- Биоэлектрические явления в возбудимых тканях

Содержание
- 2. Биоэлектрические явления в возбудимых тканях Возбуждение характеризуется совокупностью электрических, химических, функциях и структурных изменений живой клетки.
- 3. Биоэлектрические явления в возбудимых тканях 2 - перебрасывая нервы на обнаженные мышцы голени - они сокращались.
- 4. Электрические явления в живых тканях Электрические явления в живых тканях были разработаны в 40-50 г. ХIХ
- 5. Биологические мембраны Биологические мембраны, строение и функции. Состоит из билипидного слоя и фосфалипидов, гидрофильные, гидрофобными концами
- 6. Электрические ответы возбудимых тканей, клеток 1. Локальный ответ; 2. Распространяющийся ПД; 3. Следовые потенциалы; 4. Возбуждающиеся
- 7. ПОТЕНЦИАЛ ПОКОЯ (ПП) Измеряет микроэлектродом. В состоянии покоя между наружной и внутренний поверхностью существует разность потенциалов
- 8. Происхождение потенциала покоя. Мембранно-ионная теория Бернштейна (1902) модифицированы Ходжкиным,Хаксли (1952)- биоэлектрические потенциалы обусловлены неодинаковой концентрацией ионов
- 9. Потенциал покоя 1. Потенциал покоя определяется разностью концентрации; 2. Соотношением проницаемости для этих ионов. Роль обмена
- 10. Изменение возбудимости при возбуждении: . 1. локальный ответ местное повышение возбудимости; 2. Абсолютная рефрактерность; 3. Относительная
- 11. Изменение возбудимости при возбуждении При возбуждении обмен веществ усиливается в цитоплазме мембраны. Усиливается распад АТР, КФ,
- 12. Изменение возбудимости при возбуждении Инверсия – пик или заряд Реполяризация – восстановлении исходного ПД 1>K+ -----
- 14. Скачать презентацию

Биоэлектрические явления в возбудимых тканях
Возбуждение характеризуется совокупностью электрических, химических, функциях и
Биоэлектрические явления в возбудимых тканях
Возбуждение характеризуется совокупностью электрических, химических, функциях и

Биоэлектрические явления в возбудимых тканях
2 - перебрасывая нервы на обнаженные мышцы
Биоэлектрические явления в возбудимых тканях
2 - перебрасывая нервы на обнаженные мышцы

Электрические явления в живых тканях
Электрические явления в живых тканях были разработаны
Электрические явления в живых тканях
Электрические явления в живых тканях были разработаны

Биологические мембраны
Биологические мембраны, строение и функции. Состоит из билипидного слоя и
Биологические мембраны
Биологические мембраны, строение и функции. Состоит из билипидного слоя и

Электрические ответы возбудимых тканей, клеток
1. Локальный ответ;
2. Распространяющийся ПД;
Электрические ответы возбудимых тканей, клеток
1. Локальный ответ;
2. Распространяющийся ПД;

ПОТЕНЦИАЛ ПОКОЯ (ПП)
Измеряет микроэлектродом. В состоянии покоя между наружной и внутренний
ПОТЕНЦИАЛ ПОКОЯ (ПП)
Измеряет микроэлектродом. В состоянии покоя между наружной и внутренний

Происхождение потенциала покоя.
Мембранно-ионная теория Бернштейна (1902) модифицированы Ходжкиным,Хаксли (1952)- биоэлектрические потенциалы
Происхождение потенциала покоя.
Мембранно-ионная теория Бернштейна (1902) модифицированы Ходжкиным,Хаксли (1952)- биоэлектрические потенциалы

Потенциал покоя
1. Потенциал покоя определяется разностью концентрации;
2. Соотношением проницаемости для
Потенциал покоя
1. Потенциал покоя определяется разностью концентрации;
2. Соотношением проницаемости для

Изменение возбудимости при возбуждении:
. 1. локальный ответ местное повышение возбудимости;
2.
Изменение возбудимости при возбуждении:
. 1. локальный ответ местное повышение возбудимости;
2.

Изменение возбудимости при возбуждении
При возбуждении обмен веществ усиливается в цитоплазме мембраны.
Изменение возбудимости при возбуждении
При возбуждении обмен веществ усиливается в цитоплазме мембраны.

Изменение возбудимости при возбуждении
Инверсия – пик или заряд
Реполяризация – восстановлении исходного
Изменение возбудимости при возбуждении
Инверсия – пик или заряд
Реполяризация – восстановлении исходного
 Презентация Полевые сорняки
Презентация Полевые сорняки 13 пар черепных нервов
13 пар черепных нервов Скелет человека
Скелет человека Каракал. Степной рысь
Каракал. Степной рысь Д.З. п.16
Д.З. п.16  Матричные биосинтезы
Матричные биосинтезы Значение воды в жизни человека
Значение воды в жизни человека Кормление косули в естественных условиях и в условиях зоопарка
Кормление косули в естественных условиях и в условиях зоопарка Признаки живых организмов. Урок биологии - 5 класс
Признаки живых организмов. Урок биологии - 5 класс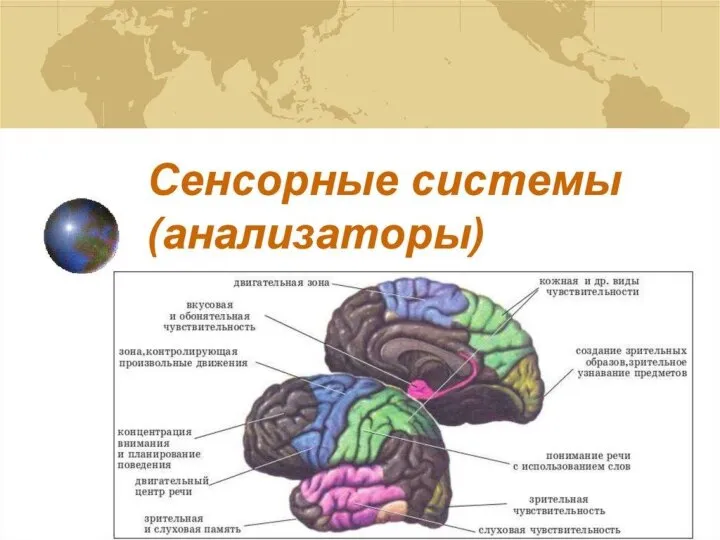 Анализаторы
Анализаторы Возбудимость мышечных тканей
Возбудимость мышечных тканей Тип Моллюски. Класс Головоногие моллюски
Тип Моллюски. Класс Головоногие моллюски Тигр - самая большая кошка на Земле
Тигр - самая большая кошка на Земле Положение человека в системе органического мира
Положение человека в системе органического мира Самые распространенные породы домашних свиней В настоящее время существует множество пород свиней. Некоторые из них распрост
Самые распространенные породы домашних свиней В настоящее время существует множество пород свиней. Некоторые из них распрост Гормоны. Регуляция обмена веществ. Биохимия гормонов. Выполнили: Темирова Н. и Тлебалдиева Ф. Проверил: Шарипов К.О.
Гормоны. Регуляция обмена веществ. Биохимия гормонов. Выполнили: Темирова Н. и Тлебалдиева Ф. Проверил: Шарипов К.О.  Генетика - наука о законах и механизмах наследственности и изменчивости организмов.
Генетика - наука о законах и механизмах наследственности и изменчивости организмов.  Размножение. Строение половых клеток. Гаметогенез. Мейоз. Оплодотворение
Размножение. Строение половых клеток. Гаметогенез. Мейоз. Оплодотворение «Коровьи» растения
«Коровьи» растения Мелкие млекопитающие- постоянные обитатели живого уголка
Мелкие млекопитающие- постоянные обитатели живого уголка Основы учения об эволюции. Эволюционная теория Ч. Дарвина
Основы учения об эволюции. Эволюционная теория Ч. Дарвина Адаптации организмов к факторам среды
Адаптации организмов к факторам среды ВВЕДЕНИЕ. Что изучает наука БИОЛОГИЯ
ВВЕДЕНИЕ. Что изучает наука БИОЛОГИЯ Урок биологии 11 класс
Урок биологии 11 класс Презентация на тему "Черепахи" - скачать презентации по Биологии
Презентация на тему "Черепахи" - скачать презентации по Биологии Фауна Африки
Фауна Африки Анатомия и физиология нервной системі
Анатомия и физиология нервной системі Деревья и кустарники
Деревья и кустарники