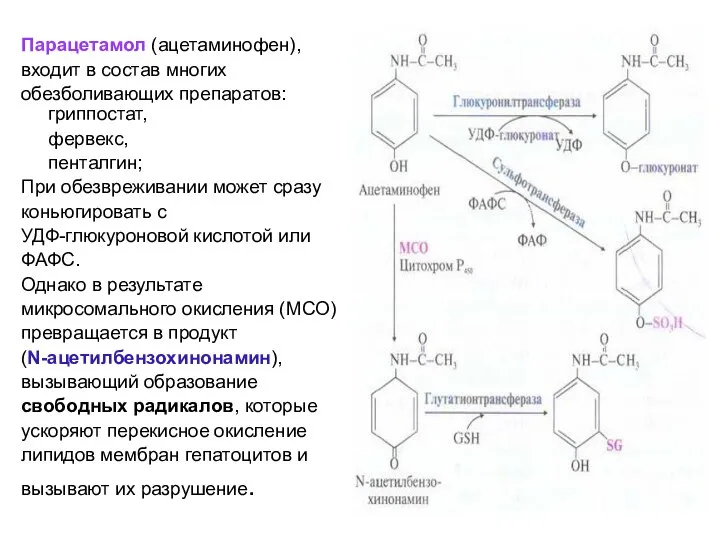
Парацетамол (ацетаминофен),
входит в состав многих
обезболивающих препаратов: гриппостат,
фервекс,
пенталгин;
При обезвреживании может
Парацетамол (ацетаминофен),
входит в состав многих
обезболивающих препаратов: гриппостат,
фервекс,
пенталгин;
При обезвреживании может
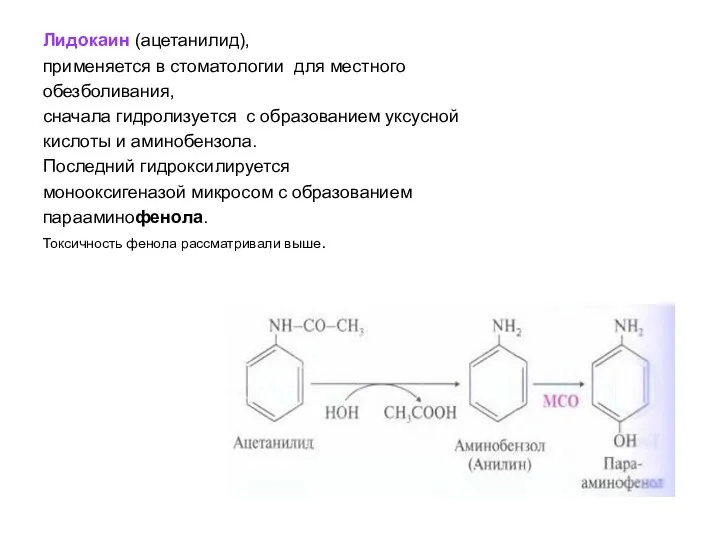
Лидокаин (ацетанилид),
применяется в стоматологии для местного
обезболивания,
сначала гидролизуется с
Лидокаин (ацетанилид),
применяется в стоматологии для местного
обезболивания,
сначала гидролизуется с
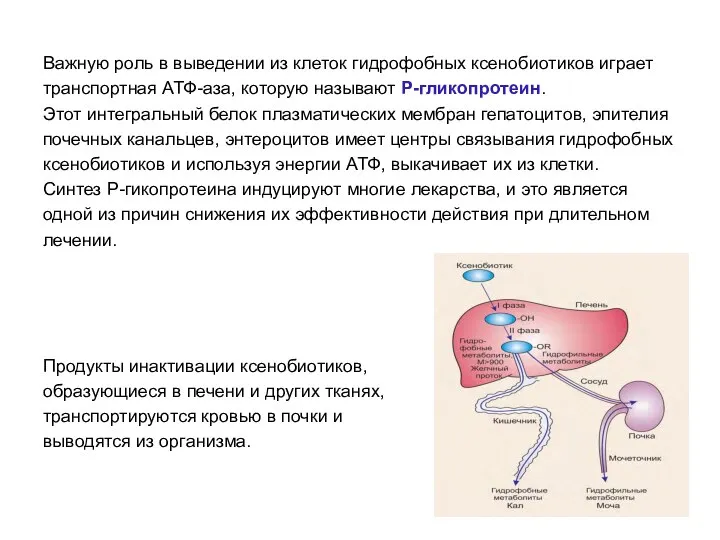
Важную роль в выведении из клеток гидрофобных ксенобиотиков играет
транспортная АТФ-аза,
Важную роль в выведении из клеток гидрофобных ксенобиотиков играет
транспортная АТФ-аза,

Опять кровь
Опять кровь
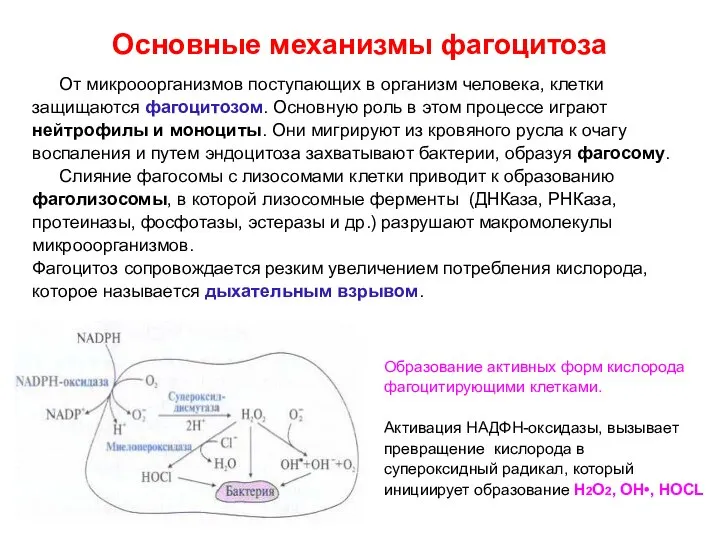
Основные механизмы фагоцитоза
От микрооорганизмов поступающих в организм человека, клетки
защищаются фагоцитозом.
Основные механизмы фагоцитоза
От микрооорганизмов поступающих в организм человека, клетки
защищаются фагоцитозом.
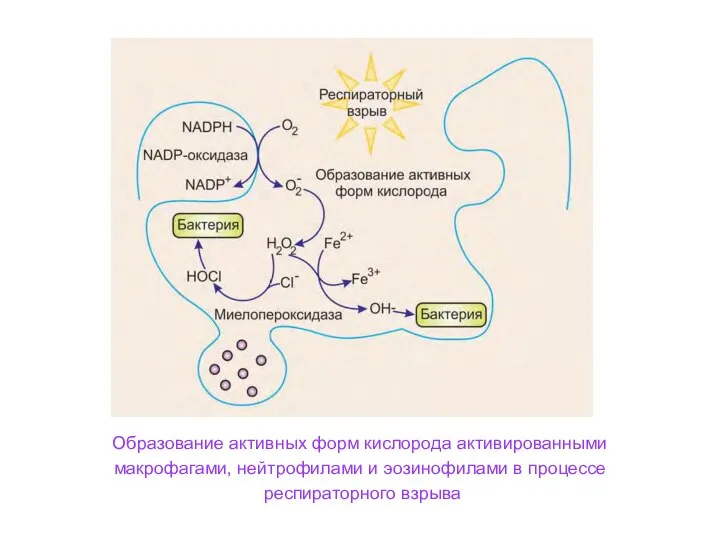
Образование активных форм кислорода активированными
макрофагами, нейтрофилами и эозинофилами в процессе
Образование активных форм кислорода активированными
макрофагами, нейтрофилами и эозинофилами в процессе
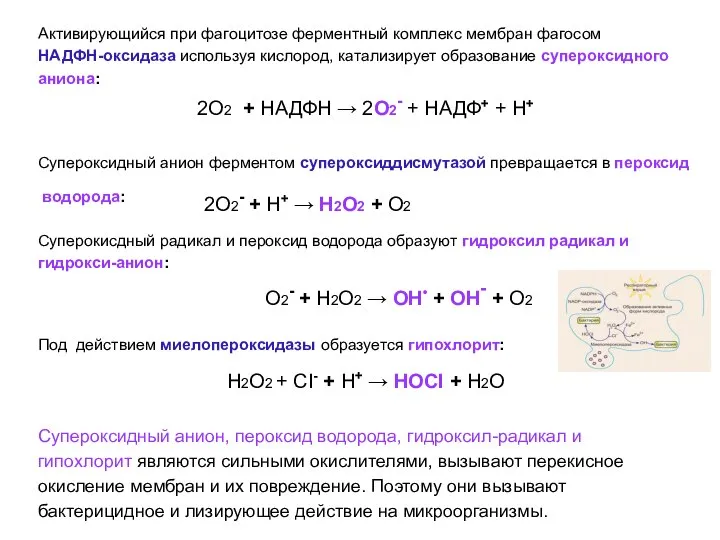
Активирующийся при фагоцитозе ферментный комплекс мембран фагосом
НАДФН-оксидаза используя кислород, катализирует
Активирующийся при фагоцитозе ферментный комплекс мембран фагосом
НАДФН-оксидаза используя кислород, катализирует

Благодарю за внимание
Благодарю за внимание
 Бақытты сәм
Бақытты сәм Қазақтың ұлттық ойындары
Қазақтың ұлттық ойындары Презентация "Ненецкая кукла – нгухуко" - скачать презентации по МХК
Презентация "Ненецкая кукла – нгухуко" - скачать презентации по МХК Banya. A russian tradition
Banya. A russian tradition Основы применения языка SFC в CoDeSys. Реализация многозадачности
Основы применения языка SFC в CoDeSys. Реализация многозадачности Північна Корея
Північна Корея Форма государства. Форма правления. Монархия и республика
Форма государства. Форма правления. Монархия и республика Клеедощатые балки
Клеедощатые балки Функции финансового менеджмента в процессе осуществления ликвидационных процедур при банкротстве
Функции финансового менеджмента в процессе осуществления ликвидационных процедур при банкротстве Главное политическое событие декабря 1991 года - распад СССР
Главное политическое событие декабря 1991 года - распад СССР Технологический процесс изготовления детали «Соединительная втулка»
Технологический процесс изготовления детали «Соединительная втулка» Управление рисками
Управление рисками Государственное регулирование предпринимательской деятельности
Государственное регулирование предпринимательской деятельности Участие в разработке ИС. Создание WEB-сайта «Магазин канцелярских товаров»
Участие в разработке ИС. Создание WEB-сайта «Магазин канцелярских товаров» Футбольная команда «Рассвет»
Футбольная команда «Рассвет» Тоталітарна політична культура
Тоталітарна політична культура Кредит и кредитная система
Кредит и кредитная система Что проверяет инспектор в кадровых документах. Бесплатный онлайн-курс
Что проверяет инспектор в кадровых документах. Бесплатный онлайн-курс Технология строительного производства. Оборудование строительных площадок
Технология строительного производства. Оборудование строительных площадок История куклы Барби
История куклы Барби Антигены. Антитела. Реакции агглютинации и преципитации.
Антигены. Антитела. Реакции агглютинации и преципитации. Жители саванны - презентация для начальной школы_
Жители саванны - презентация для начальной школы_ Накшатры и брак
Накшатры и брак Творческий проект Тема: «ЛОГАРИФМЫ В АСТРОНОМИИ»
Творческий проект Тема: «ЛОГАРИФМЫ В АСТРОНОМИИ»  История изобретения турбин
История изобретения турбин Презентация по предмету: «Региональная экономика и управление» на тему: «Основные угрозы экономической безопасности.» 2011
Презентация по предмету: «Региональная экономика и управление» на тему: «Основные угрозы экономической безопасности.» 2011 Klikounova Publisher представляет
Klikounova Publisher представляет ИГРОВЫЕ ТЕХНОЛОГИИ В ОБУЧЕНИИ иностранному языку
ИГРОВЫЕ ТЕХНОЛОГИИ В ОБУЧЕНИИ иностранному языку