-
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА

Содержание
- 2. Если Вы НЕ ЗНАЕТЕ: 1. Что такое репликация транскрипция процессинг трансляция 2. Из чего состоит ДНК
- 3. УРОВНИ ТОНКОЙ СТРУКТУРНОЙ ОРГАНИЗАЦИИ ХРОМАТИНА СПб., 2013 http://www.cytspb.rssi.ru/education/chikhirzhina_structure_chromatine_2013.pdf КОНИЧЕВ А. С. Молекулярная биология. М.: Академия, 2012
- 4. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ. Словарик
- 5. Словарь Хроматин – это вещество хромосом – комплекс ДНК и белков Нуклеиновые кислоты – это линейные
- 6. 1868 г. Швейцарский патологоанатом Иоган Фридрих Мишер выделил из клеточного ядра новое вещество, которое он назвал
- 7. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ. Строение нуклеиновых кислот
- 8. Компоненты нуклеиновых кислот структурными единицами НК являются нуклеотиды, которые состоят из гетероциклического азотистого основания, моносахарида (пентозы)
- 9. Номенклатура нуклеотидов Пуриновый и пиримидиновый нуклеотиды
- 10. Нуклеотиды – фосфорные эфиры нуклеозидов. В зависимости от числа остатков фосфорной кислоты различают: нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты
- 11. Строение нуклеоидов. Азотистые основания производные пиримидинового и пуринового гетероциклов (нумерация атомов в основаниях записывается внутри цикла)
- 12. Строение нуклеотидов. Сахара. представлены рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Номера атомов записывают
- 13. Значение нуклеотидов
- 14. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ. Обмен нуклеотидов
- 15. Обмен нуклеотидов Пищевые пурины и пиримидины мало используются для синтеза нуклеиновых кислот. до 90% пуриновых и
- 16. Биосинтез пуриновых нуклеотидов Образование 5-фосфорибозил-1-дифосфата. Происхождение атомов С и N в пуриновом кольце.
- 17. Синтез АМФ и ГМФ из ИМФ. 1. аденилосукцинатсинтетаза 2. аденилосукциназа 3. ИМФ-дегидрогеназа 4. ГМФ-синтетаза.
- 18. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ. Структура ДНК
- 19. Первичная структура Уникальность структуры и функциональная индивидуальность молекул НК определяются их первичной структурой. Первичная структура ДНК
- 20. Связи в одноцепочечной ДНК Фосфодиэфирная связь 5’-группа атома одного нуклеотида связана с 3’-группой дезоксирибозы соседнего нуклеотида.
- 21. Вторичная структура 1953 г. Дж. Уотсон и Ф. Крик предложили модель пространственной структуры ДНК. Согласно этой
- 22. ВОДОРОДНАЯ СВЯЗЬ Образуется между комплементарными парами оснований противоположный цепей. Пара А-Т стабилизируется двумя водородными связями, Г-Ц
- 24. Цепи ДНК образуют 2 желоба - малую и большую борозды.
- 25. Характеристики вторичной структуры ДНК Нерегулярность Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование
- 26. Антипараллельность ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой. Характеристики
- 27. Комплементарность Каждому азотистому основанию одной цепи соответствует строго определенное основание другой цепи. Соответствие задается стереохимией. Характеристики
- 28. Регулярная вторичная структура Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью Характеристики
- 29. В зависимости от рН среды, ионной силы, концентрации воды и т.п. конфигурация двойной спирали может меняться.
- 30. Формы двойной спирали ДНК Å - Ангстрем (10) -10 м В-ФОРМА на один виток (шаг) спирали
- 31. Формы двойной спирали ДНК -10 Å - Ангстрем (10) м А-ФОРМА 11 пар азотистых оснований на
- 32. Формы двойной спирали ДНК -10 Å - Ангстрем (10) м С-ФОРМА шаг спирали 31Å, 9.3 пар
- 33. Формы двойной спирали ДНК -10 Å - Ангстрем (10) м Z –ФОРМА «ЗИГЗАГ» Высота шага спирали
- 34. Форма SBS (side by side) лишена взаимозакрученности в двойную спираль. Важна для биосинтеза ДНК Формы двойной
- 35. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ. Молекулярная организация ядерного хроматина
- 37. Третичная структура. Уровни компактизации ДНК. НУКЛЕОСОМА – это повторяющийся структурный элемент хроматина, содержащий гистоновый октамер и
- 38. Гистоны – основные белки Выделяют 5 фракций гистонов Образуется около 60 млн. молекул каждой фракции на
- 40. С октамером контактирует 145 п.н. и 20-30-40 п.н. между нуклеосомными корами. (Нуклеосомный уровень упаковки свойственен всей
- 41. Структура нуклеосом
- 42. Негистоновые белки Цинковый палец - фрагмент белка, содержащий около 20 аминокислотных остатков, в котором атом цинка
- 43. Н1 взаимодействует с ОКТАМЕРАМИ, сближает их, на него наматывается ДНК. Образуется СУПЕРБИД. (Происходит сокращение линейного размера
- 44. Негистоновые белки узнают определенные последовательности ДНК и связываются с ними и друг другом, образуя петли по
- 45. Третичная структура. Уровни компактизации ДНК. МЕТАФАЗНАЯ ХРОМОСОМА
- 47. Скачать презентацию

Если Вы НЕ ЗНАЕТЕ:
1. Что такое репликация
транскрипция
процессинг
трансляция
2. Из
Если Вы НЕ ЗНАЕТЕ: 1. Что такое репликация транскрипция процессинг трансляция 2. Из

УРОВНИ ТОНКОЙ СТРУКТУРНОЙ ОРГАНИЗАЦИИ ХРОМАТИНА
СПб., 2013
http://www.cytspb.rssi.ru/education/chikhirzhina_structure_chromatine_2013.pdf
КОНИЧЕВ А. С. Молекулярная биология. М.:
УРОВНИ ТОНКОЙ СТРУКТУРНОЙ ОРГАНИЗАЦИИ ХРОМАТИНА
СПб., 2013
http://www.cytspb.rssi.ru/education/chikhirzhina_structure_chromatine_2013.pdf
КОНИЧЕВ А. С. Молекулярная биология. М.:

МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Словарик
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Словарик

Словарь
Хроматин – это вещество хромосом – комплекс ДНК и белков
Нуклеиновые кислоты
Словарь
Хроматин – это вещество хромосом – комплекс ДНК и белков
Нуклеиновые кислоты

1868 г.
Швейцарский патологоанатом Иоган Фридрих Мишер выделил из клеточного ядра новое
1868 г.
Швейцарский патологоанатом Иоган Фридрих Мишер выделил из клеточного ядра новое

МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Строение нуклеиновых кислот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Строение нуклеиновых кислот
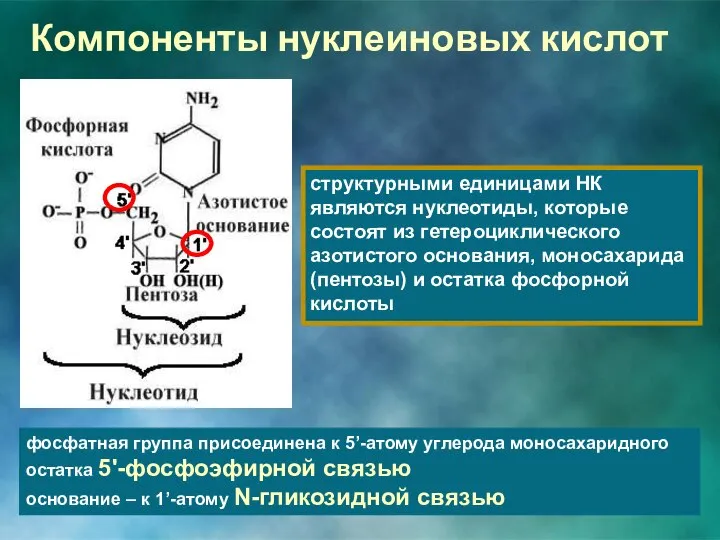
Компоненты нуклеиновых кислот
структурными единицами НК являются нуклеотиды, которые состоят из гетероциклического
Компоненты нуклеиновых кислот
структурными единицами НК являются нуклеотиды, которые состоят из гетероциклического
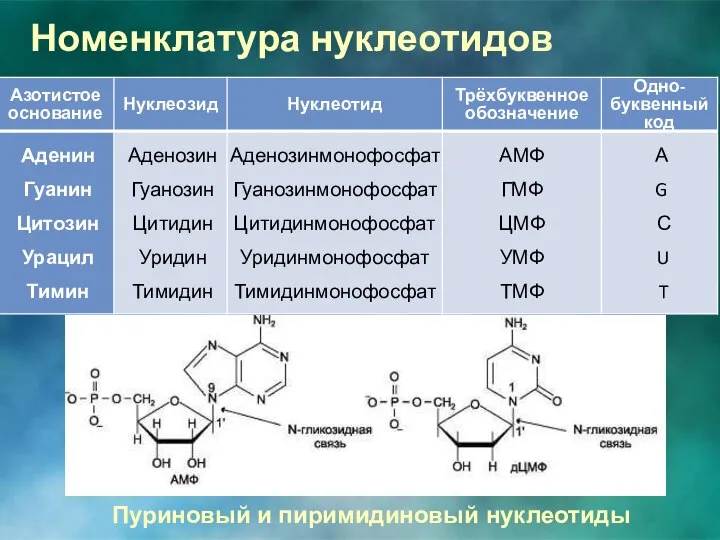
Номенклатура нуклеотидов
Пуриновый и пиримидиновый нуклеотиды
Номенклатура нуклеотидов
Пуриновый и пиримидиновый нуклеотиды
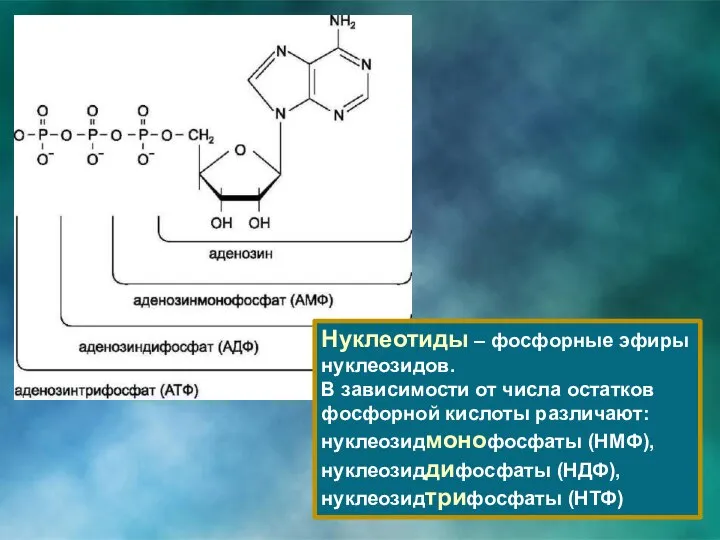
Нуклеотиды – фосфорные эфиры нуклеозидов.
В зависимости от числа остатков фосфорной
Нуклеотиды – фосфорные эфиры нуклеозидов.
В зависимости от числа остатков фосфорной
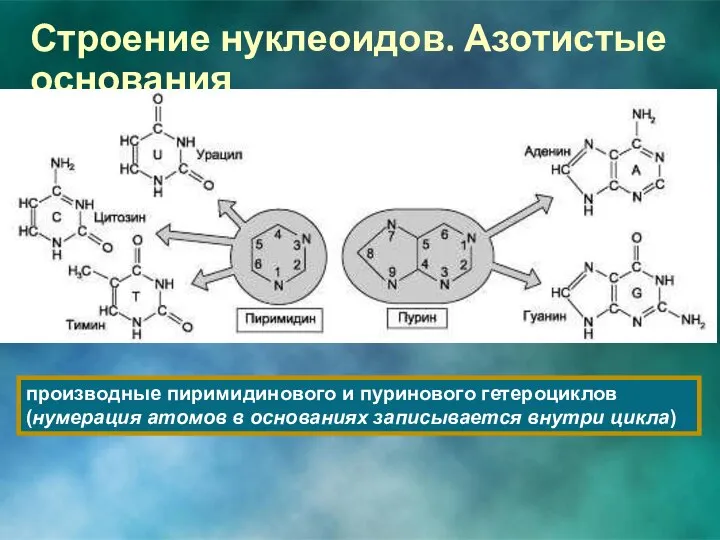
Строение нуклеоидов. Азотистые основания
производные пиримидинового и пуринового гетероциклов (нумерация атомов в
Строение нуклеоидов. Азотистые основания
производные пиримидинового и пуринового гетероциклов (нумерация атомов в
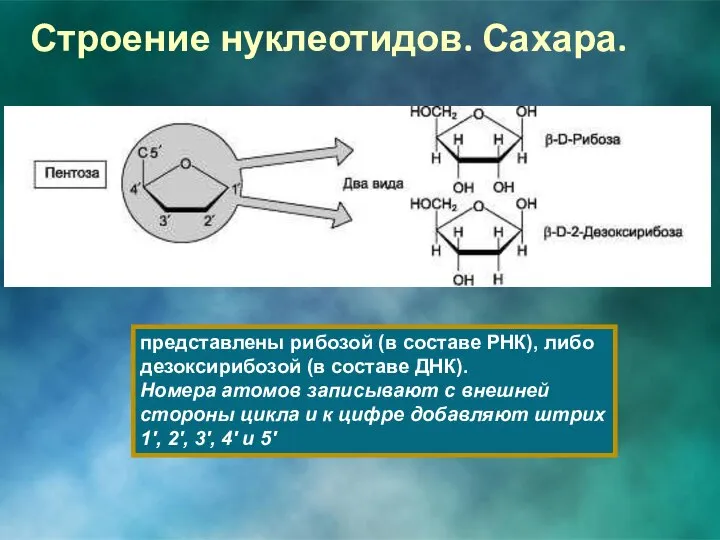
Строение нуклеотидов. Сахара.
представлены рибозой (в составе РНК), либо дезоксирибозой (в составе
Строение нуклеотидов. Сахара.
представлены рибозой (в составе РНК), либо дезоксирибозой (в составе

Значение нуклеотидов
Значение нуклеотидов

МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Обмен нуклеотидов
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Обмен нуклеотидов
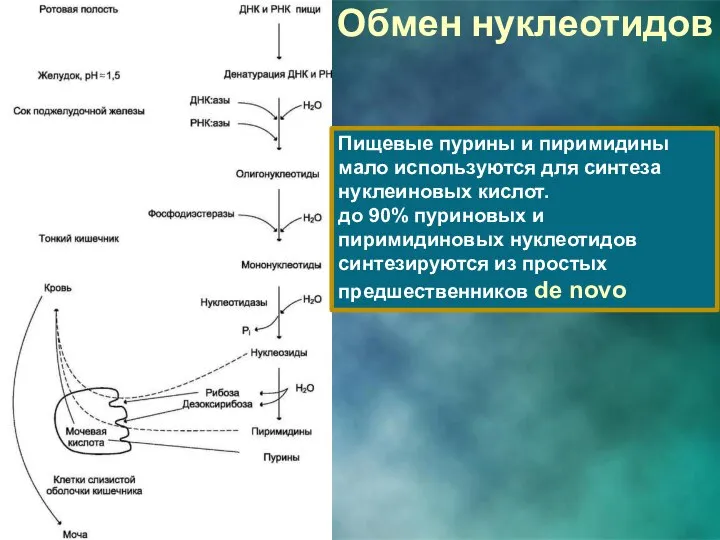
Обмен нуклеотидов
Пищевые пурины и пиримидины мало используются для синтеза нуклеиновых кислот.
Обмен нуклеотидов
Пищевые пурины и пиримидины мало используются для синтеза нуклеиновых кислот.
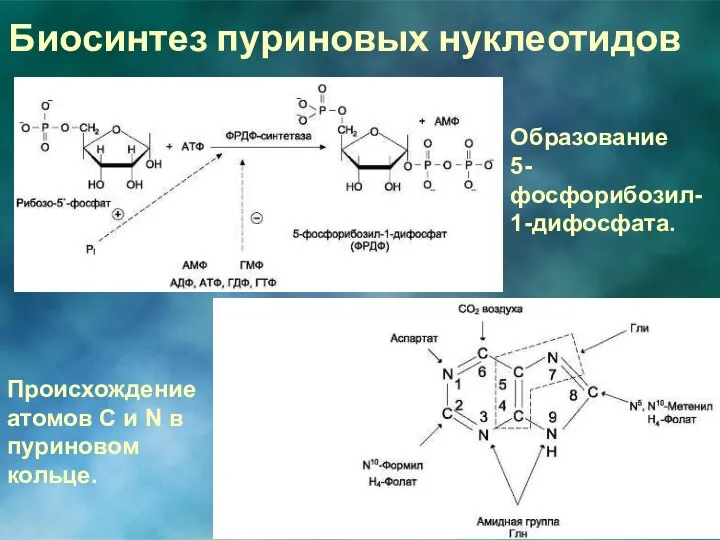
Биосинтез пуриновых нуклеотидов
Образование
5-фосфорибозил-1-дифосфата.
Происхождение атомов С и N в пуриновом кольце.
Биосинтез пуриновых нуклеотидов
Образование
5-фосфорибозил-1-дифосфата.
Происхождение атомов С и N в пуриновом кольце.
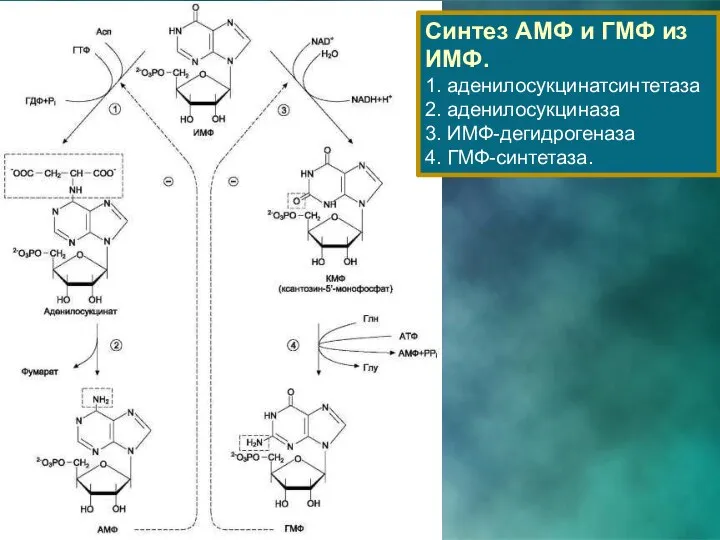
Синтез АМФ и ГМФ из ИМФ.
1. аденилосукцинатсинтетаза 2. аденилосукциназа
3. ИМФ-дегидрогеназа
4.
Синтез АМФ и ГМФ из ИМФ.
1. аденилосукцинатсинтетаза 2. аденилосукциназа
3. ИМФ-дегидрогеназа
4.

МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Структура ДНК
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Структура ДНК
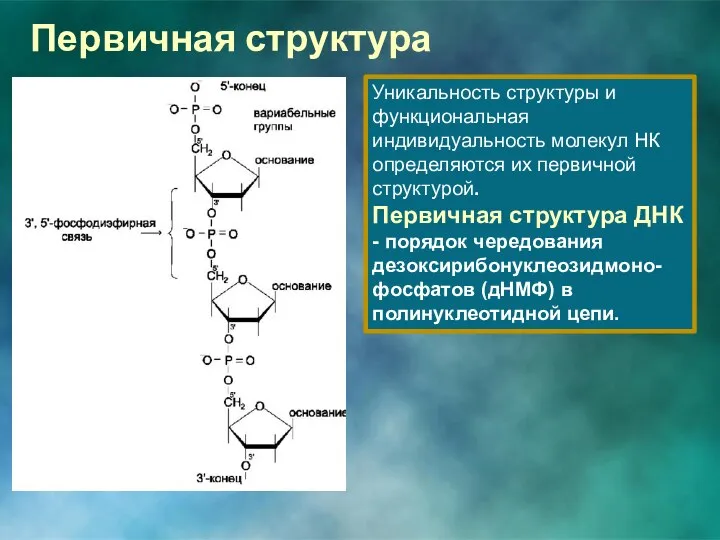
Первичная структура
Уникальность структуры и функциональная индивидуальность молекул НК определяются их первичной
Первичная структура
Уникальность структуры и функциональная индивидуальность молекул НК определяются их первичной
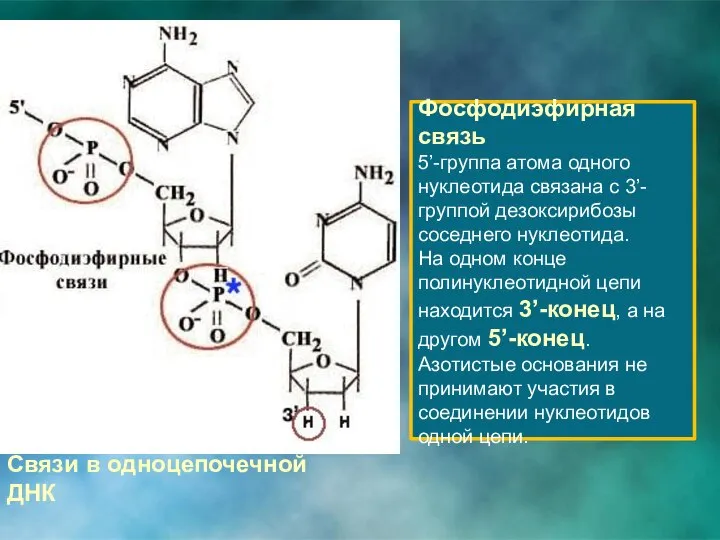
Связи в одноцепочечной ДНК
Фосфодиэфирная связь
5’-группа атома одного нуклеотида связана
Связи в одноцепочечной ДНК
Фосфодиэфирная связь
5’-группа атома одного нуклеотида связана

Вторичная структура
1953 г.
Дж. Уотсон и Ф. Крик
предложили модель пространственной
Вторичная структура
1953 г.
Дж. Уотсон и Ф. Крик
предложили модель пространственной
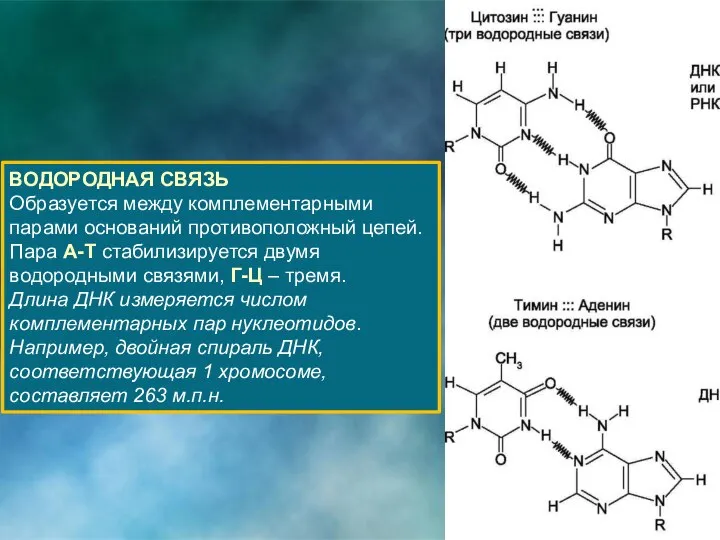
ВОДОРОДНАЯ СВЯЗЬ
Образуется между комплементарными парами оснований противоположный цепей.
Пара А-Т стабилизируется
ВОДОРОДНАЯ СВЯЗЬ
Образуется между комплементарными парами оснований противоположный цепей.
Пара А-Т стабилизируется

Цепи ДНК образуют 2 желоба - малую и большую борозды.
Цепи ДНК образуют 2 желоба - малую и большую борозды.
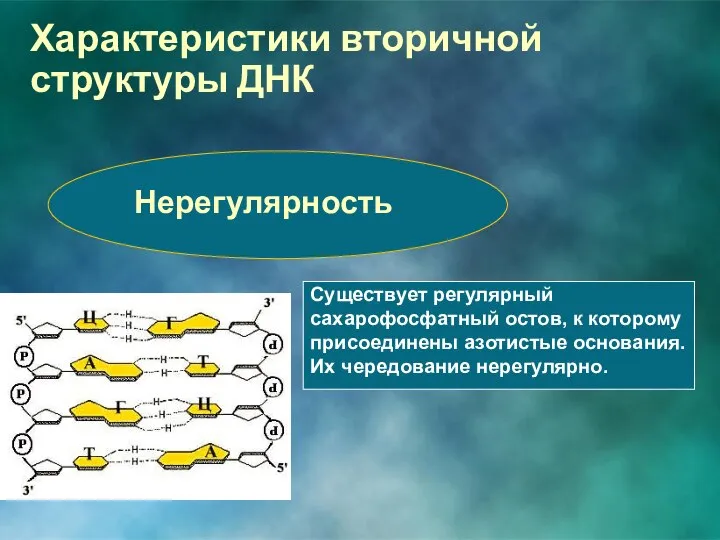
Характеристики вторичной структуры ДНК
Нерегулярность
Существует регулярный сахарофосфатный остов, к которому присоединены
Характеристики вторичной структуры ДНК
Нерегулярность
Существует регулярный сахарофосфатный остов, к которому присоединены
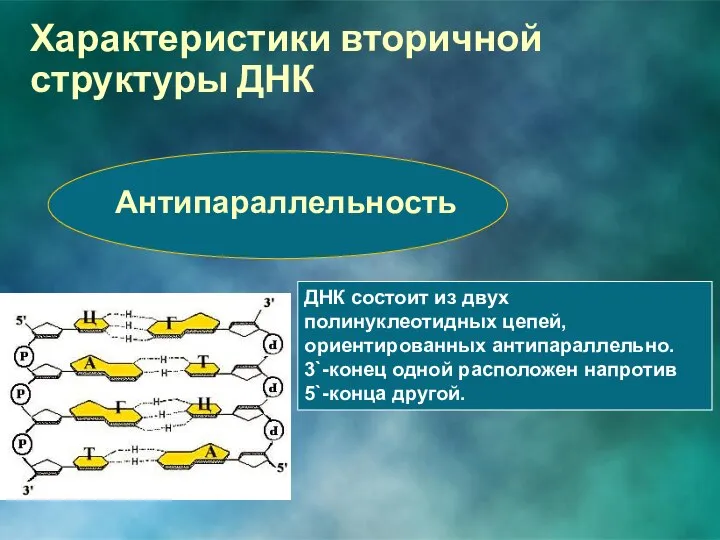
Антипараллельность
ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен
Антипараллельность
ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен
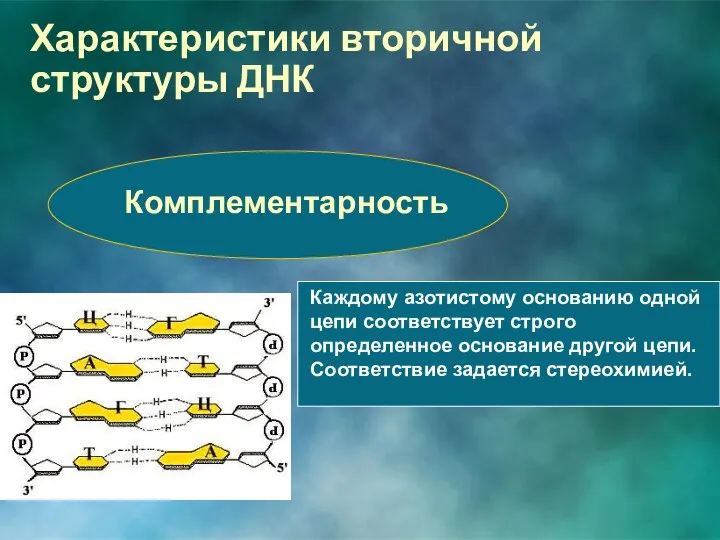
Комплементарность
Каждому азотистому основанию одной цепи соответствует строго определенное основание другой
Комплементарность
Каждому азотистому основанию одной цепи соответствует строго определенное основание другой

Регулярная вторичная структура
Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые
Регулярная вторичная структура
Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые
В зависимости от рН среды, ионной силы, концентрации воды и т.п.
В зависимости от рН среды, ионной силы, концентрации воды и т.п.
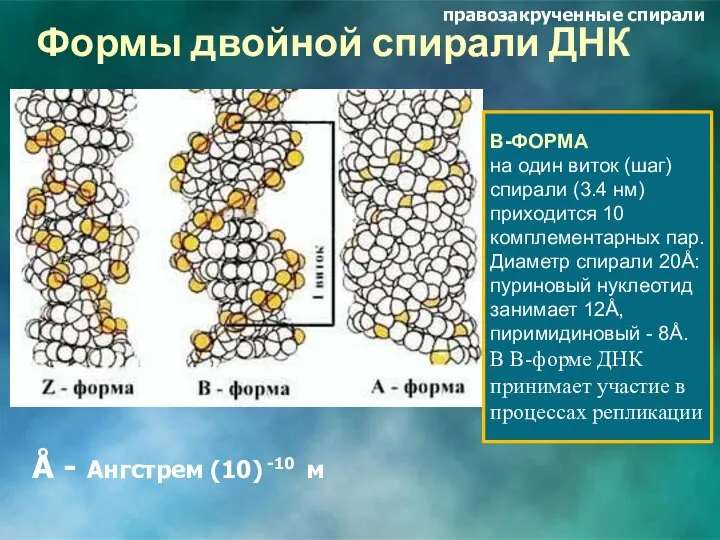
Формы двойной спирали ДНК
Å - Ангстрем (10) -10 м
В-ФОРМА
на
Формы двойной спирали ДНК
Å - Ангстрем (10) -10 м
В-ФОРМА
на
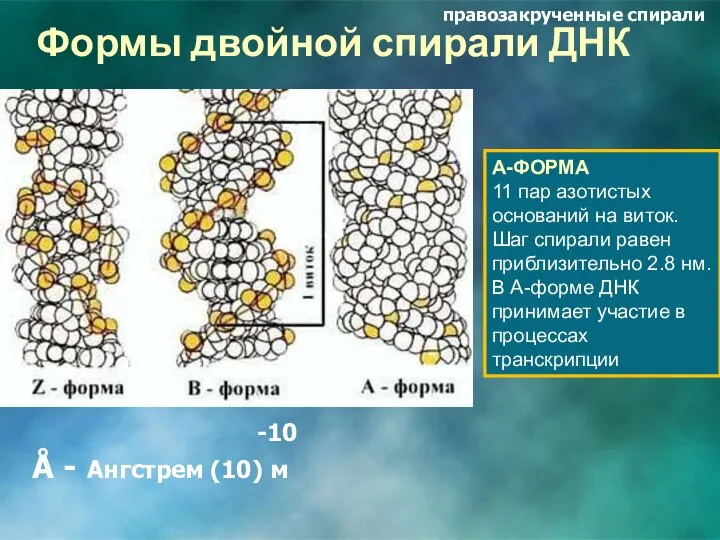
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
А-ФОРМА
11 пар
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
А-ФОРМА
11 пар
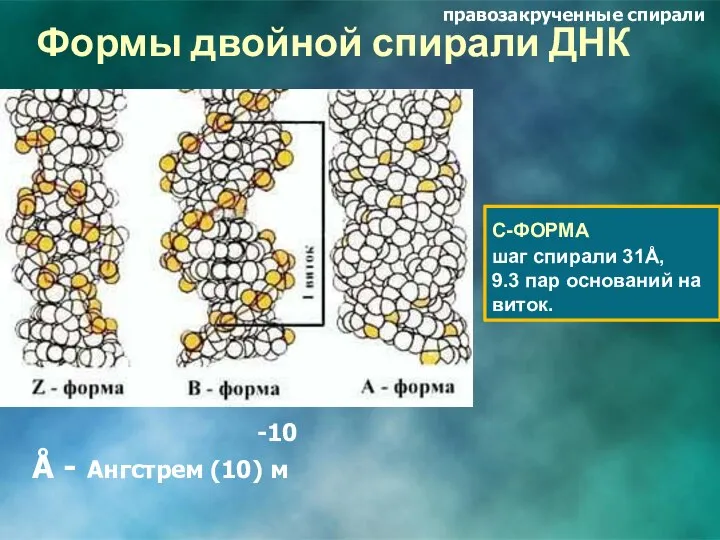
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
С-ФОРМА
шаг спирали
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
С-ФОРМА
шаг спирали
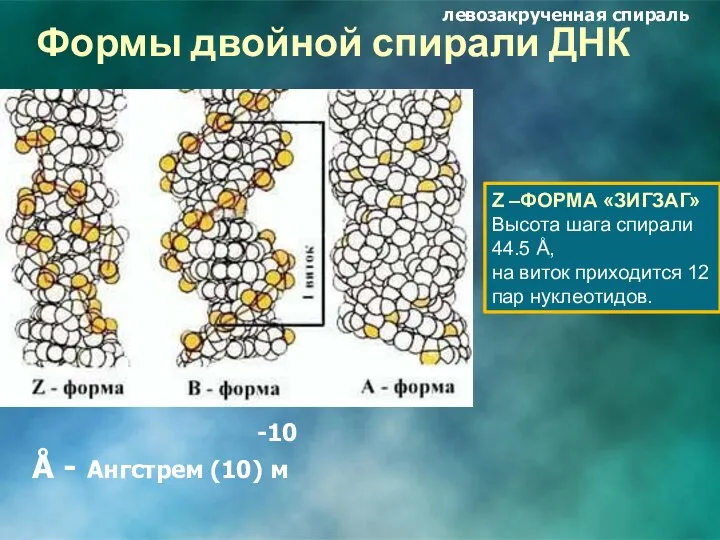
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
Z –ФОРМА «ЗИГЗАГ»
Формы двойной спирали ДНК
-10
Å - Ангстрем (10) м
Z –ФОРМА «ЗИГЗАГ»

Форма SBS (side by side)
лишена взаимозакрученности в двойную спираль.
Форма SBS (side by side)
лишена взаимозакрученности в двойную спираль.

МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Молекулярная организация ядерного хроматина
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. НУКЛЕИНОВЫЕ КИСЛОТЫ.
Молекулярная организация ядерного хроматина
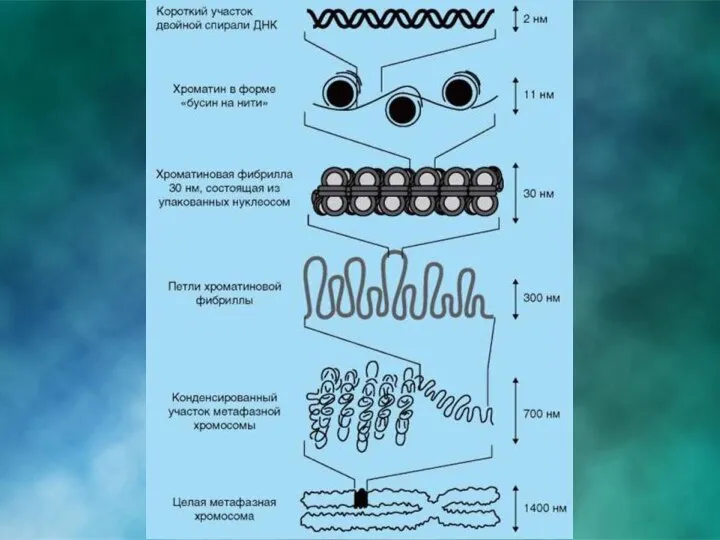

Третичная структура. Уровни компактизации ДНК. НУКЛЕОСОМА
– это повторяющийся структурный элемент
Третичная структура. Уровни компактизации ДНК. НУКЛЕОСОМА
– это повторяющийся структурный элемент

Гистоны – основные белки
Выделяют 5 фракций гистонов
Образуется около 60 млн. молекул
Гистоны – основные белки
Выделяют 5 фракций гистонов
Образуется около 60 млн. молекул

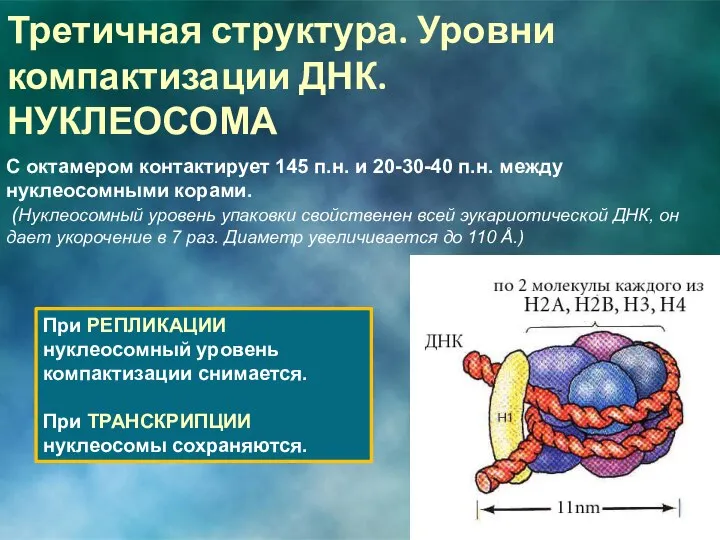
С октамером контактирует 145 п.н. и 20-30-40 п.н. между нуклеосомными корами.
С октамером контактирует 145 п.н. и 20-30-40 п.н. между нуклеосомными корами.
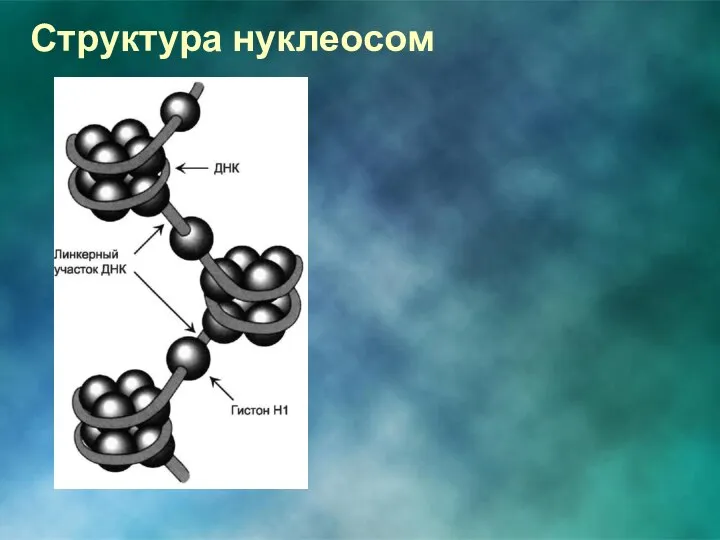
Структура нуклеосом
Структура нуклеосом
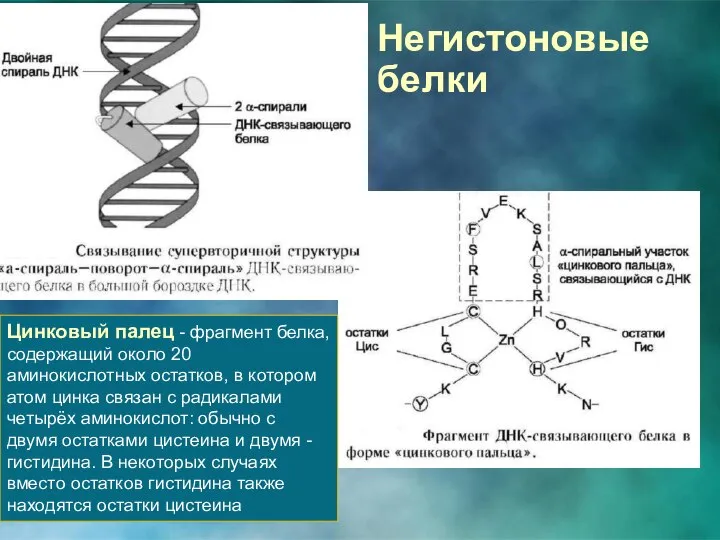
Негистоновые белки
Цинковый палец - фрагмент белка, содержащий около 20 аминокислотных остатков,
Негистоновые белки
Цинковый палец - фрагмент белка, содержащий около 20 аминокислотных остатков,

Н1 взаимодействует с ОКТАМЕРАМИ, сближает их, на него наматывается ДНК. Образуется
Н1 взаимодействует с ОКТАМЕРАМИ, сближает их, на него наматывается ДНК. Образуется

Негистоновые белки узнают определенные последовательности ДНК и связываются с ними и
Негистоновые белки узнают определенные последовательности ДНК и связываются с ними и

Третичная структура. Уровни компактизации ДНК. МЕТАФАЗНАЯ ХРОМОСОМА
Третичная структура. Уровни компактизации ДНК. МЕТАФАЗНАЯ ХРОМОСОМА
 Максимизация прибыли в краткосрочном периоде Выполнила Редок Полина, студентка ЭФ группы Э122Б
Максимизация прибыли в краткосрочном периоде Выполнила Редок Полина, студентка ЭФ группы Э122Б Образование Белгородской области Презентация для модуля краеведения на уроках природоведения 5 класса Выполнил: учитель природ
Образование Белгородской области Презентация для модуля краеведения на уроках природоведения 5 класса Выполнил: учитель природ Перспектива группы геометрических тел. Принципы формообразования и этапы выполнения рисунка. (Лекция 4)
Перспектива группы геометрических тел. Принципы формообразования и этапы выполнения рисунка. (Лекция 4) Corporate culture in Sweden
Corporate culture in Sweden Психология восприятия 1
Психология восприятия 1 Два пути
Два пути Этика и этикет
Этика и этикет SEO Коучинг. Семантическое ядро. часть 2
SEO Коучинг. Семантическое ядро. часть 2 Microsoft official course. Creating methods, handling exceptions, and monitoring applications. (Module 2)
Microsoft official course. Creating methods, handling exceptions, and monitoring applications. (Module 2) Военная биография моего прадеда Нестерова Михаила Нестеровича
Военная биография моего прадеда Нестерова Михаила Нестеровича Подумаем!
Подумаем! Витоки української культури
Витоки української культури Культура Китая. Изобразительное искусство. Традиции и новации
Культура Китая. Изобразительное искусство. Традиции и новации Фундаменты. Сборные фундаменты
Фундаменты. Сборные фундаменты Мы, выпускники 2000 года, благодарим наших любимых учителей за то, что они ввели нас в большой и интересный мир знаний! Мы, выпускники
Мы, выпускники 2000 года, благодарим наших любимых учителей за то, что они ввели нас в большой и интересный мир знаний! Мы, выпускники  Язык C++
Язык C++ Вне конкуренции. Вне границ
Вне конкуренции. Вне границ 3GPP Long Term Evolution (LTE). Обзор физического уровня
3GPP Long Term Evolution (LTE). Обзор физического уровня Монтаж оптических кабелей
Монтаж оптических кабелей hessen
hessen Валютный рынок и конвертируемость валют Экономика, профильный уровень, 11 класс, учитель Г.И. Непершина, МОУ СОШ № 15 г. Балашова Сар
Валютный рынок и конвертируемость валют Экономика, профильный уровень, 11 класс, учитель Г.И. Непершина, МОУ СОШ № 15 г. Балашова Сар Люминесцентный сенсор кислородный на основе иридиевых комплексов
Люминесцентный сенсор кислородный на основе иридиевых комплексов Работа, энергия. Применение законов сохранения
Работа, энергия. Применение законов сохранения  Банковская система
Банковская система  Волновая оптика. Электромагнитные волны. Уравнения Максвелла и волновое уравнение.
Волновая оптика. Электромагнитные волны. Уравнения Максвелла и волновое уравнение. Презентация на тему "Формирование возрастных особенностей и половых различий подростков" - скачать презентации по Педагогик
Презентация на тему "Формирование возрастных особенностей и половых различий подростков" - скачать презентации по Педагогик Презентация на тему "Психолого-педагогическое сопровождение первоклассников в адаптационный период" - скачать презентации п
Презентация на тему "Психолого-педагогическое сопровождение первоклассников в адаптационный период" - скачать презентации п Валютное право: понятие, предмет и метод Валютное право Лекция 3
Валютное право: понятие, предмет и метод Валютное право Лекция 3