- Эндокринология. Общие свойства гормонов

Содержание
- 3. Гормоны-определение Гормоны - это биологически активные вещества, которые синтезируются в малых количествах в специализированнных клетках эндокринной
- 4. Информация структуры Информация действия СТРУКТУРА ИНФОРМАЦИЯ структуры (гормоны) ЭНЕРГИЯ СТРУКТУРЫ ФУНКЦИЯ ИНФОРМАЦИЯ ДЕЙСТВИЯ (свет, ЭМП)
- 5. Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами. выделяются из вырабатывающих их клеток во
- 6. Гормональная регуляция — пример эволюционно наиболее древнего химического дистантного и локального контроля. Термин гормон введен Р.
- 7. Во второй половине XX века было показано, что в гипоталамической области мозга существуют клетки способные генерировать
- 8. Оказалось, что нейроэндокринные клетки мозга и клетки иммунной системы функционируют в тесной кооперации и проявляют признаки
- 9. Было показано, что в ответ на действие патогенных агентов из макрофагов выделяется ИЛ-1. Этот иммунный пептид
- 10. Взаимодействия нейроэндокринной и иммунной систем в различные периоды жизни изменяются. С возрастом снижаются реакции иммунной защиты
- 11. Варианты взаимодействия специфических регуляторных систем Нервная система мембранные рецепторы = ионные каналы изменение мембранного потенциала Эндокринная
- 12. Эндокринная система организма животных и человека включает комплекс центральных и периферических эндокринных желез, а также другие
- 13. Особенности группирования разных типов эндокринных клеток Эндокринные клетки не эндокринных органов (энтероциты ЖКТ, эндокринные нейроны гипоталамуса)
- 14. Гормоны, секретируемые эффекторными типами клеток Клетки печени ИФР-1 Ангиотензиноген (прогормон) Клетки почек Эритропоэтин Клетки жировой ткани
- 15. Эндокринной активностью обладает также плацента. На разных этапах беременности её значение и активность существенно меняются.
- 16. Изменение гормонов во время беременности
- 17. Гормональные изменения во время беременности
- 18. Биологический смысл интеграции эндокринных клеток в эндокринную железу – создание более совершенного механизма управления потоками информации
- 19. Существуют три основных класса гормонов по химическому строению 1. Белки и полипептиды, включая гормоны, секретируемые передней
- 20. По свойствам и механизму действия все гормоны можно разделить на 2 группы 1) гидрофильные (растворяющиеся в
- 21. Гормональные эффекты срочные реакции - изменение проницаемости мембраны для ионов или глюкозы и аминокислот, что приводит,
- 22. Общие свойства гормонов: Строгая специфичность действия. Высокая биологическая активность. Дистантный характер действия. Многие гормоны не имеют
- 23. Этапы гуморальной регуляции Место выработки Стимул для выделения Транспорт Рецепция и клеточные механизмы действия Физиологические эффекты
- 24. Место выработки Эндокринная железа Неэндокринные органы Одно или несколько мест выработки
- 25. Стимул для выделения 1.Нервный импульс 2. Влияние другого гормона 3. Конкретное вещество, уровень которого регулируется гормоном
- 26. Дистантность действия гормона ГОРМОН Дистантные эффекты Паракринные эффекты Аутокринные и юкстакринные эффекты Внутриклеточные эффекты Системное действие
- 27. Типы местного контроля Внутриклеточный контроль Региональный контроль Аутокринный Юкстакринный Паракринный С Р Э Э Э Э
- 28. Типы системного контроля Эндокринный Нейроэндокринный Нейрокринный Г Э1 Э3 Э2 НГ Р Р Р Р Р
- 29. Транспорт Транспорта нет – паракринный эффект Транспорт со специальным белком Транспорт в неактивной или малоактивной форме
- 30. Транспорт Водорастворимые гормоны растворены в плазме. Стероидные, тиреоидные в связи со специфическими белками переносчиками, но около
- 31. Метаболизм гормонов Метаболизм в тканях с изменением активности Связывание в тканях Разрушение в лизосомах Катаболизм в
- 32. Рецепция, клеточные и молекулярные механизмы действия
- 33. Орган или ткань мишень Один Несколько Ответ тканей-мишеней формируется при образовании гормон-рецепторного комплекса.
- 34. Регуляция рецепции Количество рецепторов Чувствительность рецепторов Индуцибельность рецепторов Типы рецепторов (мембранные, цитоплазматические, ядерные)
- 35. Механизмы рецепции стероидных гормонов
- 36. Стероидные соединения плохо растворяются в воде и хорошо в органических растворителях и растительных маслах. Благодаря липофильности
- 37. Рецепция и метаболизм жирорастворимых гормонов
- 38. Механизмы рецепции гормонов белковой природы
- 39. Клеточные рецепторы делятся на следующие классы мембранные рецепторы, сопряжённые с G-белками рецепторные тирозинкиназы ионные каналы. цитоплазматические
- 40. Значение вторичных посредников для рецепции белковых типов гормонов Эффективность действия малых доз гормонов Универсальный ответ клетки
- 41. Вторичные посредники
- 43. Вторичные посредникиВторичные посредники (англ. second messenger) — это низкомолекулярные вещества, которые образуются или высвобождаются в результате
- 44. Вторичные посредники ионы кальция (Ca2+); циклический аденозинмонофосфат (цАМФциклический аденозинмонофосфат (цАМФ) и циклический гуанозинмонофосфат (цГМФ). инозитолтрифосфат липофильные
- 45. Классификация вторичных посредников по растворимости в воде и размеру молекулы ГидрофобныеГидрофобные молекулы — нерастворимые в воде
- 46. Аденилатциклазная система
- 47. Аденилатциклазная система рассматривается на примере действия адреналинаАденилатциклазная система рассматривается на примере действия адреналина на клетки печени.
- 48. Особенность этой системы передачи сигнала в клетке Сигнал на большинстве этапов усиливается, например, активированная аденилатциклаза синтезирует
- 49. Таким образом, даже относительно слабый стимул может вызывать значительный ответ. Это явление называется амплификацией сигнала
- 50. Роль кальция как вторичного посредника. Кальмодулин. Кальций-кальмодулин-зависимые киназы.
- 51. Выделяют шесть типов потенциалзависимых кальциевых каналов. Наиболее важными в сердечно-сосудистой системе являются L- и Т-типы. Каналы
- 52. Два типа передачи сигнала при посредстве ионов кальция (1) Первый из них осуществляется в электро-возбудимых, преимущественно
- 53. Два типа передачи сигнала при посредстве ионов кальция (2) Второй способ передачи сигнала при посредстве ионов
- 55. Производные арахидоновой кислоты
- 56. Арахидоновая кислота Существует два основных пути метаболизма арахидоновой кислоты - циклоксигеназныйСуществует два основных пути метаболизма арахидоновой
- 57. ТБ - тромбоксан, ЛТ -лейкотриен, ГПЭТЕ - гидропероксиэйкозатетраеновая кислота, ПГ -простагландин
- 58. Роль простагландинов:
- 59. Тирозинкиназные рецепторы
- 60. Рецепторы с тирозинкиназной активностью Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки по тирозину, подразделяют на 2
- 61. Рецептор состоит из двух α- и двух β-субъединиц, связанных дисульфидными связями и нековалентными взаимодействиями. α- и
- 64. Скачать презентацию


Гормоны-определение
Гормоны - это биологически активные вещества, которые синтезируются в малых количествах
Гормоны-определение
Гормоны - это биологически активные вещества, которые синтезируются в малых количествах

Информация структуры
Информация действия
СТРУКТУРА
ИНФОРМАЦИЯ структуры
(гормоны)
ЭНЕРГИЯ СТРУКТУРЫ
ФУНКЦИЯ
ИНФОРМАЦИЯ ДЕЙСТВИЯ
(свет, ЭМП)
Информация структуры
Информация действия
СТРУКТУРА
ИНФОРМАЦИЯ структуры
(гормоны)
ЭНЕРГИЯ СТРУКТУРЫ
ФУНКЦИЯ
ИНФОРМАЦИЯ ДЕЙСТВИЯ
(свет, ЭМП)

Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами.
Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами.

Гормональная регуляция — пример эволюционно наиболее древнего химического дистантного и локального
Гормональная регуляция — пример эволюционно наиболее древнего химического дистантного и локального

Во второй половине XX века было показано, что в гипоталамической области
Во второй половине XX века было показано, что в гипоталамической области

Оказалось, что нейроэндокринные клетки мозга и клетки иммунной системы функционируют в
Оказалось, что нейроэндокринные клетки мозга и клетки иммунной системы функционируют в

Было показано, что в ответ на действие патогенных агентов из макрофагов
Было показано, что в ответ на действие патогенных агентов из макрофагов

Взаимодействия нейроэндокринной и иммунной систем в различные периоды жизни изменяются.
С
Взаимодействия нейроэндокринной и иммунной систем в различные периоды жизни изменяются.
С
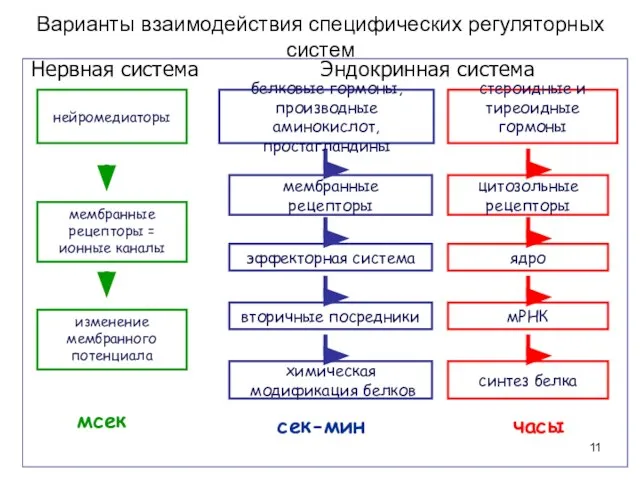
Варианты взаимодействия специфических регуляторных систем
Нервная система
мембранные
рецепторы =
ионные каналы
изменение мембранного потенциала
Эндокринная система
сек-мин
часы
мсек
белковые
Варианты взаимодействия специфических регуляторных систем
Нервная система
мембранные
рецепторы =
ионные каналы
изменение мембранного потенциала
Эндокринная система
сек-мин
часы
мсек
белковые

Эндокринная система организма животных и человека включает комплекс центральных и периферических
Эндокринная система организма животных и человека включает комплекс центральных и периферических

Особенности группирования разных типов эндокринных клеток
Эндокринные клетки не эндокринных органов (энтероциты
Особенности группирования разных типов эндокринных клеток
Эндокринные клетки не эндокринных органов (энтероциты

Гормоны, секретируемые эффекторными типами клеток
Клетки печени
ИФР-1
Ангиотензиноген (прогормон)
Клетки почек
Эритропоэтин
Клетки жировой ткани
Лептин
Адипонектин
Кардиомиоциты
Атриопептиды
Гормоны, секретируемые эффекторными типами клеток
Клетки печени
ИФР-1
Ангиотензиноген (прогормон)
Клетки почек
Эритропоэтин
Клетки жировой ткани
Лептин
Адипонектин
Кардиомиоциты
Атриопептиды

Эндокринной активностью обладает также плацента.
На разных этапах беременности её значение и
Эндокринной активностью обладает также плацента.
На разных этапах беременности её значение и

Изменение гормонов во время беременности
Изменение гормонов во время беременности
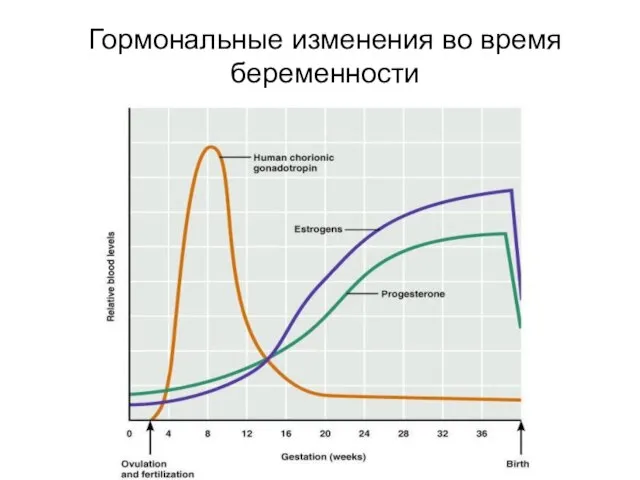
Гормональные изменения во время беременности
Гормональные изменения во время беременности

Биологический смысл интеграции эндокринных клеток в эндокринную железу –
создание
создание

Существуют три основных класса гормонов по химическому строению
1. Белки и
Существуют три основных класса гормонов по химическому строению 1. Белки и

По свойствам и механизму действия
все гормоны можно разделить на 2 группы
1)
По свойствам и механизму действия
все гормоны можно разделить на 2 группы
1)

Гормональные эффекты
срочные реакции - изменение проницаемости мембраны для ионов или глюкозы
Гормональные эффекты
срочные реакции - изменение проницаемости мембраны для ионов или глюкозы

Общие свойства гормонов:
Строгая специфичность действия.
Высокая биологическая активность.
Дистантный характер действия.
Многие
Общие свойства гормонов:
Строгая специфичность действия.
Высокая биологическая активность.
Дистантный характер действия.
Многие

Этапы гуморальной регуляции
Место выработки
Стимул для выделения
Транспорт
Рецепция и клеточные механизмы действия
Физиологические эффекты
Метаболизм
Этапы гуморальной регуляции
Место выработки
Стимул для выделения
Транспорт
Рецепция и клеточные механизмы действия
Физиологические эффекты
Метаболизм

Место выработки
Эндокринная железа
Неэндокринные органы
Одно или несколько мест выработки
Место выработки
Эндокринная железа
Неэндокринные органы
Одно или несколько мест выработки

Стимул для выделения
1.Нервный импульс
2. Влияние другого гормона
3. Конкретное вещество,
Стимул для выделения
1.Нервный импульс
2. Влияние другого гормона
3. Конкретное вещество,
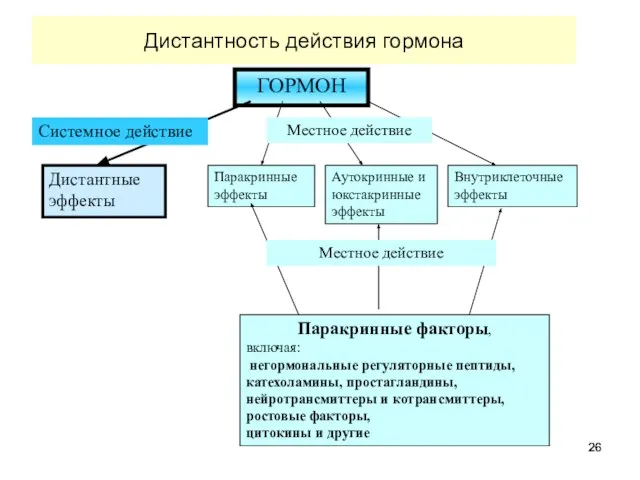
Дистантность действия гормона
ГОРМОН
Дистантные эффекты
Паракринные эффекты
Аутокринные и юкстакринные эффекты
Внутриклеточные эффекты
Системное действие
Местное действие
Паракринные
Дистантность действия гормона
ГОРМОН
Дистантные эффекты
Паракринные эффекты
Аутокринные и юкстакринные эффекты
Внутриклеточные эффекты
Системное действие
Местное действие
Паракринные

Типы местного контроля
Внутриклеточный контроль
Региональный контроль
Аутокринный Юкстакринный Паракринный
С Р
Э
Типы местного контроля
Внутриклеточный контроль
Региональный контроль
Аутокринный Юкстакринный Паракринный
С Р
Э
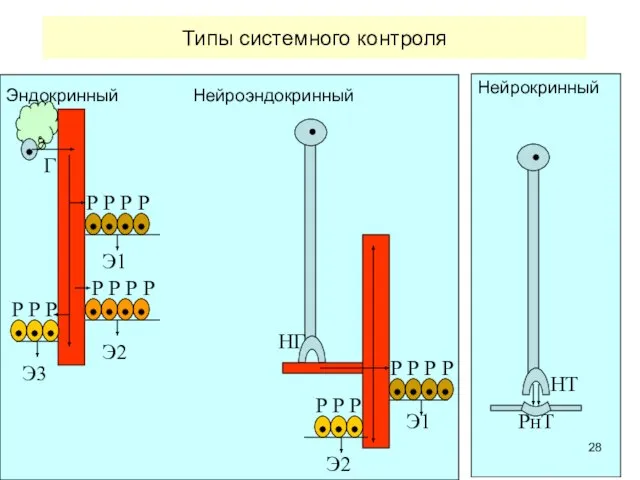
Типы системного контроля
Эндокринный Нейроэндокринный
Нейрокринный
Г
Э1
Э3
Э2
НГ
Р Р Р Р
Р Р Р Р
Р
Типы системного контроля
Эндокринный Нейроэндокринный
Нейрокринный
Г
Э1
Э3
Э2
НГ
Р Р Р Р
Р Р Р Р
Р

Транспорт
Транспорта нет – паракринный эффект
Транспорт со специальным белком
Транспорт в неактивной или
Транспорт
Транспорта нет – паракринный эффект
Транспорт со специальным белком
Транспорт в неактивной или

Транспорт
Водорастворимые гормоны растворены в плазме.
Стероидные, тиреоидные в связи со специфическими белками
Транспорт
Водорастворимые гормоны растворены в плазме.
Стероидные, тиреоидные в связи со специфическими белками

Метаболизм гормонов
Метаболизм в тканях с изменением активности
Связывание в тканях
Разрушение в лизосомах
Катаболизм
Метаболизм гормонов
Метаболизм в тканях с изменением активности
Связывание в тканях
Разрушение в лизосомах
Катаболизм

Рецепция, клеточные и молекулярные механизмы действия
Рецепция, клеточные и молекулярные механизмы действия

Орган
или ткань мишень
Один
Несколько
Ответ тканей-мишеней формируется при образовании гормон-рецепторного
Орган
или ткань мишень
Один
Несколько
Ответ тканей-мишеней формируется при образовании гормон-рецепторного

Регуляция рецепции
Количество рецепторов
Чувствительность рецепторов
Индуцибельность рецепторов
Типы рецепторов (мембранные, цитоплазматические, ядерные)
Регуляция рецепции
Количество рецепторов
Чувствительность рецепторов
Индуцибельность рецепторов
Типы рецепторов (мембранные, цитоплазматические, ядерные)

Механизмы рецепции стероидных гормонов
Механизмы рецепции стероидных гормонов

Стероидные соединения плохо растворяются в воде и хорошо в органических растворителях
Стероидные соединения плохо растворяются в воде и хорошо в органических растворителях

Рецепция и метаболизм жирорастворимых гормонов
Рецепция и метаболизм жирорастворимых гормонов

Механизмы рецепции гормонов белковой природы
Механизмы рецепции гормонов белковой природы

Клеточные рецепторы делятся на следующие классы
мембранные
рецепторы, сопряжённые с G-белками
рецепторные
Клеточные рецепторы делятся на следующие классы
мембранные
рецепторы, сопряжённые с G-белками
рецепторные

Значение вторичных посредников для рецепции белковых типов гормонов
Эффективность действия малых доз
Значение вторичных посредников для рецепции белковых типов гормонов
Эффективность действия малых доз

Вторичные посредники
Вторичные посредники


Вторичные посредникиВторичные посредники (англ. second messenger) — это низкомолекулярные вещества, которые образуются
Вторичные посредникиВторичные посредники (англ. second messenger) — это низкомолекулярные вещества, которые образуются

Вторичные посредники
ионы кальция (Ca2+);
циклический аденозинмонофосфат (цАМФциклический аденозинмонофосфат (цАМФ) и циклический
Вторичные посредники
ионы кальция (Ca2+);
циклический аденозинмонофосфат (цАМФциклический аденозинмонофосфат (цАМФ) и циклический

Классификация вторичных посредников по растворимости в воде и размеру молекулы
ГидрофобныеГидрофобные молекулы —
Классификация вторичных посредников по растворимости в воде и размеру молекулы
ГидрофобныеГидрофобные молекулы —

Аденилатциклазная система
Аденилатциклазная система

Аденилатциклазная система рассматривается на примере действия адреналинаАденилатциклазная система рассматривается на примере
Аденилатциклазная система рассматривается на примере действия адреналинаАденилатциклазная система рассматривается на примере

Особенность этой системы передачи сигнала в клетке
Сигнал на большинстве этапов
Особенность этой системы передачи сигнала в клетке
Сигнал на большинстве этапов

Таким образом, даже относительно слабый стимул может вызывать значительный ответ.
Это
Таким образом, даже относительно слабый стимул может вызывать значительный ответ.
Это

Роль кальция как вторичного посредника. Кальмодулин. Кальций-кальмодулин-зависимые киназы.
Роль кальция как вторичного посредника. Кальмодулин. Кальций-кальмодулин-зависимые киназы.

Выделяют шесть типов потенциалзависимых кальциевых каналов.
Наиболее важными в сердечно-сосудистой системе
Выделяют шесть типов потенциалзависимых кальциевых каналов.
Наиболее важными в сердечно-сосудистой системе

Два типа передачи сигнала при посредстве ионов кальция (1)
Первый из них
Два типа передачи сигнала при посредстве ионов кальция (1)
Первый из них

Два типа передачи сигнала при посредстве ионов кальция (2)
Второй способ передачи
Два типа передачи сигнала при посредстве ионов кальция (2)
Второй способ передачи


Производные арахидоновой кислоты
Производные арахидоновой кислоты

Арахидоновая кислота
Существует два основных пути метаболизма арахидоновой кислоты - циклоксигеназныйСуществует два
Арахидоновая кислота
Существует два основных пути метаболизма арахидоновой кислоты - циклоксигеназныйСуществует два

ТБ - тромбоксан,
ЛТ -лейкотриен,
ГПЭТЕ - гидропероксиэйкозатетраеновая кислота,
ПГ -простагландин
ТБ - тромбоксан,
ЛТ -лейкотриен,
ГПЭТЕ - гидропероксиэйкозатетраеновая кислота,
ПГ -простагландин
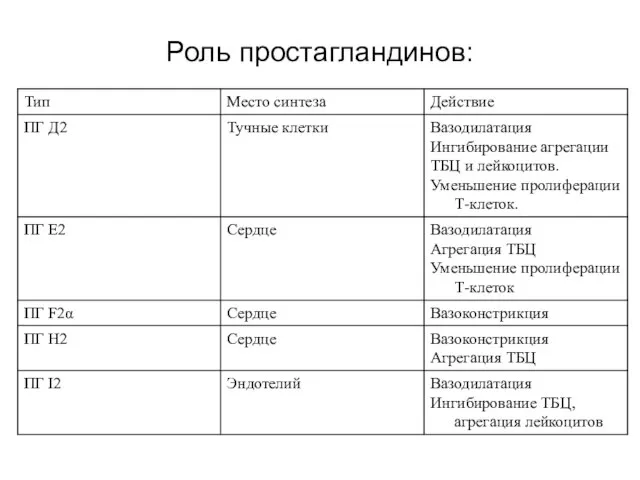
Роль простагландинов:
Роль простагландинов:

Тирозинкиназные рецепторы
Тирозинкиназные рецепторы

Рецепторы с тирозинкиназной активностью
Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки
Рецепторы с тирозинкиназной активностью
Тирозиновые протеинкиназы - ферменты, фосфорилирующие специфические белки

Рецептор состоит из двух α- и двух β-субъединиц, связанных дисульфидными связями
Рецептор состоит из двух α- и двух β-субъединиц, связанных дисульфидными связями

 Экологические группы водорослей. Значение
Экологические группы водорослей. Значение Презентация на тему "Кожа: строение и значение" - скачать бесплатно презентации по Биологии
Презентация на тему "Кожа: строение и значение" - скачать бесплатно презентации по Биологии Опорно-двигательная система
Опорно-двигательная система Типы биотических связей в биоценозе
Типы биотических связей в биоценозе Презентация на тему Класс Земноводные
Презентация на тему Класс Земноводные  Кожа – наружный покровный орган
Кожа – наружный покровный орган Зеленая аптека Перелюбского района. Исследовательская работа ученика МОУ «СОШ имени М.М. Рудченко с.Перелюб» Бондарева Алексея
Зеленая аптека Перелюбского района. Исследовательская работа ученика МОУ «СОШ имени М.М. Рудченко с.Перелюб» Бондарева Алексея Культуры клеток, тканей и органов растений как основа БТ растений. Тотипотентность растительной клетки (лекция 6)
Культуры клеток, тканей и органов растений как основа БТ растений. Тотипотентность растительной клетки (лекция 6) Синтетическая теория эволюции
Синтетическая теория эволюции Русская выхухоль имеет право на жизнь
Русская выхухоль имеет право на жизнь Презентация на тему Улитки
Презентация на тему Улитки История изучения клетки. Клеточная теория
История изучения клетки. Клеточная теория Презентация на тему "Класс млекопитающие или звери" - скачать презентации по Биологии
Презентация на тему "Класс млекопитающие или звери" - скачать презентации по Биологии Лучше один раз увидеть?
Лучше один раз увидеть? Пищеварение в желудке Автор: Ледкова Марина Борисовна учитель биологии МОУ гимназия № 2 2008 год
Пищеварение в желудке Автор: Ледкова Марина Борисовна учитель биологии МОУ гимназия № 2 2008 год Закон гомологических рядов Вавилова
Закон гомологических рядов Вавилова Дары рек и озёр. 3 класс
Дары рек и озёр. 3 класс Совы
Совы Презентация на тему "Скелет" - скачать бесплатно презентации по Биологии
Презентация на тему "Скелет" - скачать бесплатно презентации по Биологии Презентация на тему "Полевые сорняки" - скачать презентации по Биологии
Презентация на тему "Полевые сорняки" - скачать презентации по Биологии Отряд грызунов
Отряд грызунов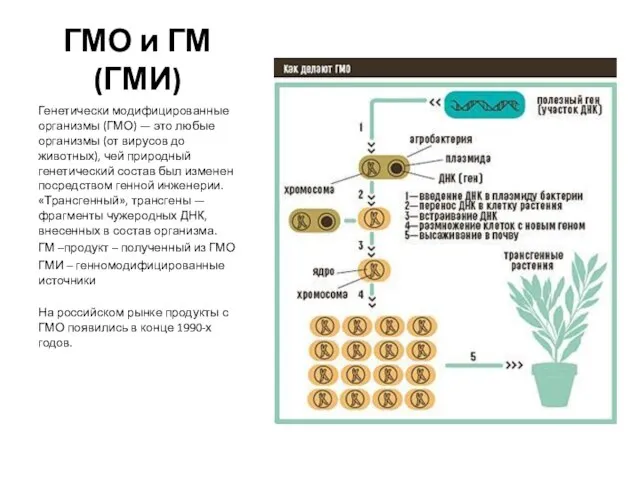 ГМО и ГМ (ГМИ) Генетически модифицированные организмы (ГМО) — это любые организмы (от вирусов до животных), чей природный генетичес
ГМО и ГМ (ГМИ) Генетически модифицированные организмы (ГМО) — это любые организмы (от вирусов до животных), чей природный генетичес Филогенез скелета позвоночных животных
Филогенез скелета позвоночных животных Организм человека. Введение
Организм человека. Введение Физиология кровообращения
Физиология кровообращения Репродуктивная система мужчины. Внутренние половые органы
Репродуктивная система мужчины. Внутренние половые органы Тема «Внутреннее строение рыбы» Тыквенко Наталья Вениаминовна Учитель географии и биологии Гляденской ООШ №11 Красноярский край
Тема «Внутреннее строение рыбы» Тыквенко Наталья Вениаминовна Учитель географии и биологии Гляденской ООШ №11 Красноярский край Шлюбна поведінка птахів та улаштування гнізд
Шлюбна поведінка птахів та улаштування гнізд