- Энергетический обмен. (Лекция 4)

Содержание
- 2. Метаболизм- система строго регулируемых реакций в клетке, обеспечивающих жизнедеятельность. Метаболизм представлен двумя путями: катаболизм — система
- 3. Взаимосвязь катаболизма с анаболизмом АТФ катаболизм анаболизм
- 4. 1.Снабжение организма химической энергией, трансформация (преобразование) ее в форму АТФ или (восстановительных потенциалов- например НАДФН+); 2.Превращение
- 5. 1.Переваривание в ЖКТ(ферментативный гидролиз или деполимеризация) сложных пищевых веществ до мономеров, способных всасываться; 2.Всасывание веществ; 3.
- 6. Биоэнергетика - раздел биохимии энергетический обмен Основные вопросы: 1. Источники энергии для клетки; 2. Механизмы высвобождения
- 7. Особенности биоэнергетики Организм идеально устроен для выполнения различных видов работ. Для этого нужна энергия!!! Превращение энергии
- 8. Механизмы синтеза АТФ в клетках гетеротрофов Для живой клетки гетеротрофов характерны три механизма синтеза АТФ :
- 9. 1.Потенциальные источники энергии для клетки - термодинамически нестабильные органические вещества, главным образом, биомолекулы - углеводы, липиды,
- 10. Механизм высвобождения энергии из потенциальных источников. Высвобождение энергии из химических связей потенциальных источников энергии осуществляется в
- 11. Характерные особенности биологического окисления По химизму процесс представляет цепь окислительно-восстановительных реакций. Конкретно, реакций дегидрирования окисляемого вещества
- 12. Биологическое окисление –общая схема 1. Тканевое дыхание – конечный акцептор атмосферный кислород!!!!! SH2 ½ O2 ½
- 13. Тканевое дыхание Процесс- в митохондриях!!! Внутренняя мембрана Включает этапы: А. дегидрирование субстрата т.е. отщепление водорода (протоны
- 14. Тканевое дыхание обеспечивает: - создание градиента концентрации протонов по обе стороны внутренней мембраны митохондрии [ H+];
- 15. Схема дыхательной цепи + + + + + + + + + + + ФМНН2 ФМН
- 16. Ферменты тканевого дыхания Класс - оксидоредуктазы, подклассы - дегидрогеназы, оксидазы Коферменты этих ферментов - промежуточные переносчики
- 17. Характеристика ферментов дыхательной цепи 1. Первичные дегидрогеназы - осуществляют первичное дегидрирование первичных субстратов (SH2- промежуточный метаболит
- 18. 2. Система ферментов, сформированная в последовательную цепь переноса электронов (ЦПЭ). Все ферменты сформированы в 3 комплекса.
- 19. III. Третий ферментативный комплекс (III) –Убихинон-оксидоредуктаза. Представлен двумя типами ферментов- цитохромов В, С1. Кофермент - гем
- 20. IV. Четвертый ферментативный комплекс (IV) – цитохромоксидаза. Обладает абсолютной специфичностью к кислороду. Представлен двумя цитохромами а
- 21. Участки ( пункт) сопряжения дыхательной цепи. В дыхательной цепи выделяют три участка, в которых перенос электронов
- 22. Окислительное фосфорилирование Трансформация электрохимического потенциала в энергию макроэргической связи АТФ - происходит в процессе окислительного фосфорилирования,
- 23. Система стремится к равновесию и при достижении определенного значения градиента концентраций протонов и потенциала протоны стремятся
- 24. Коэффициент сопряжения (коэффициент окислительного фосфорилирования) Р/О коэффициент сопряжения – количественная характеристика сопряжения . Р – количество
- 25. Дыхательный контроль Интенсивность тканевого дыхания зависит от соотношения в клетке АТФ / АДФ. Образованная АТФ покидает
- 26. По механизмы действия на 3 группы: 1.Ингибиторы тканевого дыхания 2.Ингибиторы окислительного фосфорилирования 3. Разобщители тканевого дыхания
- 27. Ингибиторы тканевого дыхания Снижают создание электрохимического потенциала, прерывая поток электронов на участках дыхательной цепи, обратимо ингибируя
- 28. Ингибиторы окислительного фосфорилирования Связываются с протомерами АТФ-синтазы, запечатывая протонный канал. Прекращается синтез АТФ. Примеры: антибиотики (
- 30. Скачать презентацию

Метаболизм- система строго регулируемых реакций в клетке, обеспечивающих жизнедеятельность.
Метаболизм представлен двумя
Метаболизм- система строго регулируемых реакций в клетке, обеспечивающих жизнедеятельность. Метаболизм представлен двумя
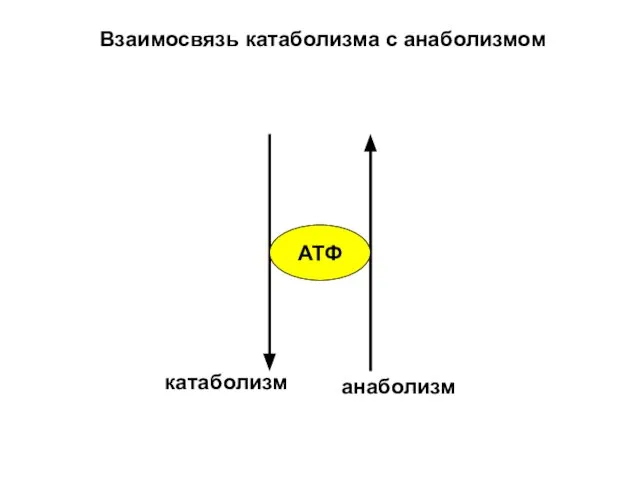
Взаимосвязь катаболизма с анаболизмом
АТФ
катаболизм
анаболизм
Взаимосвязь катаболизма с анаболизмом
АТФ
катаболизм
анаболизм

1.Снабжение организма химической энергией, трансформация (преобразование) ее в форму АТФ или
1.Снабжение организма химической энергией, трансформация (преобразование) ее в форму АТФ или

1.Переваривание в ЖКТ(ферментативный гидролиз или деполимеризация) сложных пищевых веществ до мономеров,
1.Переваривание в ЖКТ(ферментативный гидролиз или деполимеризация) сложных пищевых веществ до мономеров,

Биоэнергетика - раздел биохимии
энергетический обмен
Основные вопросы:
1. Источники энергии для
Биоэнергетика - раздел биохимии
энергетический обмен
Основные вопросы:
1. Источники энергии для

Особенности биоэнергетики
Организм идеально устроен для выполнения различных видов работ. Для
Особенности биоэнергетики
Организм идеально устроен для выполнения различных видов работ. Для

Механизмы синтеза АТФ в клетках гетеротрофов
Для живой клетки гетеротрофов характерны
Механизмы синтеза АТФ в клетках гетеротрофов
Для живой клетки гетеротрофов характерны

1.Потенциальные источники энергии для клетки - термодинамически нестабильные органические вещества, главным
1.Потенциальные источники энергии для клетки - термодинамически нестабильные органические вещества, главным

Механизм высвобождения энергии из потенциальных источников.
Высвобождение энергии из химических связей
Механизм высвобождения энергии из потенциальных источников.
Высвобождение энергии из химических связей

Характерные особенности биологического окисления
По химизму процесс представляет цепь окислительно-восстановительных реакций. Конкретно,
Характерные особенности биологического окисления
По химизму процесс представляет цепь окислительно-восстановительных реакций. Конкретно,

Биологическое окисление –общая схема
1. Тканевое дыхание – конечный акцептор атмосферный
Биологическое окисление –общая схема
1. Тканевое дыхание – конечный акцептор атмосферный

Тканевое дыхание
Процесс- в митохондриях!!! Внутренняя мембрана
Включает этапы:
А. дегидрирование субстрата
Тканевое дыхание
Процесс- в митохондриях!!! Внутренняя мембрана
Включает этапы:
А. дегидрирование субстрата

Тканевое дыхание обеспечивает:
- создание градиента концентрации протонов по обе стороны внутренней
Тканевое дыхание обеспечивает: - создание градиента концентрации протонов по обе стороны внутренней

Схема дыхательной цепи
+ + + + + + + + +
Схема дыхательной цепи
+ + + + + + + + +

Ферменты тканевого дыхания
Класс - оксидоредуктазы,
подклассы - дегидрогеназы, оксидазы
Коферменты
Ферменты тканевого дыхания
Класс - оксидоредуктазы,
подклассы - дегидрогеназы, оксидазы
Коферменты

Характеристика ферментов дыхательной цепи
1. Первичные дегидрогеназы - осуществляют первичное дегидрирование
Характеристика ферментов дыхательной цепи
1. Первичные дегидрогеназы - осуществляют первичное дегидрирование

2. Система ферментов, сформированная в последовательную цепь переноса электронов (ЦПЭ). Все
2. Система ферментов, сформированная в последовательную цепь переноса электронов (ЦПЭ). Все

III. Третий ферментативный комплекс (III) –Убихинон-оксидоредуктаза. Представлен двумя типами ферментов- цитохромов
III. Третий ферментативный комплекс (III) –Убихинон-оксидоредуктаза. Представлен двумя типами ферментов- цитохромов

IV. Четвертый ферментативный комплекс (IV) – цитохромоксидаза. Обладает абсолютной специфичностью к
IV. Четвертый ферментативный комплекс (IV) – цитохромоксидаза. Обладает абсолютной специфичностью к

Участки ( пункт) сопряжения дыхательной цепи.
В дыхательной цепи выделяют
Участки ( пункт) сопряжения дыхательной цепи.
В дыхательной цепи выделяют

Окислительное фосфорилирование
Трансформация электрохимического потенциала в энергию макроэргической связи АТФ -
Окислительное фосфорилирование
Трансформация электрохимического потенциала в энергию макроэргической связи АТФ -

Система стремится к равновесию и при достижении определенного значения градиента концентраций
Система стремится к равновесию и при достижении определенного значения градиента концентраций

Коэффициент сопряжения (коэффициент окислительного фосфорилирования)
Р/О коэффициент сопряжения – количественная характеристика сопряжения
Коэффициент сопряжения (коэффициент окислительного фосфорилирования)
Р/О коэффициент сопряжения – количественная характеристика сопряжения

Дыхательный контроль
Интенсивность тканевого дыхания зависит от соотношения в клетке АТФ /
Дыхательный контроль
Интенсивность тканевого дыхания зависит от соотношения в клетке АТФ /

По механизмы действия на 3 группы:
1.Ингибиторы тканевого дыхания
2.Ингибиторы окислительного
фосфорилирования
По механизмы действия на 3 группы: 1.Ингибиторы тканевого дыхания 2.Ингибиторы окислительного фосфорилирования

Ингибиторы тканевого дыхания
Снижают создание электрохимического потенциала, прерывая поток электронов на участках
Ингибиторы тканевого дыхания
Снижают создание электрохимического потенциала, прерывая поток электронов на участках

Ингибиторы окислительного фосфорилирования
Связываются с протомерами АТФ-синтазы, запечатывая протонный канал. Прекращается синтез
Ингибиторы окислительного фосфорилирования
Связываются с протомерами АТФ-синтазы, запечатывая протонный канал. Прекращается синтез
 Ботаника. Цветок
Ботаника. Цветок Комнатные цветы
Комнатные цветы Подтип Chelicerata (хелицеровые)
Подтип Chelicerata (хелицеровые) Полужесткокрылые
Полужесткокрылые Стадии и общая схема почвообразования
Стадии и общая схема почвообразования Презентация ГМО и ГМ (ГМИ)
Презентация ГМО и ГМ (ГМИ)  Водоём и его обитатели
Водоём и его обитатели Красная книга
Красная книга Презентация на тему "Биоэкология" - скачать презентации по Биологии
Презентация на тему "Биоэкология" - скачать презентации по Биологии Химический состав клетки
Химический состав клетки Презентация на тему "Селекция растений" - скачать презентации по Биологии
Презентация на тему "Селекция растений" - скачать презентации по Биологии Размножение и развитие организмов
Размножение и развитие организмов Психофизиология эмоций. Дефиниция. Субстрат эмоций. Межполушарная асимметрия и эмоции
Психофизиология эмоций. Дефиниция. Субстрат эмоций. Межполушарная асимметрия и эмоции Бобр-строитель
Бобр-строитель Общая характеристика класса Млекопитающие
Общая характеристика класса Млекопитающие Вирусы. Бактериофаги
Вирусы. Бактериофаги Металлы в организме человека
Металлы в организме человека Клонування організмів
Клонування організмів Организм как биосистема Автор : Кудяшева Вера Григорьевна МОУ СОШ №12 Имени ОЛЕГА Кошевого Сыктывкара
Организм как биосистема Автор : Кудяшева Вера Григорьевна МОУ СОШ №12 Имени ОЛЕГА Кошевого Сыктывкара  Ваше тело просит воды
Ваше тело просит воды Презентация на тему "Строение нервной системы" - скачать презентации по Биологии
Презентация на тему "Строение нервной системы" - скачать презентации по Биологии Адаптации организмов
Адаптации организмов Водоросли, мхи, лишайники. Урок №1
Водоросли, мхи, лишайники. Урок №1 Тип инфузории. Происхождение простейших
Тип инфузории. Происхождение простейших Класс круглоротых. Миксины
Класс круглоротых. Миксины Экологические биотехнологии в Удмуртии
Экологические биотехнологии в Удмуртии Презентация на тему "Курение и здоровье будущего поколения" - скачать презентации по Биологии
Презентация на тему "Курение и здоровье будущего поколения" - скачать презентации по Биологии Основы культуры здоровья. Биологический возраст человека
Основы культуры здоровья. Биологический возраст человека