- Биосинтез белков. Фактор терминации трансляции тРНК. (Лекция 14)

Содержание
- 2. Этап 1 (связывание аминоацил-тРНК) катализируется фактором элонгации EF1 (эукариоты) или EF-Tu (прокариоты), а этап 3 (транс-локация)
- 3. Факторы элонгации продвигают трансляцию и повышают ее точность. Два фактора элонгации входят в рибосому и покидают
- 4. Эпицикл трансляции (задает фазу по триплетам) Стадии инициации и терминации – это модификации стадии элонгации. В
- 5. Инициация трансляции – это серия молекулярных событий, происходящих с рибосомой, которая приводит к взаимодействию рибосомы с
- 6. Основные события при инициации трансляции Инициация начинается с диссоциации рибосомы. Малая субчастица связывает специальные белки –
- 7. Участки связывания рибосом (RBS) в мРНК Наличие RBS в мРНК обеспечивают возможность малой субчастице отличить стартовый
- 8. Инициаторы трансляции у про- и эукариот Специальная инициаторная метионил-тРНК (Met-тРНКiMet ) с тем же антикодоном CAU,
- 9. Неканоническое спаривание третьего (редко - первого) нуклеотида антикодона CAU тРНКFMet прокариот Малая субчастица обеспечивает взаимодействие инициаторного
- 10. Способы распознавания инициаторного кодона мРНК у эукариот и прокариот Эукариоты (моноцистронная мРНК): терминальная инициация – трансляция
- 11. Внутренняя инициация на полицистронной мРНК прокариот Рибосома прокариот узнает в мРНК участки связывания рибосомы (RBS) -
- 12. Узнавание инициирующего кодона на мРНК прокариот Инициирующий триплет AUG (GUG, UUG и др.) расположен через 3-10
- 13. «Терминальная инициация по сканирующему механизму» происходит с первого от 5’-конца триплета AUG, который находится в оптимальном
- 14. Фактор инициации IF3 (eIF3) -это фактор диссоциации субчастиц рибосомы IF2:ГТФ – аналог фактора элонгации EF1:ГТФ, связывается
- 15. Инициация трансляции (2): ассоциация субчастиц и функции большой субчастицы IF2:ГТФ Большая субчастица «наводит» ГТРазную активность на
- 16. Инициация синтеза белка у эукариот Для эффективного запуска трансляции необходимо, чтобы поли-А хвост мРНК был связан
- 17. Элонгация пептида на рибосоме Меt-tRNA Val-tRNA а - инициаторная метионил-тРНК в Р-участке; первая элонгаторная валил-тРНК, комплементарная
- 18. Сопряженная транскрипция-трансляция у прокариот Сопряжение осуществляется в пространстве (комплекс ДНК : РНК-полимераза : мРНК) и во
- 19. Эпицикл трансляции ИНИЦИАЦИЯ – ЭЛОНГАЦИЯ – ТЕРМИНАЦИЯ мРНК: нет специального триплеты терминаторный инициаторного триплета аминокислот триплет
- 20. Терминация трансляции Стадия терминации трансляции может рассматриваться как модифицированный элонгационный цикл, где пептидил-тРНК в качестве донорного
- 21. «Универсальный» кодовый словарь и факторы терминации прокариот RF1 RF2
- 22. Терминация трансляции а - после добавления к растущему полипептиду С-концевого аминокислотного остатка в А-участке устанавливается кодон
- 23. Молекулярная мимикрия: структура фактора терминации еRF1человека и его сходство с молекулой тРНК
- 24. «Портрет» большой рибосомной субчастицы бактерий белки рРНК Со стороны малой субчастицы С другой строны Слабое вращение
- 25. Комплекс терминации/реинициации трансляции у эукариот: петля 5‘кэп - 3‘поли-A-хвост мРНК и участие поли(А)-связывающего белка (PABP) eRF1–eRF3
- 26. Синтез большинства белковых молекул занимает от 20 секунд до нескольких минут. В течение этого промежутка времени
- 27. Стадии инициации и терминации – это модифицированный элонгационный цикл При инициации Met-tRNAfMet:IF2:GTP аналогична Aa-tRNA:EF1:GTP при элонгации.
- 28. Транспортно-матричная РНК (тмРНК) Транспортно-матричная РНК (тмРНК) содержит элементы как мРНК, так и тРНК. Матричная часть тмРНК
- 29. Роль транспортно-матричной РНК (тмРНК) Деградация КОНТРОЛЬ КАЧЕСТВА мРНК Деградация КОНТРОЛЬ КАЧЕСТВА БЕЛКА Освобождение мРНК Маркировка (tagging)
- 30. Регуляция трансляции Регуляция осуществляется путем изменения интенсивности трансляции на данной мРНК (в диапозоне от индукции трансляции
- 31. Позитивная регуляция: дискриминация мРНК Различные мРНК обладают разным сродством к инициирующим рибосомным частицам У прокариотических организмов
- 32. «Сила» мРНК в значительной мере определяет соотношение продукции различных белков в клетке Структурные белки мембран, рибосомные
- 33. Трансляционное сопряжение у прокариот В полицистронных мРНК работает механизм внутренней инициации, и рибосомы во многих случаях
- 34. Независимая и сопряженная инициация трансляции последовательных цистронов прокариотических мРНК а - независимая инициация трансляции б -
- 35. Негативная регуляция: трансляционная репрессия а) Сайт-специфические РНК-связывающие белки подавляют трансляцию специфических мРНК, блокируя доступ рибосомы к
- 36. Свойственно только эукариотам. Связывание маскирующего белка с сегментом маскирования в З'-нетранслируемой области (З'-UTR) мРНК приводит к
- 37. Функионирование RISC c miRNA и siRNA Мисмэтчи miRNA с мРНК не расщепляются и накапливаются в Р-тельцах
- 38. Маскирование мРНК у эукариот Маскирование и демаскирование мРНК являются особенно характерными для процессов гаметогенеза (оогенеза и
- 40. Скачать презентацию
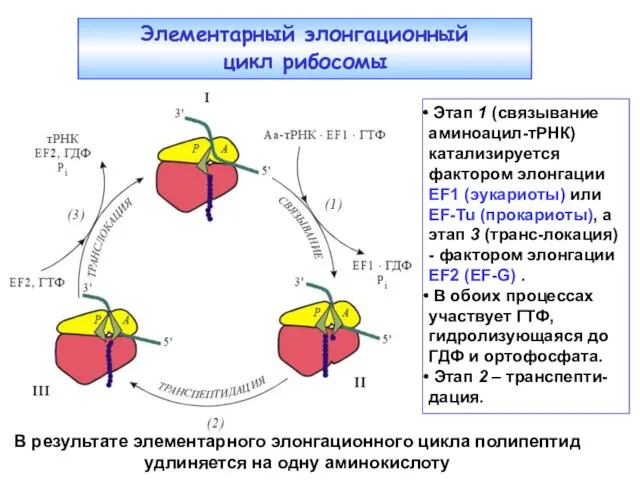
Этап 1 (связывание аминоацил-тРНК) катализируется фактором элонгации EF1 (эукариоты) или
Этап 1 (связывание аминоацил-тРНК) катализируется фактором элонгации EF1 (эукариоты) или
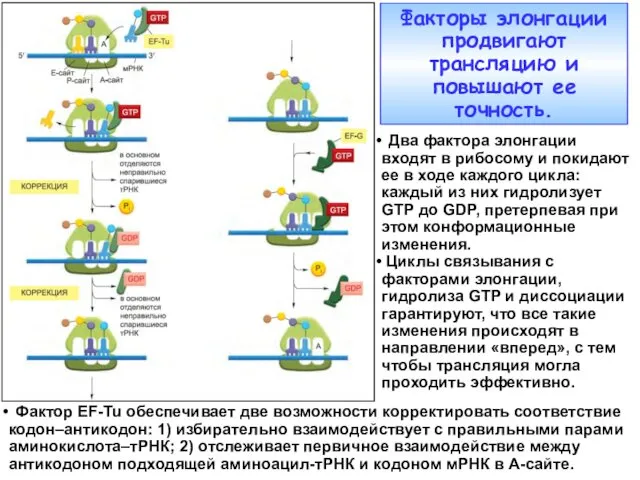
Факторы элонгации продвигают трансляцию и повышают ее точность.
Два фактора элонгации
Факторы элонгации продвигают трансляцию и повышают ее точность.
Два фактора элонгации
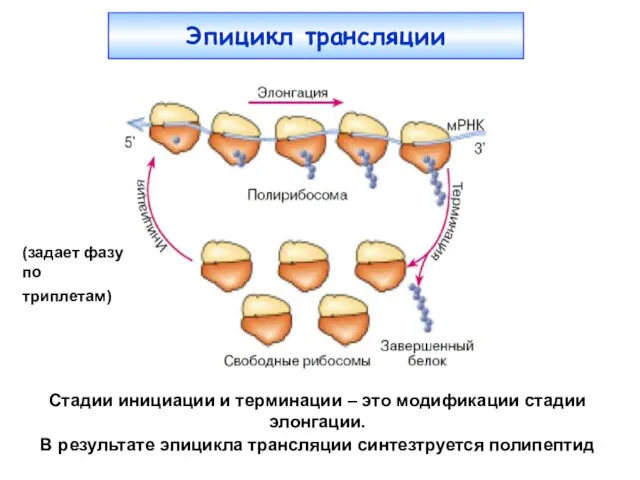
Эпицикл трансляции
(задает фазу
по триплетам)
Стадии инициации и терминации – это
Эпицикл трансляции
(задает фазу
по триплетам)
Стадии инициации и терминации – это

Инициация трансляции – это серия молекулярных событий, происходящих с рибосомой, которая
Инициация трансляции – это серия молекулярных событий, происходящих с рибосомой, которая

Основные события при инициации трансляции
Инициация начинается с диссоциации рибосомы.
Малая субчастица
Основные события при инициации трансляции
Инициация начинается с диссоциации рибосомы.
Малая субчастица
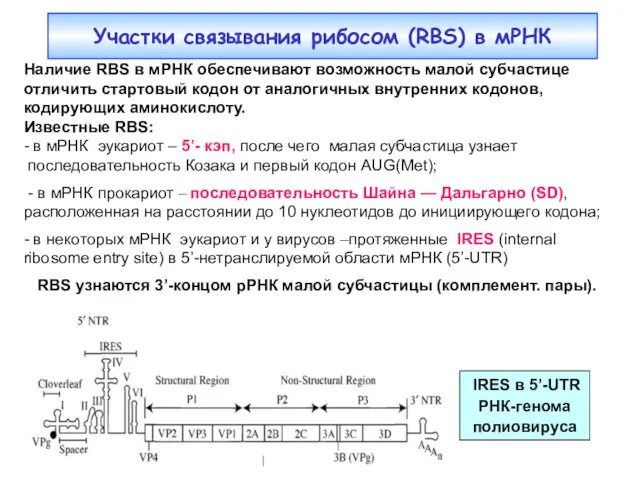
Участки связывания рибосом (RBS) в мРНК
Наличие RBS в мРНК обеспечивают возможность
Участки связывания рибосом (RBS) в мРНК
Наличие RBS в мРНК обеспечивают возможность

Инициаторы трансляции у про- и эукариот
Специальная инициаторная метионил-тРНК (Met-тРНКiMet )
Инициаторы трансляции у про- и эукариот
Специальная инициаторная метионил-тРНК (Met-тРНКiMet )

Неканоническое спаривание третьего (редко - первого) нуклеотида антикодона CAU тРНКFMet прокариот
Неканоническое спаривание третьего (редко - первого) нуклеотида антикодона CAU тРНКFMet прокариот
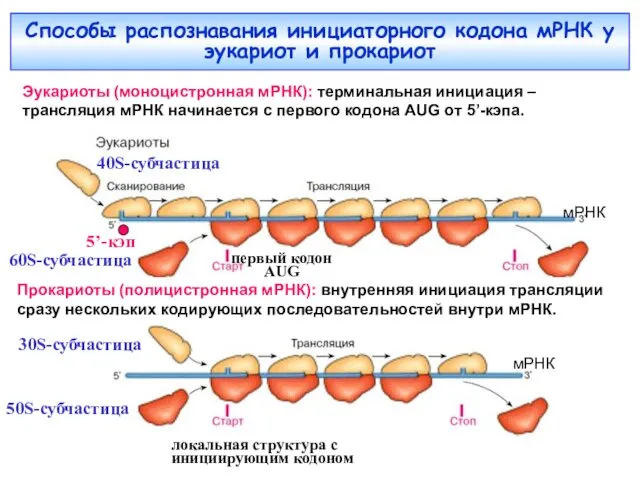
Способы распознавания инициаторного кодона мРНК у эукариот и прокариот
Эукариоты (моноцистронная мРНК):
Способы распознавания инициаторного кодона мРНК у эукариот и прокариот
Эукариоты (моноцистронная мРНК):
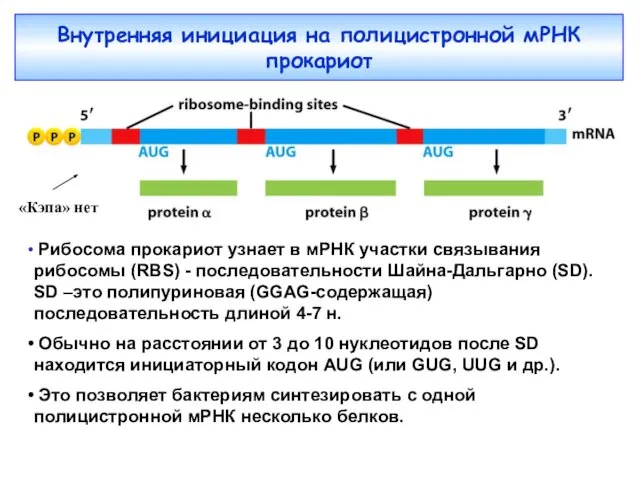
Внутренняя инициация на полицистронной мРНК прокариот
Рибосома прокариот узнает в
Внутренняя инициация на полицистронной мРНК прокариот
Рибосома прокариот узнает в
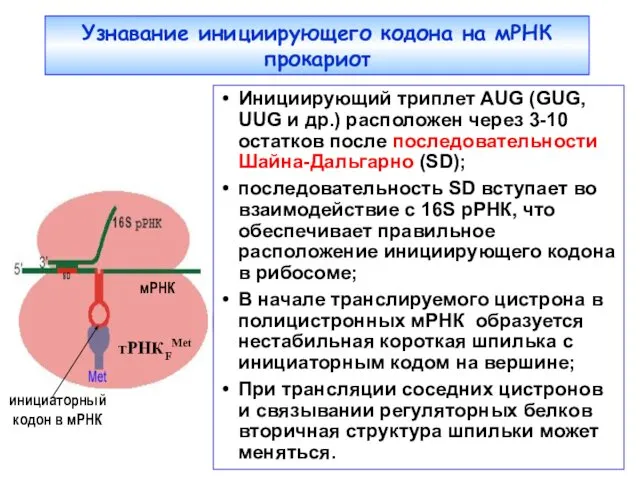
Узнавание инициирующего кодона на мРНК прокариот
Инициирующий триплет AUG (GUG, UUG и
Узнавание инициирующего кодона на мРНК прокариот
Инициирующий триплет AUG (GUG, UUG и

«Терминальная инициация по сканирующему механизму» происходит с первого от 5’-конца триплета
«Терминальная инициация по сканирующему механизму» происходит с первого от 5’-конца триплета
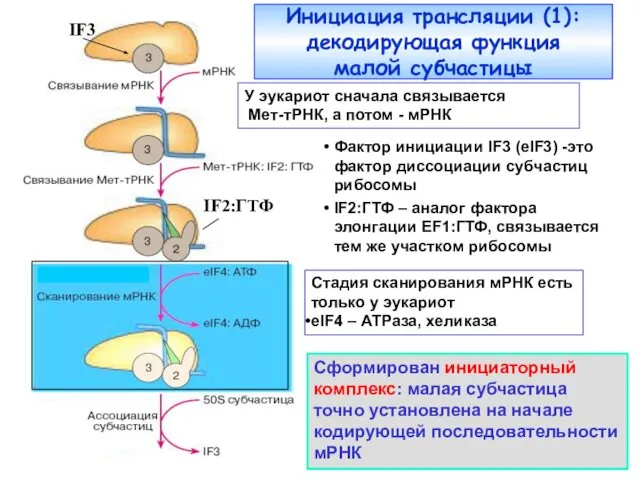
Фактор инициации IF3 (eIF3) -это фактор диссоциации субчастиц рибосомы
IF2:ГТФ – аналог
Фактор инициации IF3 (eIF3) -это фактор диссоциации субчастиц рибосомы
IF2:ГТФ – аналог
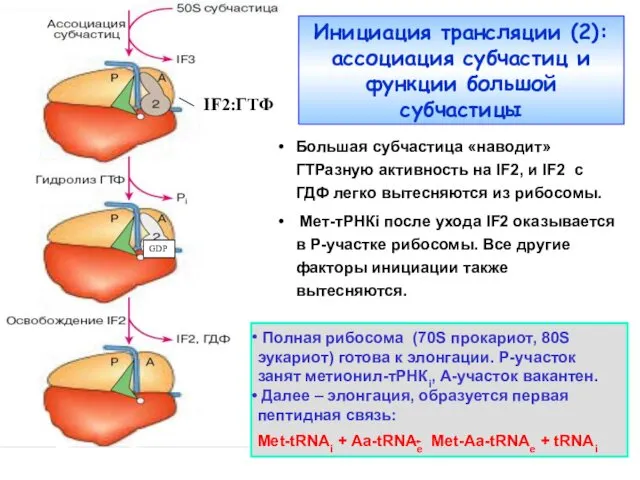
Инициация трансляции (2):
ассоциация субчастиц и функции большой субчастицы
IF2:ГТФ
Большая субчастица «наводит»
Инициация трансляции (2):
ассоциация субчастиц и функции большой субчастицы
IF2:ГТФ
Большая субчастица «наводит»
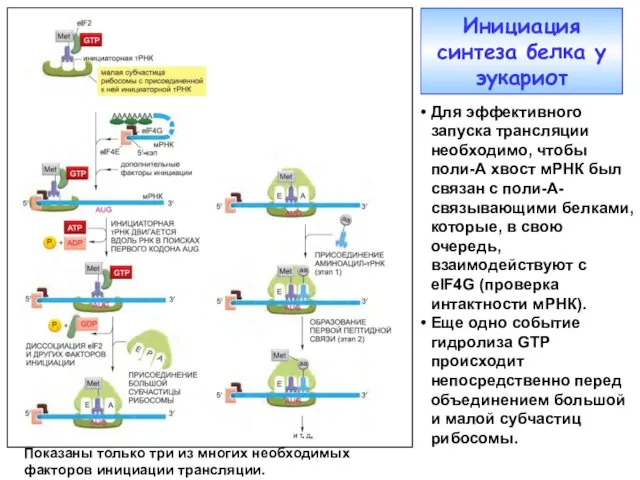
Инициация синтеза белка у эукариот
Для эффективного запуска трансляции необходимо, чтобы
Инициация синтеза белка у эукариот
Для эффективного запуска трансляции необходимо, чтобы
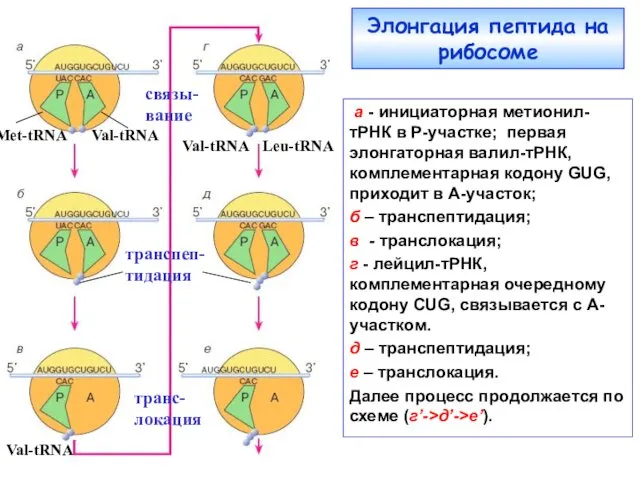
Элонгация пептида на рибосоме
Меt-tRNA
Val-tRNA
а - инициаторная метионил-тРНК в Р-участке; первая
Элонгация пептида на рибосоме
Меt-tRNA
Val-tRNA
а - инициаторная метионил-тРНК в Р-участке; первая
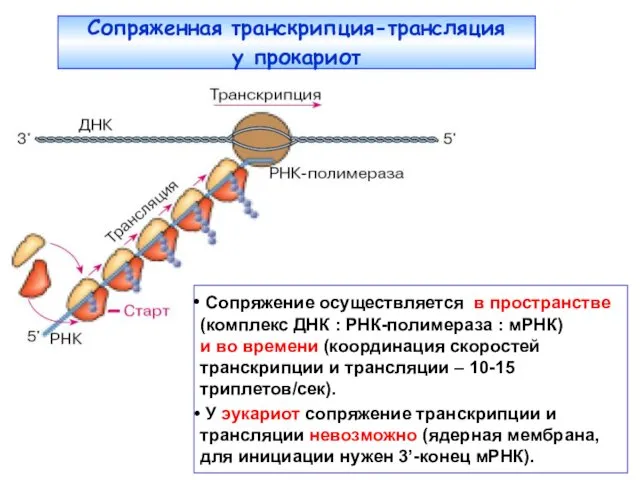
Сопряженная транскрипция-трансляция
у прокариот
Сопряжение осуществляется в пространстве (комплекс ДНК
Сопряженная транскрипция-трансляция
у прокариот
Сопряжение осуществляется в пространстве (комплекс ДНК
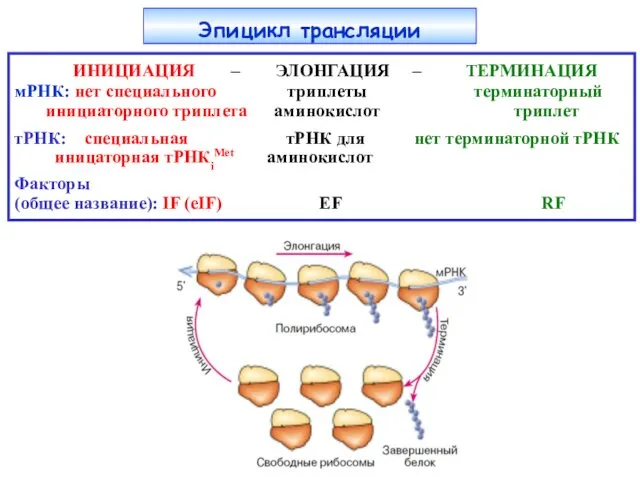
Эпицикл трансляции
ИНИЦИАЦИЯ – ЭЛОНГАЦИЯ – ТЕРМИНАЦИЯ
мРНК: нет специального триплеты
Эпицикл трансляции
ИНИЦИАЦИЯ – ЭЛОНГАЦИЯ – ТЕРМИНАЦИЯ
мРНК: нет специального триплеты

Терминация трансляции
Стадия терминации трансляции может рассматриваться как модифицированный элонгационный цикл, где
Терминация трансляции
Стадия терминации трансляции может рассматриваться как модифицированный элонгационный цикл, где
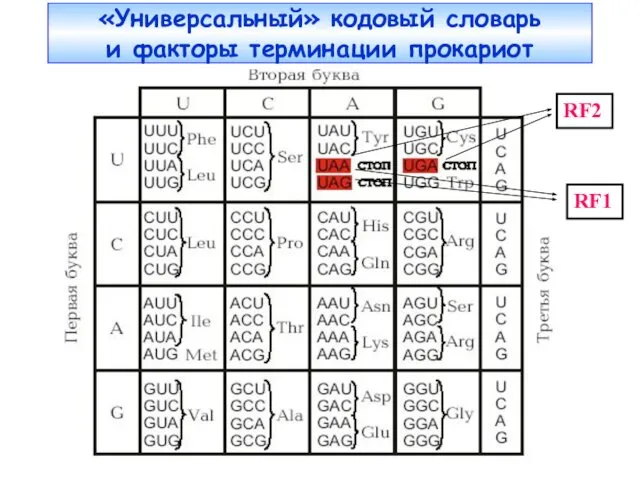
«Универсальный» кодовый словарь
и факторы терминации прокариот
RF1
RF2
«Универсальный» кодовый словарь
и факторы терминации прокариот
RF1
RF2
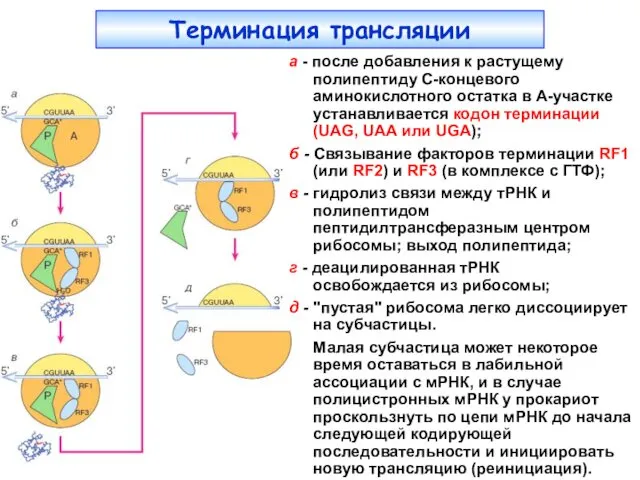
Терминация трансляции
а - после добавления к растущему полипептиду С-концевого аминокислотного остатка
Терминация трансляции
а - после добавления к растущему полипептиду С-концевого аминокислотного остатка
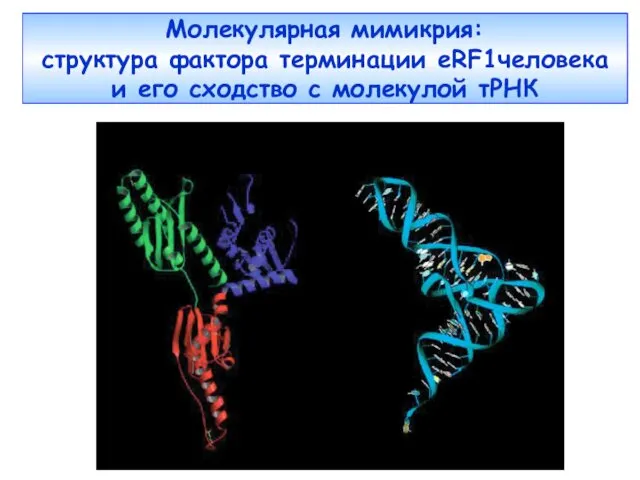
Молекулярная мимикрия:
структура фактора терминации еRF1человека
и его сходство с молекулой
Молекулярная мимикрия: структура фактора терминации еRF1человека и его сходство с молекулой

«Портрет» большой рибосомной субчастицы бактерий
белки
рРНК
Со стороны малой субчастицы С другой строны
«Портрет» большой рибосомной субчастицы бактерий
белки
рРНК
Со стороны малой субчастицы С другой строны
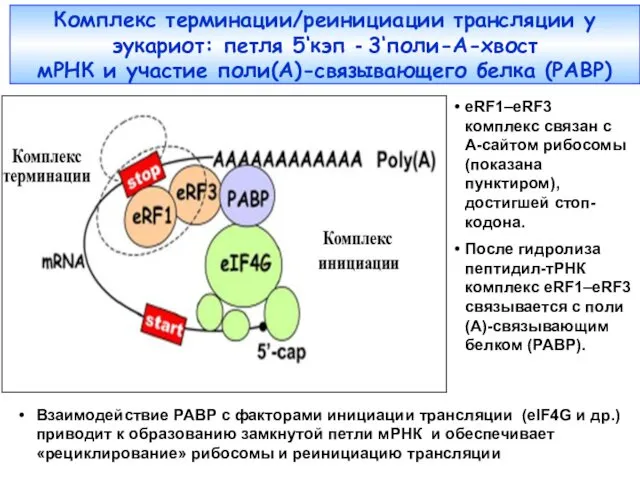
Комплекс терминации/реинициации трансляции у эукариот: петля 5‘кэп - 3‘поли-A-хвост
мРНК и участие
Комплекс терминации/реинициации трансляции у эукариот: петля 5‘кэп - 3‘поли-A-хвост мРНК и участие
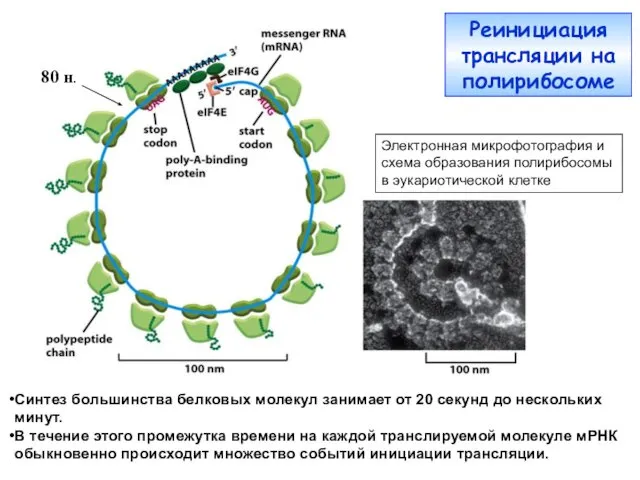
Синтез большинства белковых молекул занимает от 20 секунд до нескольких минут.
В
Синтез большинства белковых молекул занимает от 20 секунд до нескольких минут.
В

Стадии инициации и терминации – это модифицированный элонгационный цикл
При инициации Met-tRNAfMet:IF2:GTP
Стадии инициации и терминации – это модифицированный элонгационный цикл
При инициации Met-tRNAfMet:IF2:GTP

Транспортно-матричная РНК (тмРНК)
Транспортно-матричная РНК (тмРНК) содержит элементы как мРНК, так и
Транспортно-матричная РНК (тмРНК)
Транспортно-матричная РНК (тмРНК) содержит элементы как мРНК, так и
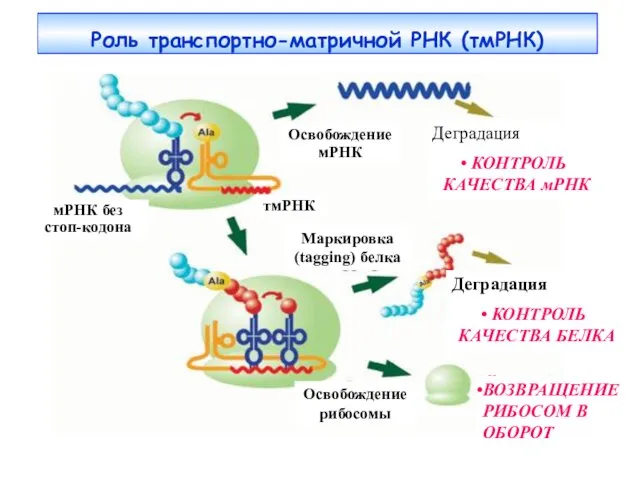
Роль транспортно-матричной РНК (тмРНК)
Деградация
КОНТРОЛЬ КАЧЕСТВА мРНК
Деградация
Роль транспортно-матричной РНК (тмРНК)
Деградация
КОНТРОЛЬ КАЧЕСТВА мРНК
Деградация

Регуляция трансляции
Регуляция осуществляется путем изменения интенсивности трансляции на данной мРНК (в
Регуляция трансляции
Регуляция осуществляется путем изменения интенсивности трансляции на данной мРНК (в
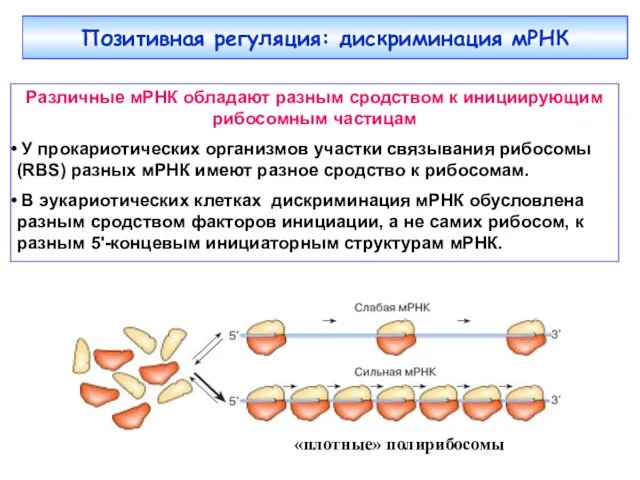
Позитивная регуляция: дискриминация мРНК
Различные мРНК обладают разным сродством к инициирующим рибосомным
Позитивная регуляция: дискриминация мРНК
Различные мРНК обладают разным сродством к инициирующим рибосомным

«Сила» мРНК в значительной мере определяет соотношение продукции
различных белков в
«Сила» мРНК в значительной мере определяет соотношение продукции различных белков в

Трансляционное сопряжение у прокариот
В полицистронных мРНК работает механизм внутренней инициации, и
Трансляционное сопряжение у прокариот
В полицистронных мРНК работает механизм внутренней инициации, и
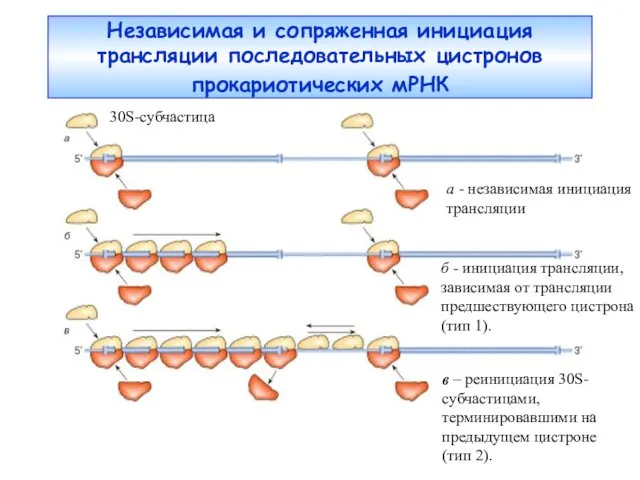
Независимая и сопряженная инициация трансляции последовательных цистронов прокариотических мРНК
а -
Независимая и сопряженная инициация трансляции последовательных цистронов прокариотических мРНК
а -
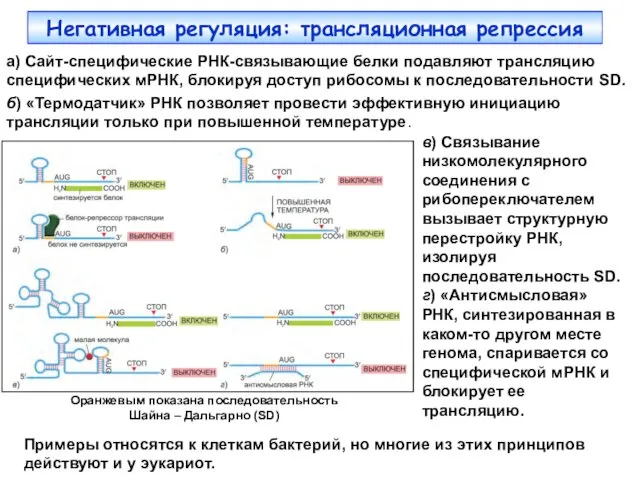
Негативная регуляция: трансляционная репрессия
а) Сайт-специфические РНК-связывающие белки подавляют трансляцию специфических мРНК,
Негативная регуляция: трансляционная репрессия
а) Сайт-специфические РНК-связывающие белки подавляют трансляцию специфических мРНК,
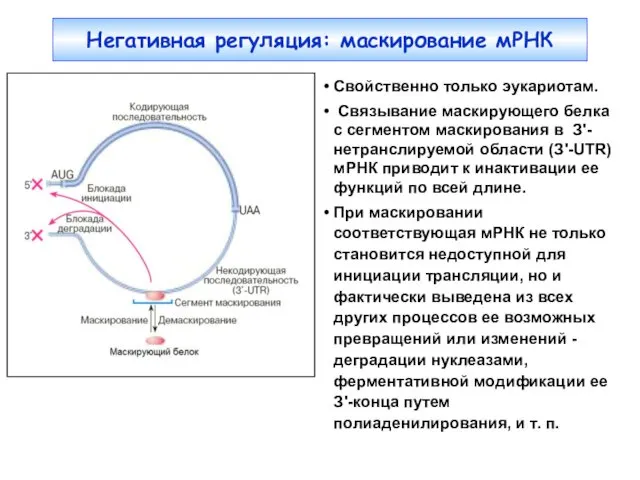
Свойственно только эукариотам.
Связывание маскирующего белка с сегментом маскирования в З'-нетранслируемой
Свойственно только эукариотам.
Связывание маскирующего белка с сегментом маскирования в З'-нетранслируемой
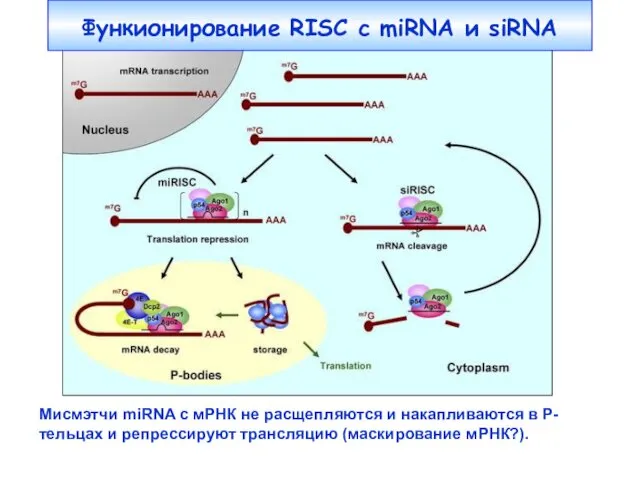
Функионирование RISC c miRNA и siRNA
Мисмэтчи miRNA с мРНК не
Функионирование RISC c miRNA и siRNA
Мисмэтчи miRNA с мРНК не

Маскирование мРНК у эукариот
Маскирование и демаскирование мРНК являются особенно характерными для
Маскирование мРНК у эукариот
Маскирование и демаскирование мРНК являются особенно характерными для
 Память. Диагностика и развитие памяти
Память. Диагностика и развитие памяти  Презентация на тему Земноводные
Презентация на тему Земноводные  Презентация на тему Опора и движение
Презентация на тему Опора и движение Защита сельскохозяйственных растений от вредителей и болезней
Защита сельскохозяйственных растений от вредителей и болезней Старость как этап онтогенеза
Старость как этап онтогенеза Живородящие птицы
Живородящие птицы Изменчивость. Готовимся к ЕГЭ
Изменчивость. Готовимся к ЕГЭ Тестовые задания по биологии Тема тестирования: «Человек как биологический вид. Общий обзор строения и функций организма че
Тестовые задания по биологии Тема тестирования: «Человек как биологический вид. Общий обзор строения и функций организма че Строение тела человека
Строение тела человека Методы и задачи селекции
Методы и задачи селекции Разнообразие животных
Разнообразие животных Удивительные способности земноводных
Удивительные способности земноводных Презентация Почвы
Презентация Почвы Сон. Фізіологія сну
Сон. Фізіологія сну Растения на школьном участке
Растения на школьном участке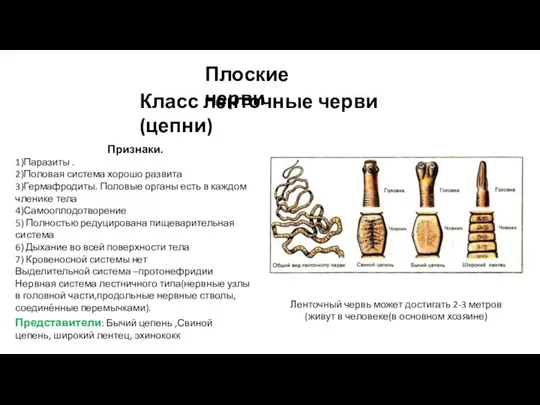 Плоские черви
Плоские черви Вегетативная нервная система
Вегетативная нервная система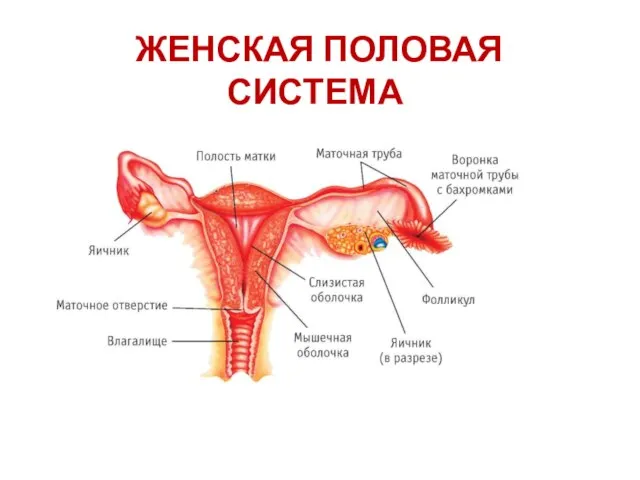 Женская половая система
Женская половая система Конечный мозг: обонятельный мозг, базальные ядра, белое веществ
Конечный мозг: обонятельный мозг, базальные ядра, белое веществ Обобщение по теме Эволюционная теория
Обобщение по теме Эволюционная теория Лось – обитатель наших лесов
Лось – обитатель наших лесов Блуждающий нерв (п. vagus). X пара черепно- мозговых нервов
Блуждающий нерв (п. vagus). X пара черепно- мозговых нервов Семейство Астровые или Сложноцветые (Asteraceae или Compositae)
Семейство Астровые или Сложноцветые (Asteraceae или Compositae)  Коммуникационные сети
Коммуникационные сети Проект Тайны куриного яйца
Проект Тайны куриного яйца Влияние кошки на жизнь человека
Влияние кошки на жизнь человека Жизнь в океанах и морях
Жизнь в океанах и морях Презентация по теме: (предмет – биология, 8 класс) Карпова Вера Михайловна, учитель биологии МОУ «Киреевская гимназия» 2007
Презентация по теме: (предмет – биология, 8 класс) Карпова Вера Михайловна, учитель биологии МОУ «Киреевская гимназия» 2007