- Депонирование и мобилизация жиров

Содержание
- 2. Обмен триацилглицеринов С пищей в сутки поступает 70 г ТГ. Эндогенный синтез ТГ идёт в: печени,
- 3. Обмен липидов
- 4. Функции резервных жиров резервуар энергии, теплоизоляционная, защита от механических травм.
- 5. Транспорт ТГ осуществляют ХМ, ЛПОНП.
- 6. Липопротеидлипаза осуществляет гидролиз ТГ. В жировой ткани жир накапливается за счёт поступления из ЛП, образования из
- 7. Тканевые липазы активируются адреналином, глюкагоном, АКТГ.
- 8. Мобилизация депонированных жиров происходит путём их гидролиза до жирных кислот и глицерина липазами жировых клеток. Жирные
- 9. Окисление глицерина Глицеролкиназа АТФ АДФ Mg2+ Глицерин Глицерол-3-фосфат
- 10. Глицеролфосфат- дегидрогеназа НАД НАДН+Н+ Глицерол-3-фосфат Диоксиацетонфосфат
- 11. CO2 + H2O + E Лактат Анаэробный путь Аэробный путь ФГА
- 12. Диоксиацетон Триозофосфат- изомераза Фосфоглицериновый альдегид
- 13. + НАД+ + H3PO4 Фосфоглицериновый альдегид + НАДН+Н+ Глицеральдегидфосфат дегидрогеназа 1,3-дифосфоглицерат
- 14. Фосфоглицераткиназа 1,3-дифосфоглицерат 3-фосфоглицерат АДФ АТФ Mg2+
- 15. 3-фосфоглицерат Фосфоглицеро мутаза 2-фосфоглицерат
- 16. 2-фосфоглицерат Енолаза Н2О Mg2+ Фосфоенолпируват
- 17. Фосфоенолпируват Пируваткиназа Пируват (ПВК) АДФ АТФ Mg2+
- 18. Пируват (ПВК) Ацетил- КоА Цикл Кребса
- 19. Цикл Кребса Цитрат-синтаза Аконитат- гидратаза Аконитат- гидратаза Изоцитрат- дегидрогеназа α-Кетоглутарат дегидрогеназный комплекс Сукцинил-КоА -синтетаза Сукцинат- дегидрогеназа
- 20. CH2-COOH | HOC-COOH | H2C-COOH Цитрат Цикл трикарбоновых кислот Цитратсинтаза
- 21. COOH | CH2 | HO-C-COOH | H-C-H | COOH Цитрат COOH | CH2 | C-COOH ||
- 22. COOH | CH2 | H-C-COOH | HO-C-H | COOH Изоцитрат COOH | CH2 | CH2 |
- 23. COOH | CH2 | CH2 | C=O | COOH L-кетоглутарат COOH | CH2 | СН2 |
- 24. COOH | CH2 | СН2 | C=O S-KoA Cукцинил-КоА + Фн COOH | CH2 | СН2
- 25. COOH | CH2 | СН2 | COOH Сукцинат COOH | CH || HС | COOH Фумарат
- 26. COOH | CH || HС | COOH Фумарат COOH | HO-C-H | H-С-H | COOH Малат
- 27. COOH | HO-C-H | H-С-H | COOH Малат COOH | C=O | СH2 | COOH Оксалоацетат
- 28. Баланс аэробного распада глицерина От глицерина до ФГА затрата – 1 АТФ и получение + 3
- 29. ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ аэробный процесс. В катаболизме жирных кислот выделяют 3 части: β-окисление, ЦТК, дыхательная цепь.
- 30. Кнооп установил, что окисление идёт в β-положении. Ленинджер и Кеннеди установили, что процесс протекает в митохондриях
- 31. Активация жирных кислот происходит на наружной поверхности мембраны митохондрий. R-COOH + HS-KoA +АТФ + АМФ +
- 32. Транспорт жирных кислот в митохондрии из цитоплазмы осуществляет карнитин. Ацил-КоА Карнитин Ацилкарнитин (в цитоплазме) Карнитинацил- трансфераза
- 33. Ацилкарнитин (в цитоплазме) Ацил-КоА Карнитин (в митохондриях) Карнитинацил- трансфераза
- 34. - первая стадия дегидрирования, - стадия гидратации, - вторая стадия дегидрирования, - тиолазная реакция. β-ОКИСЛЕНИЕ ЖИРНЫХ
- 35. Первая стадия дегидрирования + ФАД Ацил-КоА ДГ + ФАДН2 Ацил-КоА Еноил-КоА
- 36. Стадия гидратации Еноил-КоА- гидратаза β − Оксиацил-КоА Еноил-КоА
- 37. Вторая стадия дегидрирования + НАД β-гидроксиацил-КоА- дегидрогеназа + НАДН+ + Н+ β-кетоацил-КоА β-оксиацил-КоА
- 38. Тиолазная реакция β-кетоацил-КоА + HS-KoA Тиолаза + Ацил-КоА вновь проходит путь β-окисления вплоть до образования бутирил-КоА,
- 40. Баланс β-окисления пальмитиновой кислоты (С16) При окислении жирной кислоты, содержащей n углеродных атомов получается n/2 ацетил-КоА,
- 41. Баланс β-окисления
- 42. β-окисление жирных кислот с нечётным числом атомов углерода В конечном итоге образуются ацетил-КоА и пропионил-КоА. Пропионил-КоА
- 43. β-окисление ненасыщенных жирных кислот Наличие дополнительных ферментов изомеразы и эпимеразы обеспечивает возможность полного окисления всех ненасыщенных
- 44. β-окисление ненасыщенных жирных кислот
- 45. Окисление жирных кислот протекает в печени, мышцах, жировой ткани. Мышцы, миокард, печень активно используют жирные кислоты
- 46. Регуляция β−окисления Регуляторный фермент – карнитинацилтрансфераза. Чем интенсивнее идёт распад АТФ, тем быстрее окисляются жирные кислоты.
- 47. Аллостерическая регуляция метаболизма жирных кислот в печени
- 48. Биосинтез липидов идёт в жировой ткани, печени, почках, нервной ткани.
- 49. Биосинтез липидов зависит от распада глюкозы АТФ, НАДФН2, ацетил-КоА.
- 50. Пути образования и использования Ацетил-КоА Стероидные гормоны.
- 51. Биосинтез жирных кислот идёт в цитоплазме, нужен ацетил-КоА из митохондрий, участвует малонил-КоА, происходит перенос ацетил-КоА в
- 52. Перенос ацетил-КоА из митохондрий в цитоплазму Ацетил-КоА не проникает через мембрану митохондрий в цитоплазму. Ацетил-КоА +
- 53. Реакции синтеза жирных кислот СН3-CО-S-KoA + биотин-СО2 + АТФ НООС-СН2-CО-S-KoA + АДФ + Фн малонил-КоА Ацетил-КоА-карбоксилаза
- 54. Ацетил-КоА-карбоксилаза аллостерический фермент, активатором является цитрат, повышение содержания цитрата в митохондриях приводит к тому, что при
- 55. Далее действует мультиферментный комплекс – синтетаза жирных кислот, который представляет собой 6 ферментов, связанных с АПБ.
- 56. Роль простетической группы в АПБ играет 4-фосфопантетеин. Это подвижная «рука», переносящая остатки жирных кислот от активного
- 58. СН3-CО-S-KoA + HS-АПБ HS-КоА + СН3-CО-S-АПБ ацетил-АПБ АПБ-ацетилтрансфераза
- 59. НООС-СН2-CО-S-KoA + HS-АПБ (малонил-КоА) АПБ-малонилтрансфераза HS-КоА + НООС-СН2-CО-S-АПБ малонил-АПБ
- 60. Далее происходит конденсация малонил-АПБ и ацетил-АПБ. СН3-CО-S-АПБ + НООС-СН2-CО-S-АПБ ацетил-АПБ малонил-АПБ СН3-CО-СН2-CО-S-АПБ + СO2 ацетоацетил-АПБ β-Кетоацил-
- 61. СН3-CО-СН2-CО-S-АПБ + НАДФН+Н+ ацетоацетил-АПБ СН3-CH(ОH)-СН2-CО-S-АПБ+НАДФ+ β-оксибутирил-АПБ β-Кетоацил- АПБ-редуктаза
- 62. СН3-CH(ОH)-СН2-CО-S-АПБ β-оксибутирил-АПБ СН3-CH=СН-CО-S-АПБ + Н2О кротонил-АПБ β-оксиацил- АПБ-дегидратаза
- 63. СН3-CH=СН-CО-S-АПБ + НАДФН+Н+ кротонил-АПБ СН3-CH2-СН2-CО-S-АПБ +НАДФ+ бутирил-АПБ еноилАПБ -редуктаза
- 64. Далее цикл повторяется. Малонил-КоА переносится на SH-группу фосфопантетеина АПБ. Бутирил-АПБ + малонил-АПБ кетокапронилАПБ + СО2 Пальмитиновая
- 66. Источники НАДФН+Н на 50% -пентозный цикл, изоцитратдегидрогеназная реакция, малик-реакция. Малат СО2 + ПВК МДГ декарбоксилирующая НАДФН+Н+
- 67. Полиненасыщенные жирные кислоты Линолевая, линоленовая жирные кислоты в организме не синтезируются. Арахидоновая кислота синтезируется из линолевой,
- 68. Гормональная регуляция обмена жирных кислот
- 69. Биосинтез ТАГ В почках, стенке кишечника, печени высока активность глицеролкиназы. +АТФ глицеролкиназа Глицерол-3-фосфат +АДФ Mg2+
- 70. В мышцах, жировой ткани активность глицеролкиназы низкая и образование глицерол-3-фосфата связано с гликолизом и гликогенолизом. Глицерол-3-фосфат
- 71. Глицерол-3-фосфат глицеролфосфатацил трансфераза 2 Фосфатидная кислота
- 72. Фосфатидная кислота фосфатидатфосфогидролаза H3PO4 1,2-диглицерид
- 73. 1,2-диглицерид Жиры, синтезированные в жировой ткани, там и откладываются, а из печени жиры транспортируются в составе
- 74. Показатели липидного обмена в крови здорового человека Общие липиды 4-10 г/л Триглицериды 1-1,5 г/л Фосфолипиды 2-2,5
- 76. Скачать презентацию

Обмен триацилглицеринов
С пищей в сутки поступает 70 г ТГ.
Эндогенный синтез ТГ
Обмен триацилглицеринов
С пищей в сутки поступает 70 г ТГ.
Эндогенный синтез ТГ

Обмен липидов
Обмен липидов

Функции резервных жиров
резервуар энергии,
теплоизоляционная,
защита от механических травм.
Функции резервных жиров
резервуар энергии,
теплоизоляционная,
защита от механических травм.

Транспорт ТГ осуществляют
ХМ,
ЛПОНП.
Транспорт ТГ осуществляют
ХМ,
ЛПОНП.

Липопротеидлипаза
осуществляет гидролиз ТГ.
В жировой ткани жир накапливается за счёт
поступления
Липопротеидлипаза
осуществляет гидролиз ТГ.
В жировой ткани жир накапливается за счёт
поступления
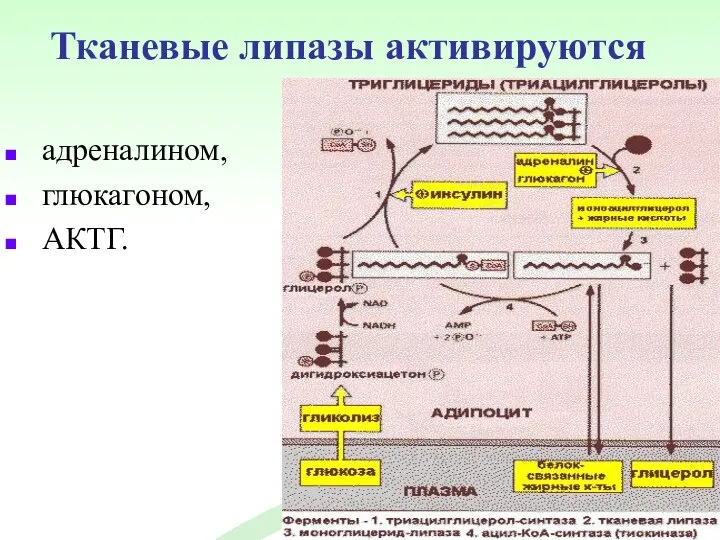
Тканевые липазы активируются
адреналином,
глюкагоном,
АКТГ.
Тканевые липазы активируются
адреналином,
глюкагоном,
АКТГ.

Мобилизация депонированных жиров
происходит путём их гидролиза до жирных кислот и
Мобилизация депонированных жиров
происходит путём их гидролиза до жирных кислот и

Окисление глицерина
Глицеролкиназа
АТФ
АДФ
Mg2+
Глицерин
Глицерол-3-фосфат
Окисление глицерина
Глицеролкиназа
АТФ
АДФ
Mg2+
Глицерин
Глицерол-3-фосфат

Глицеролфосфат-
дегидрогеназа
НАД
НАДН+Н+
Глицерол-3-фосфат
Диоксиацетонфосфат
Глицеролфосфат-
дегидрогеназа
НАД
НАДН+Н+
Глицерол-3-фосфат
Диоксиацетонфосфат

CO2 + H2O + E
Лактат
Анаэробный путь
Аэробный путь
ФГА
CO2 + H2O + E
Лактат
Анаэробный путь
Аэробный путь
ФГА

Диоксиацетон
Триозофосфат-
изомераза
Фосфоглицериновый
альдегид
Диоксиацетон
Триозофосфат-
изомераза
Фосфоглицериновый
альдегид

+ НАД+ + H3PO4
Фосфоглицериновый
альдегид
+ НАДН+Н+
Глицеральдегидфосфат
дегидрогеназа
1,3-дифосфоглицерат
+ НАД+ + H3PO4
Фосфоглицериновый
альдегид
+ НАДН+Н+
Глицеральдегидфосфат
дегидрогеназа
1,3-дифосфоглицерат

Фосфоглицераткиназа
1,3-дифосфоглицерат
3-фосфоглицерат
АДФ
АТФ
Mg2+
Фосфоглицераткиназа
1,3-дифосфоглицерат
3-фосфоглицерат
АДФ
АТФ
Mg2+

3-фосфоглицерат
Фосфоглицеро
мутаза
2-фосфоглицерат
3-фосфоглицерат
Фосфоглицеро
мутаза
2-фосфоглицерат

2-фосфоглицерат
Енолаза
Н2О
Mg2+
Фосфоенолпируват
2-фосфоглицерат
Енолаза
Н2О
Mg2+
Фосфоенолпируват

Фосфоенолпируват
Пируваткиназа
Пируват (ПВК)
АДФ
АТФ
Mg2+
Фосфоенолпируват
Пируваткиназа
Пируват (ПВК)
АДФ
АТФ
Mg2+

Пируват (ПВК)
Ацетил- КоА
Цикл Кребса
Пируват (ПВК)
Ацетил- КоА
Цикл Кребса

Цикл Кребса
Цитрат-синтаза
Аконитат-
гидратаза
Аконитат-
гидратаза
Изоцитрат-
дегидрогеназа
α-Кетоглутарат дегидрогеназный
комплекс
Сукцинил-КоА
-синтетаза
Сукцинат-
дегидрогеназа
Фумараза
Малат-
дегидрогеназа
Цикл Кребса
Цитрат-синтаза
Аконитат-
гидратаза
Аконитат-
гидратаза
Изоцитрат-
дегидрогеназа
α-Кетоглутарат дегидрогеназный
комплекс
Сукцинил-КоА
-синтетаза
Сукцинат-
дегидрогеназа
Фумараза
Малат-
дегидрогеназа

CH2-COOH
|
HOC-COOH
|
H2C-COOH
Цитрат
CH2-COOH
|
HOC-COOH
|
H2C-COOH
Цитрат

COOH
|
CH2
|
HO-C-COOH
|
H-C-H
COOH
|
CH2
|
HO-C-COOH
|
H-C-H

COOH
|
CH2
|
H-C-COOH
|
HO-C-H
|
COOH
|
CH2
|
H-C-COOH
|
HO-C-H
|

COOH
|
CH2
|
CH2
|
C=O
COOH
|
CH2
|
CH2
|
C=O

COOH
|
CH2
|
СН2
|
C=O
COOH
|
CH2
|
СН2
|
C=O

COOH
|
CH2
|
СН2
|
COOH
COOH
|
CH2
|
СН2
|
COOH

COOH
|
CH
||
HС
|
COOH
Фумарат
COOH
|
CH
||
HС
|
COOH
Фумарат
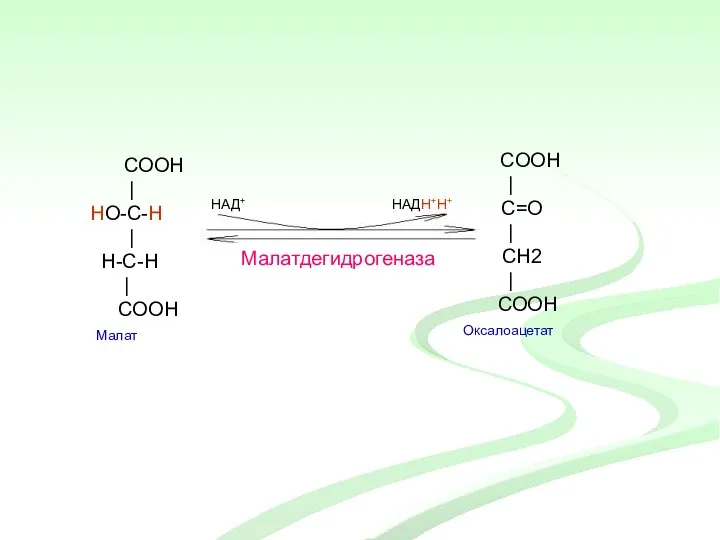
COOH
|
HO-C-H
|
H-С-H
|
COOH
COOH
|
HO-C-H
|
H-С-H
|
COOH

Баланс аэробного распада глицерина
От глицерина до ФГА
затрата – 1 АТФ
Баланс аэробного распада глицерина
От глицерина до ФГА
затрата – 1 АТФ

ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
аэробный процесс.
В катаболизме жирных кислот выделяют 3 части:
β-окисление,
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
аэробный процесс.
В катаболизме жирных кислот выделяют 3 части:
β-окисление,

Кнооп установил, что окисление идёт в β-положении.
Ленинджер и Кеннеди установили, что
Кнооп установил, что окисление идёт в β-положении.
Ленинджер и Кеннеди установили, что

Активация жирных кислот происходит на наружной поверхности мембраны митохондрий.
R-COOH + HS-KoA
Активация жирных кислот происходит на наружной поверхности мембраны митохондрий.
R-COOH + HS-KoA

Транспорт жирных кислот в митохондрии из цитоплазмы осуществляет карнитин.
Ацил-КоА
Карнитин
Ацилкарнитин
(в цитоплазме)
Карнитинацил-
трансфераза
Транспорт жирных кислот в митохондрии из цитоплазмы осуществляет карнитин.
Ацил-КоА
Карнитин
Ацилкарнитин
(в цитоплазме)
Карнитинацил-
трансфераза

Ацилкарнитин
(в цитоплазме)
Ацил-КоА
Карнитин
(в митохондриях)
Карнитинацил-
трансфераза
Ацилкарнитин
(в цитоплазме)
Ацил-КоА
Карнитин
(в митохондриях)
Карнитинацил-
трансфераза

- первая стадия дегидрирования,
- стадия гидратации,
- вторая стадия дегидрирования,
- стадия гидратации,
- вторая стадия дегидрирования,

Первая стадия дегидрирования
+ ФАД
Ацил-КоА ДГ
+ ФАДН2
Ацил-КоА
Еноил-КоА
Первая стадия дегидрирования
+ ФАД
Ацил-КоА ДГ
+ ФАДН2
Ацил-КоА
Еноил-КоА

Стадия гидратации
Еноил-КоА-
гидратаза
β − Оксиацил-КоА
Еноил-КоА
Стадия гидратации
Еноил-КоА-
гидратаза
β − Оксиацил-КоА
Еноил-КоА

Вторая стадия дегидрирования
+ НАД
β-гидроксиацил-КоА-
дегидрогеназа
+ НАДН+ + Н+
β-кетоацил-КоА
β-оксиацил-КоА
Вторая стадия дегидрирования
+ НАД
β-гидроксиацил-КоА-
дегидрогеназа
+ НАДН+ + Н+
β-кетоацил-КоА
β-оксиацил-КоА

Тиолазная реакция
β-кетоацил-КоА
+ HS-KoA
Тиолаза
+
Ацил-КоА вновь проходит путь β-окисления
вплоть до
Тиолазная реакция
β-кетоацил-КоА
+ HS-KoA
Тиолаза
+
Ацил-КоА вновь проходит путь β-окисления
вплоть до


Баланс β-окисления пальмитиновой кислоты (С16)
При окислении жирной кислоты, содержащей n углеродных
Баланс β-окисления пальмитиновой кислоты (С16)
При окислении жирной кислоты, содержащей n углеродных

Баланс β-окисления
Баланс β-окисления
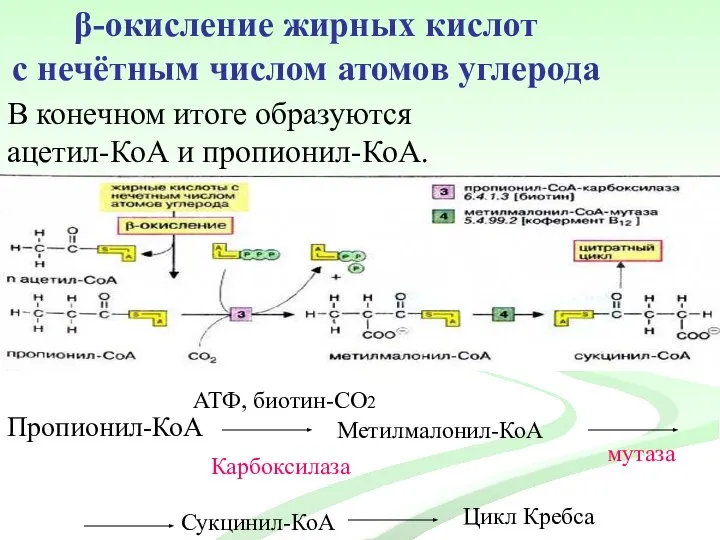
β-окисление жирных кислот
с нечётным числом атомов углерода
В конечном итоге образуются
ацетил-КоА
β-окисление жирных кислот
с нечётным числом атомов углерода
В конечном итоге образуются
ацетил-КоА

β-окисление ненасыщенных жирных кислот
Наличие дополнительных ферментов изомеразы и эпимеразы обеспечивает возможность
β-окисление ненасыщенных жирных кислот
Наличие дополнительных ферментов изомеразы и эпимеразы обеспечивает возможность

β-окисление
ненасыщенных
жирных кислот
β-окисление
ненасыщенных
жирных кислот
Окисление жирных кислот протекает в
печени,
мышцах,
жировой ткани.
Мышцы, миокард,
Окисление жирных кислот протекает в
печени,
мышцах,
жировой ткани.
Мышцы, миокард,

Регуляция β−окисления
Регуляторный фермент – карнитинацилтрансфераза.
Чем интенсивнее идёт распад АТФ, тем
Регуляция β−окисления
Регуляторный фермент – карнитинацилтрансфераза.
Чем интенсивнее идёт распад АТФ, тем

Аллостерическая регуляция метаболизма жирных кислот в печени
Аллостерическая регуляция метаболизма жирных кислот в печени
Биосинтез липидов идёт в
жировой ткани,
печени,
почках,
нервной ткани.
Биосинтез липидов идёт в
жировой ткани,
печени,
почках,
нервной ткани.

Биосинтез липидов зависит от распада глюкозы
АТФ,
НАДФН2,
ацетил-КоА.
Биосинтез липидов зависит от распада глюкозы
АТФ,
НАДФН2,
ацетил-КоА.

Пути образования и использования Ацетил-КоА
Стероидные
гормоны.
Пути образования и использования Ацетил-КоА
Стероидные
гормоны.

Биосинтез жирных кислот
идёт в цитоплазме,
нужен ацетил-КоА из митохондрий,
Биосинтез жирных кислот
идёт в цитоплазме,
нужен ацетил-КоА из митохондрий,

Перенос ацетил-КоА из митохондрий в цитоплазму
Ацетил-КоА не проникает через мембрану
Перенос ацетил-КоА из митохондрий в цитоплазму
Ацетил-КоА не проникает через мембрану

Реакции синтеза жирных кислот
СН3-CО-S-KoA + биотин-СО2 + АТФ
НООС-СН2-CО-S-KoA + АДФ +
Реакции синтеза жирных кислот
СН3-CО-S-KoA + биотин-СО2 + АТФ
НООС-СН2-CО-S-KoA + АДФ +

Ацетил-КоА-карбоксилаза
аллостерический фермент, активатором является цитрат,
повышение содержания цитрата в митохондриях
Ацетил-КоА-карбоксилаза
аллостерический фермент, активатором является цитрат,
повышение содержания цитрата в митохондриях

Далее действует мультиферментный комплекс – синтетаза жирных кислот, который представляет собой
Далее действует мультиферментный комплекс – синтетаза жирных кислот, который представляет собой

Роль простетической группы в АПБ играет 4-фосфопантетеин. Это подвижная «рука»,
Роль простетической группы в АПБ играет 4-фосфопантетеин. Это подвижная «рука»,


СН3-CО-S-KoA + HS-АПБ
HS-КоА + СН3-CО-S-АПБ
ацетил-АПБ
АПБ-ацетилтрансфераза
СН3-CО-S-KoA + HS-АПБ
HS-КоА + СН3-CО-S-АПБ
ацетил-АПБ
АПБ-ацетилтрансфераза
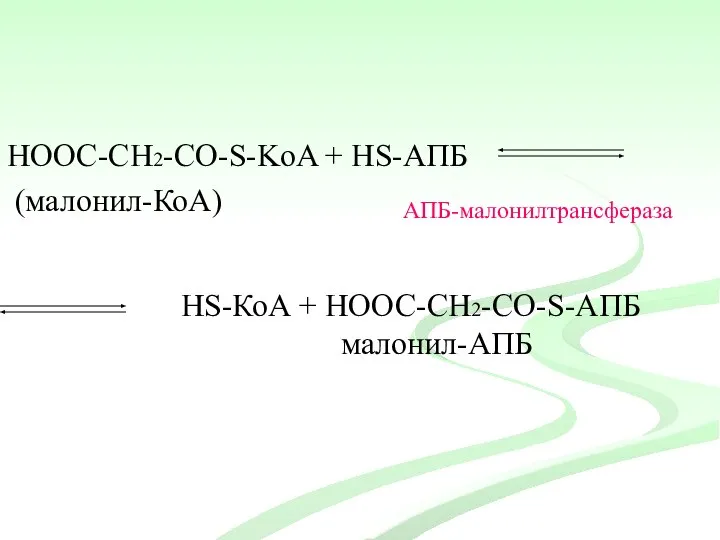
НООС-СН2-CО-S-KoA + HS-АПБ
(малонил-КоА)
АПБ-малонилтрансфераза
HS-КоА + НООС-СН2-CО-S-АПБ
малонил-АПБ
НООС-СН2-CО-S-KoA + HS-АПБ
(малонил-КоА)
АПБ-малонилтрансфераза
HS-КоА + НООС-СН2-CО-S-АПБ
малонил-АПБ

Далее происходит конденсация малонил-АПБ и ацетил-АПБ.
СН3-CО-S-АПБ + НООС-СН2-CО-S-АПБ
ацетил-АПБ малонил-АПБ
Далее происходит конденсация малонил-АПБ и ацетил-АПБ.
СН3-CО-S-АПБ + НООС-СН2-CО-S-АПБ
ацетил-АПБ малонил-АПБ

СН3-CО-СН2-CО-S-АПБ + НАДФН+Н+
ацетоацетил-АПБ
СН3-CH(ОH)-СН2-CО-S-АПБ+НАДФ+
β-оксибутирил-АПБ
β-Кетоацил-
АПБ-редуктаза
СН3-CО-СН2-CО-S-АПБ + НАДФН+Н+
ацетоацетил-АПБ
СН3-CH(ОH)-СН2-CО-S-АПБ+НАДФ+
β-оксибутирил-АПБ
β-Кетоацил-
АПБ-редуктаза
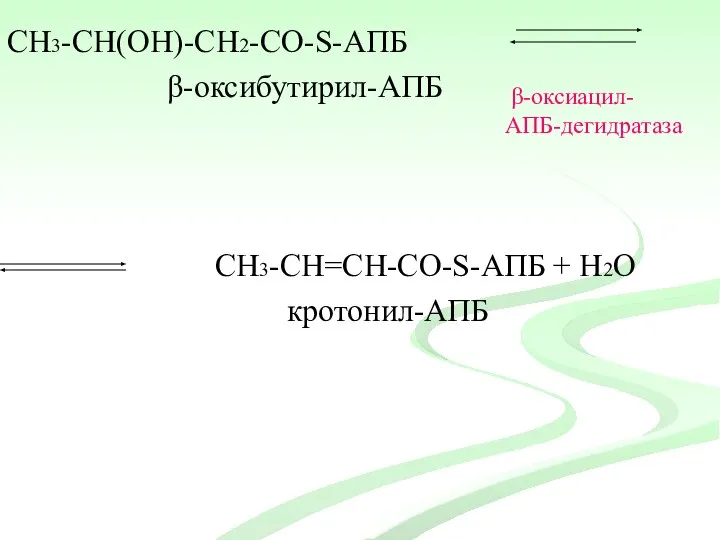
СН3-CH(ОH)-СН2-CО-S-АПБ
β-оксибутирил-АПБ
СН3-CH=СН-CО-S-АПБ + Н2О
кротонил-АПБ
β-оксиацил-
АПБ-дегидратаза
СН3-CH(ОH)-СН2-CО-S-АПБ
β-оксибутирил-АПБ
СН3-CH=СН-CО-S-АПБ + Н2О
кротонил-АПБ
β-оксиацил-
АПБ-дегидратаза

СН3-CH=СН-CО-S-АПБ + НАДФН+Н+
кротонил-АПБ
СН3-CH2-СН2-CО-S-АПБ +НАДФ+
бутирил-АПБ
еноилАПБ
-редуктаза
СН3-CH=СН-CО-S-АПБ + НАДФН+Н+
кротонил-АПБ
СН3-CH2-СН2-CО-S-АПБ +НАДФ+
бутирил-АПБ
еноилАПБ
-редуктаза

Далее цикл повторяется.
Малонил-КоА переносится на SH-группу фосфопантетеина АПБ.
Бутирил-АПБ + малонил-АПБ
Далее цикл повторяется.
Малонил-КоА переносится на SH-группу фосфопантетеина АПБ.
Бутирил-АПБ + малонил-АПБ


Источники НАДФН+Н
на 50% -пентозный цикл,
изоцитратдегидрогеназная реакция,
малик-реакция.
Малат СО2 +
Источники НАДФН+Н
на 50% -пентозный цикл,
изоцитратдегидрогеназная реакция,
малик-реакция.
Малат СО2 +

Полиненасыщенные жирные кислоты
Линолевая, линоленовая жирные кислоты в организме не синтезируются.
Арахидоновая
Полиненасыщенные жирные кислоты
Линолевая, линоленовая жирные кислоты в организме не синтезируются.
Арахидоновая

Гормональная регуляция обмена жирных кислот
Гормональная регуляция обмена жирных кислот

Биосинтез ТАГ
В почках, стенке кишечника, печени высока активность глицеролкиназы.
+АТФ
глицеролкиназа
Глицерол-3-фосфат
+АДФ
Mg2+
Биосинтез ТАГ
В почках, стенке кишечника, печени высока активность глицеролкиназы.
+АТФ
глицеролкиназа
Глицерол-3-фосфат
+АДФ
Mg2+
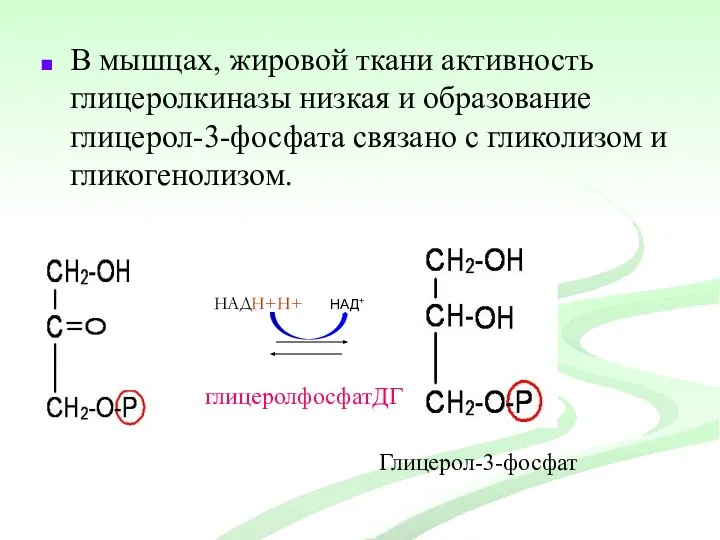
В мышцах, жировой ткани активность глицеролкиназы низкая и образование глицерол-3-фосфата связано
В мышцах, жировой ткани активность глицеролкиназы низкая и образование глицерол-3-фосфата связано

Глицерол-3-фосфат
глицеролфосфатацил
трансфераза
2
Фосфатидная
кислота
Глицерол-3-фосфат
глицеролфосфатацил
трансфераза
2
Фосфатидная
кислота

Фосфатидная
кислота
фосфатидатфосфогидролаза
H3PO4
1,2-диглицерид
Фосфатидная
кислота
фосфатидатфосфогидролаза
H3PO4
1,2-диглицерид

1,2-диглицерид
Жиры, синтезированные в жировой ткани, там и откладываются, а
1,2-диглицерид
Жиры, синтезированные в жировой ткани, там и откладываются, а

Показатели липидного обмена в крови здорового человека
Общие липиды 4-10 г/л
Триглицериды 1-1,5
Показатели липидного обмена в крови здорового человека
Общие липиды 4-10 г/л
Триглицериды 1-1,5
 Экстремизм и терроризм: основные понятия и причины его возникновения
Экстремизм и терроризм: основные понятия и причины его возникновения Интерференция света
Интерференция света Рулевое управление. Способы поворота автомобиля
Рулевое управление. Способы поворота автомобиля Сборка квадрокоптера
Сборка квадрокоптера Культура первобытного общества
Культура первобытного общества Геополитика. Современная геополитическая картина мира
Геополитика. Современная геополитическая картина мира Презентация на тему "Мониторинг физической подготовленности среди 6-х классов" - скачать презентации по Педагогике
Презентация на тему "Мониторинг физической подготовленности среди 6-х классов" - скачать презентации по Педагогике Презентация ОСО
Презентация ОСО Dziesięć Bożych przykazań
Dziesięć Bożych przykazań Требования к отчету
Требования к отчету Антон Павлович Чехов (1860 - 1904)
Антон Павлович Чехов (1860 - 1904) Благоустройство и озеленение сквера имени Дзержинского
Благоустройство и озеленение сквера имени Дзержинского Достопримечательности мира
Достопримечательности мира Координатная плоскость. Урок-лекция 6 класс.
Координатная плоскость. Урок-лекция 6 класс. Презентация Сравнительная характеристика стран зарубежной Азии: Индии и Израиля
Презентация Сравнительная характеристика стран зарубежной Азии: Индии и Израиля Монголия
Монголия  Программирование на Delphi
Программирование на Delphi  Патофизиология поджелудочной железы
Патофизиология поджелудочной железы Стандарт как элемент промышленной политики
Стандарт как элемент промышленной политики Советские деньги
Советские деньги Медицинская протозология
Медицинская протозология Алгоритмические языки и программирование. ctype.h
Алгоритмические языки и программирование. ctype.h Технология изготовления тряпичных кукол
Технология изготовления тряпичных кукол Комплекс испытательных средств для исследования ползучести и состава газообразных продуктов деления
Комплекс испытательных средств для исследования ползучести и состава газообразных продуктов деления Сила Трения
Сила Трения ТРУДОВОЕ ПРАВО
ТРУДОВОЕ ПРАВО Основы религиозных культур и светской этики
Основы религиозных культур и светской этики Футбольные площадки (Киев)
Футбольные площадки (Киев)