- Вирусы

Содержание
- 2. Классификация вирусов. Классификация включает 4 иерархических уровня: порядок, семейство (иногда и подсемейство), род и вид. На
- 3. Морфология вирусов Морфологию вирусов изучают с помощью электронной микроскопии, так как их размеры малы (18-400 нм)
- 4. СТРОЕНИЕ ВИРУСОВ Простые вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом (от лат. capsa
- 5. СТРОЕНИЕ ВИРУСОВ Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип
- 6. Просто устроенные вирусы (без оболочки) Пример просто устроенных вирусов - вирус гепатита А и паповавирус с
- 7. Сложно устроенные вирусы (с оболочкой ) У сложно устроенных вирусов (например, у вирусов герпеса, гриппа, флавивирусов)
- 8. Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус РНК)
- 9. Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК)
- 10. Репродукция вирусов Различают три типа взаимодействия вируса с клеткой: 1. продуктивный тип, при котором образуются новые
- 11. Репродукция вирусов Продуктивный тип взаимодействия вируса с клеткой - репродукция вируса проходит несколько стадий: 1) адсорбция
- 12. Репродукция вирусов Механизм репродукции отличается у вирусов, имеющих: 1) двунитевую ДНК; 2) однонитевую ДНК; 3) плюс
- 13. Репродукция вирусов Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы, тогавирусы), у которых
- 14. Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы Поглощенный вирус поставляет геном в ядро клетки. Парвовирусы
- 15. Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы, тогавирусы), у которых геномная плюс-нить
- 16. Минус однонитевые РНК–вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Схема репродукции парамиксовирусов Вирус
- 17. Ретровирусы (плюс-нитевые диплоидные РНК-вирусы). Схема репродукции вируса иммунодефицита человека (ВИЧ) ВИЧ связывается гликопротеином gp120 (1) с
- 18. Культивирование и индикация вирусов Вирусы культивируют в организме лабораторных животных, в развивающихся куриных эмбрионах и культурах
- 19. Основоположник вирусологии профессор Д.И. Ивановский (1864 - 1920)
- 22. Куриные эмбрионы. В середине 30-х годов австралийский вирусолог Ф. Вернет «открыл» новое для вирусологии экспериментальное животное
- 23. Академик АМН СССР А.А. Смородинцев (1901-1986)
- 24. Академик АМН СССР М.П. Чумаков (1909-1993)
- 25. Не инфицированы ЛИНИЯ КЛЕТОК ПОЧКИ СИРИЙСКОГО ХОМЯЧКА – BHK-21
- 26. Разведение 106 Typical CPE
- 27. Разведение 105
- 28. 4 Разведение 104 In concentration higher than 1E4, all cells were detached from the plate
- 29. ПЕРЕВИВАЕМАЯ ЛИНИЯ КЛЕТОК ПОЧКИ ЭМБРИОНА ТЕЛЯТ
- 30. MDBK клеточные культуры на 7-й день после заражения
- 31. ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в
- 32. Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом
- 33. Бляшки, или “негативные” колонии — ограниченные участки разрушенных вирусами клеток, культивируемых на питательной среде под агаровым
- 34. РГА( реакция гемагглютинации) основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов за счет вирусных гликопротеиновых
- 36. Скачать презентацию

Классификация вирусов.
Классификация включает 4 иерархических уровня: порядок, семейство (иногда и подсемейство),
Классификация вирусов.
Классификация включает 4 иерархических уровня: порядок, семейство (иногда и подсемейство),

Морфология вирусов
Морфологию вирусов изучают с помощью электронной микроскопии, так как их
Морфология вирусов
Морфологию вирусов изучают с помощью электронной микроскопии, так как их

СТРОЕНИЕ ВИРУСОВ
Простые вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой
СТРОЕНИЕ ВИРУСОВ
Простые вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой

СТРОЕНИЕ ВИРУСОВ
Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный
СТРОЕНИЕ ВИРУСОВ
Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный
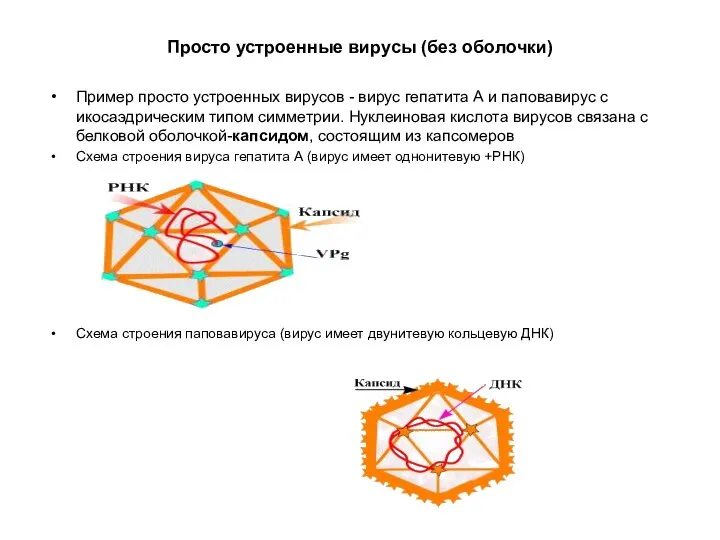
Просто устроенные вирусы (без оболочки)
Пример просто устроенных вирусов - вирус гепатита
Просто устроенные вирусы (без оболочки)
Пример просто устроенных вирусов - вирус гепатита

Сложно устроенные вирусы (с оболочкой )
У сложно устроенных вирусов (например, у
Сложно устроенные вирусы (с оболочкой )
У сложно устроенных вирусов (например, у
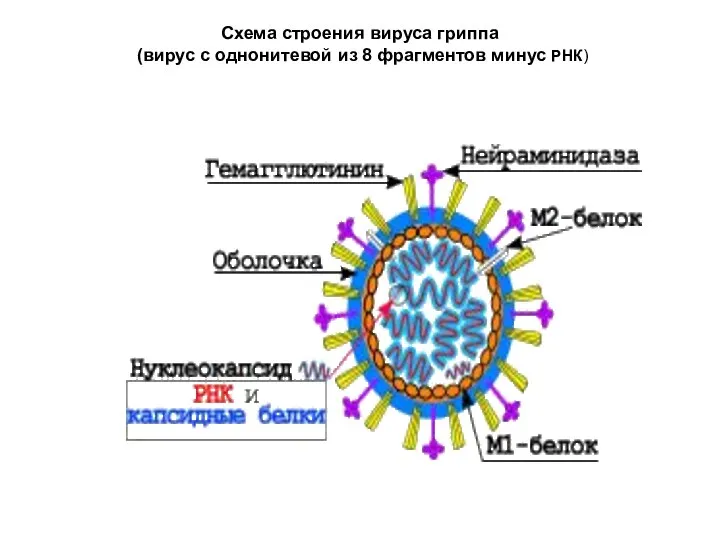
Схема строения вируса гриппа
(вирус с однонитевой из 8 фрагментов минус
Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус
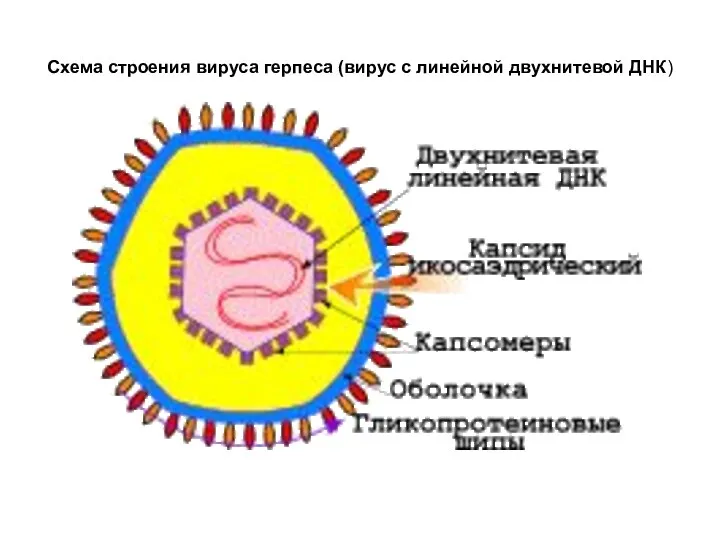
Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК)
Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК)

Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный
Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный

Репродукция вирусов
Продуктивный тип взаимодействия вируса с клеткой - репродукция вируса проходит
Репродукция вирусов
Продуктивный тип взаимодействия вируса с клеткой - репродукция вируса проходит

Репродукция вирусов
Механизм репродукции отличается у вирусов, имеющих: 1) двунитевую ДНК; 2)
Репродукция вирусов
Механизм репродукции отличается у вирусов, имеющих: 1) двунитевую ДНК; 2)

Репродукция вирусов
Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы,
Репродукция вирусов
Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы,
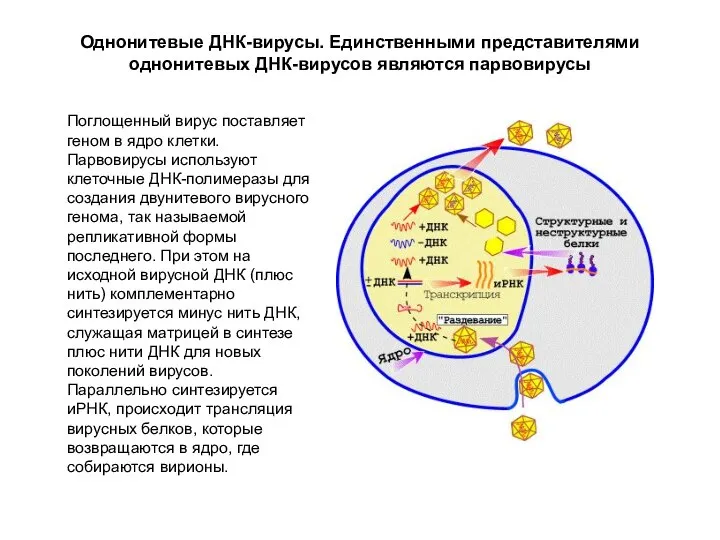
Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы
Поглощенный вирус поставляет геном
Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы
Поглощенный вирус поставляет геном
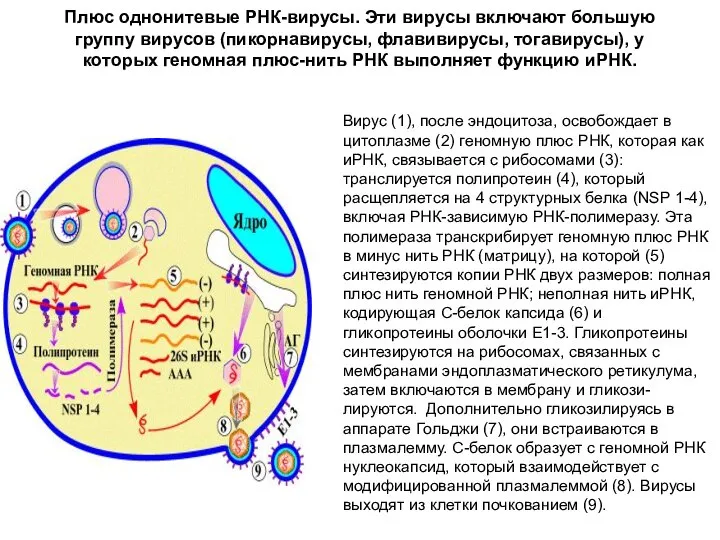
Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы,
Плюс однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы,
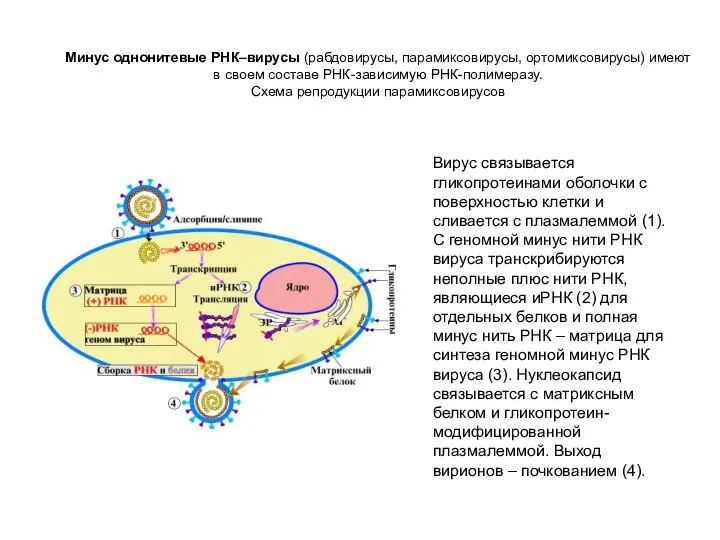
Минус однонитевые РНК–вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую
Минус однонитевые РНК–вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую
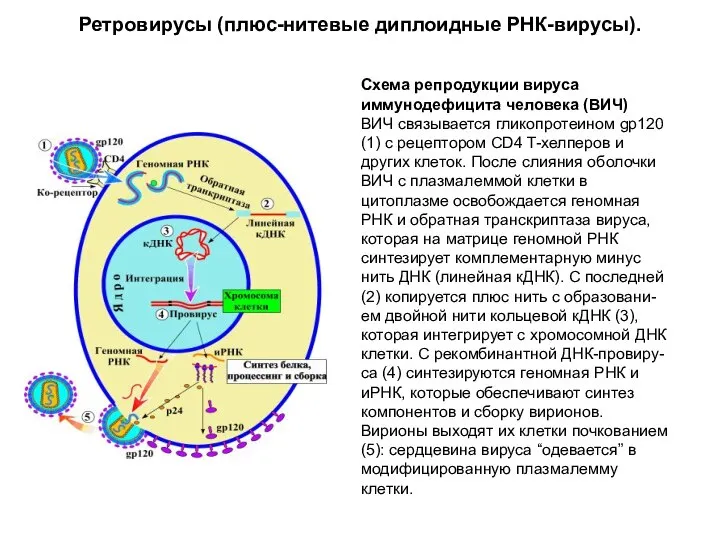
Ретровирусы (плюс-нитевые диплоидные РНК-вирусы).
Схема репродукции вируса иммунодефицита человека (ВИЧ)
ВИЧ связывается гликопротеином
Ретровирусы (плюс-нитевые диплоидные РНК-вирусы).
Схема репродукции вируса иммунодефицита человека (ВИЧ)
ВИЧ связывается гликопротеином

Культивирование и индикация вирусов
Вирусы культивируют в организме лабораторных животных, в
Культивирование и индикация вирусов
Вирусы культивируют в организме лабораторных животных, в

Основоположник вирусологии
профессор
Д.И. Ивановский
(1864 - 1920)
Основоположник вирусологии
профессор
Д.И. Ивановский
(1864 - 1920)



Куриные эмбрионы.
В середине 30-х годов австралийский вирусолог Ф. Вернет «открыл»
Куриные эмбрионы.
В середине 30-х годов австралийский вирусолог Ф. Вернет «открыл»

Академик АМН СССР
А.А. Смородинцев
(1901-1986)
Академик АМН СССР
А.А. Смородинцев
(1901-1986)

Академик АМН СССР
М.П. Чумаков
(1909-1993)
Академик АМН СССР
М.П. Чумаков
(1909-1993)

Не инфицированы
ЛИНИЯ КЛЕТОК ПОЧКИ СИРИЙСКОГО ХОМЯЧКА – BHK-21
Не инфицированы
ЛИНИЯ КЛЕТОК ПОЧКИ СИРИЙСКОГО ХОМЯЧКА – BHK-21

Разведение 106
Typical CPE
Разведение 106
Typical CPE
Разведение 105
Разведение 105

4
Разведение 104
In concentration higher than 1E4, all cells were detached from
4
Разведение 104
In concentration higher than 1E4, all cells were detached from

ПЕРЕВИВАЕМАЯ ЛИНИЯ КЛЕТОК ПОЧКИ ЭМБРИОНА ТЕЛЯТ
ПЕРЕВИВАЕМАЯ ЛИНИЯ КЛЕТОК ПОЧКИ ЭМБРИОНА ТЕЛЯТ

MDBK клеточные культуры на 7-й день после заражения
MDBK клеточные культуры на 7-й день после заражения
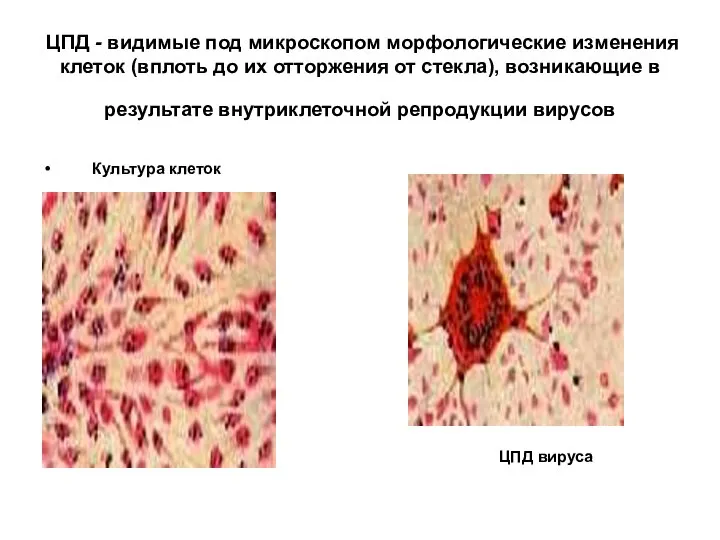
ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их
ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их
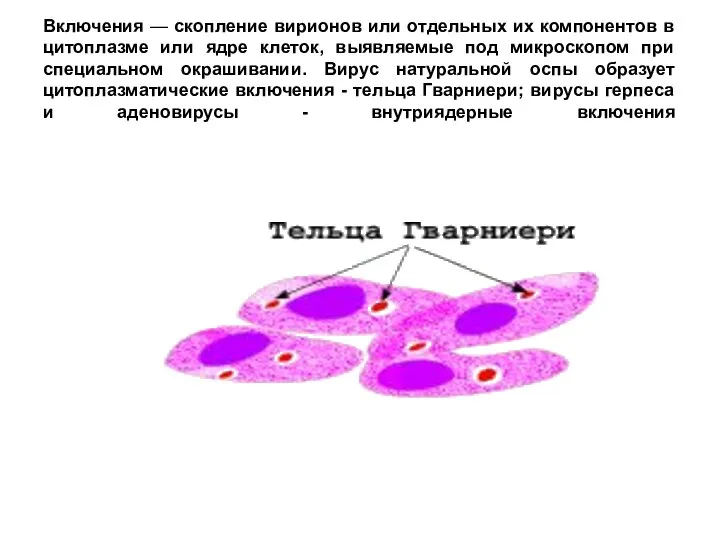
Включения — скопление вирионов или отдельных их компонентов в цитоплазме или
Включения — скопление вирионов или отдельных их компонентов в цитоплазме или
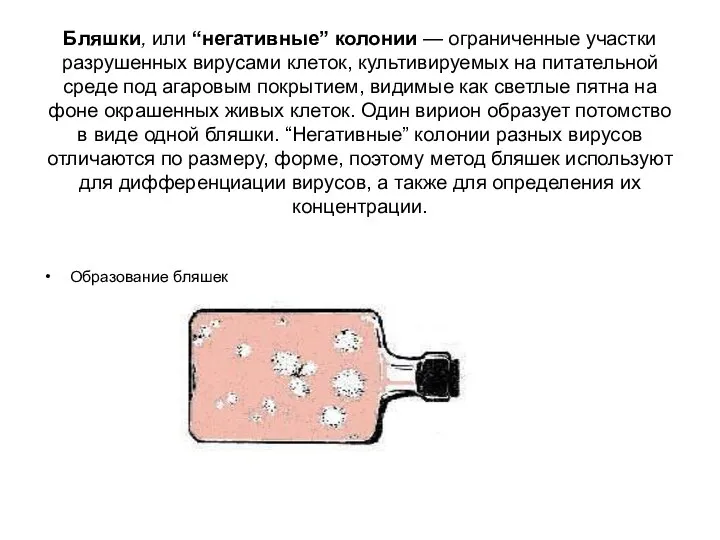
Бляшки, или “негативные” колонии — ограниченные участки разрушенных вирусами клеток, культивируемых
Бляшки, или “негативные” колонии — ограниченные участки разрушенных вирусами клеток, культивируемых
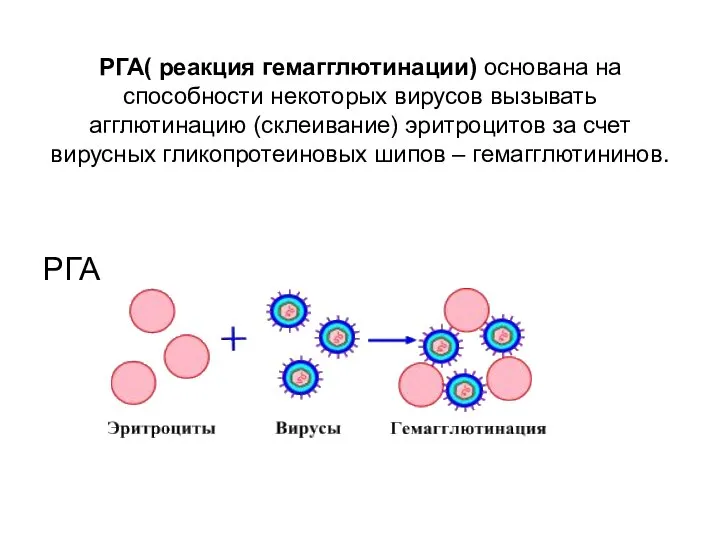
РГА( реакция гемагглютинации) основана на способности некоторых вирусов вызывать агглютинацию (склеивание)
РГА( реакция гемагглютинации) основана на способности некоторых вирусов вызывать агглютинацию (склеивание)
 Неделя семьи. Построение брака и семьи
Неделя семьи. Построение брака и семьи Законы диалектики
Законы диалектики Требования к технологической документации. Справочная, исходная и сопроводительная документация
Требования к технологической документации. Справочная, исходная и сопроводительная документация Решение задач с использованием условного оператора
Решение задач с использованием условного оператора Эрнест Резерфорд
Эрнест Резерфорд Организация технического обслуживания и ремонта автомобиля КамАЗ 6460
Организация технического обслуживания и ремонта автомобиля КамАЗ 6460 Современные подходы к анализу урока русского языка Громова В.И., к.ф.н., доцент кафедры филологического образования СарИПКиПРО
Современные подходы к анализу урока русского языка Громова В.И., к.ф.н., доцент кафедры филологического образования СарИПКиПРО Как празднуют Новый год в разных странах мира
Как празднуют Новый год в разных странах мира Программирование на языке MATLAB. Подпрограммы и функции
Программирование на языке MATLAB. Подпрограммы и функции Православие. Библия
Православие. Библия Тема: « Предмет и задачи этнопедагогики»
Тема: « Предмет и задачи этнопедагогики» Учимся записывать орф задачи
Учимся записывать орф задачи Логические схемы полусумматора и триггера
Логические схемы полусумматора и триггера  Семь чудес света
Семь чудес света  Священный месяц Рамадан
Священный месяц Рамадан Блиц опрос (ДКБ)
Блиц опрос (ДКБ) Информационный сервис “Древо познания” в форме веб-сайта
Информационный сервис “Древо познания” в форме веб-сайта Презентация "Социальный проект" - скачать презентации по Экономике
Презентация "Социальный проект" - скачать презентации по Экономике Презентация Понятия и виды субъектов трудового права
Презентация Понятия и виды субъектов трудового права  Пять самых известных храмов Японии
Пять самых известных храмов Японии Московская Русь
Московская Русь День Конституции России
День Конституции России JavaScript. Основы программирования
JavaScript. Основы программирования Достопримечальности Киева
Достопримечальности Киева Назначение и общий принцип функционирования коммутатора-усилителя. Усилитель мощности: назначение и технические данные. Устройст
Назначение и общий принцип функционирования коммутатора-усилителя. Усилитель мощности: назначение и технические данные. Устройст Operators 413/415. Electromechanical operators for swing gates
Operators 413/415. Electromechanical operators for swing gates Технология Wi-Fi
Технология Wi-Fi Economy of America
Economy of America