- Размножение и развитие организмов

Содержание
- 2. Цитология изучает клетки, их органоиды, строение, функционирование, процессы клеточного размножения, старения и смерти. Эмбриология изучает: гаметогенез,
- 3. История Гиппократ (460–377 г. до н. э.) - сборник, в котором содержались первые научные высказывания о
- 4. Огромное значение для развития эмбриологии сыграло появление в начале XVII в. первых микроскопов А.Левенгук, Я.Сваммердам, М.Мальпиги.
- 5. Во 2-ой половине XVIII века сформировалось учение о постепенном развитии и новообразовании, в процессе которого строение
- 6. В 1859 г. «Происхождения видов» Ч.Дарвина (1809-1882). Дарвинизм прежде всего подрывал главную опору телеологического мировоззрения, указывая
- 7. В 70–80-е годы XIX в. началось формирование аналитической и экспериментальной эмбриологии, внесшей наибольший вклад в изучение
- 8. Г. Шпеман (1869–1941) и его школа в 1924 г. открыли явление эмбриональной индукции. Указал, что дальнейшее
- 9. П.П.Иванов (1878–1942) – автор теории о ларвальном и постларвальном отделах тела первичноротых, устанавливающую общую закономерность сегментации
- 10. Как отражение этих новых представлений появились понятия (введенные в 1925 г. Н.В.Тимофеевым-Ресовским) «пенетрантность» (процент особей в
- 11. Размножение – свойство живых организмов воспроизводить себе подобных, это процесс смены поколений. Размножение происходит на разных
- 12. 1) в воспроизведении участвует одна родительская особь; 2) источник генетической информации – соматические клетки; 3) генотипы
- 13. Группой клеток (полицитогенное) Одной клеткой (моноцитогенное) Бесполое Простое деление надвое Множественное деление (шизогония) Неравномерное деление или
- 14. Половое Конъюгация - форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися
- 15. МИТОЗ В профазе увеличивается объем ядра, хроматин спирализуется, формируются хромосомы. К концу каждая хромосома состоит из
- 16. В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки, поэтому их подсчет и
- 17. НЕТИПИЧНЫЕ ФОРМЫ МИТОЗА 1. Амитоз — это прямое деление ядра. При этом сохраняется морфология ядра, видны
- 18. МЕЙОЗ В метафазе I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их
- 19. В профазе II происходят те же процессы, что и в профазе митоза. В метафазе II хромосомы
- 21. Скачать презентацию
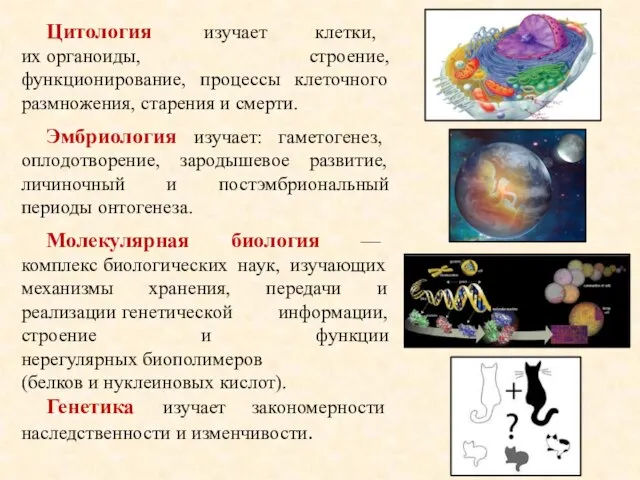
Цитология изучает клетки, их органоиды, строение, функционирование, процессы клеточного размножения, старения и
Цитология изучает клетки, их органоиды, строение, функционирование, процессы клеточного размножения, старения и

История
Гиппократ (460–377 г. до н. э.) - сборник, в котором
История
Гиппократ (460–377 г. до н. э.) - сборник, в котором

Огромное значение для развития эмбриологии сыграло появление в начале XVII в.
Огромное значение для развития эмбриологии сыграло появление в начале XVII в.

Во 2-ой половине XVIII века сформировалось учение о постепенном развитии и новообразовании,
Во 2-ой половине XVIII века сформировалось учение о постепенном развитии и новообразовании,

В 1859 г. «Происхождения видов» Ч.Дарвина (1809-1882). Дарвинизм прежде всего подрывал
В 1859 г. «Происхождения видов» Ч.Дарвина (1809-1882). Дарвинизм прежде всего подрывал

В 70–80-е годы XIX в. началось формирование аналитической и экспериментальной эмбриологии,
В 70–80-е годы XIX в. началось формирование аналитической и экспериментальной эмбриологии,

Г. Шпеман (1869–1941) и его школа в 1924 г. открыли явление
Г. Шпеман (1869–1941) и его школа в 1924 г. открыли явление

П.П.Иванов (1878–1942) – автор теории о ларвальном и постларвальном отделах тела
П.П.Иванов (1878–1942) – автор теории о ларвальном и постларвальном отделах тела

Как отражение этих новых представлений появились понятия (введенные в 1925 г.
Как отражение этих новых представлений появились понятия (введенные в 1925 г.

Размножение – свойство живых организмов
воспроизводить себе подобных, это процесс смены поколений.
Размножение – свойство живых организмов
воспроизводить себе подобных, это процесс смены поколений.

1) в воспроизведении участвует одна родительская особь;
2) источник генетической информации –
1) в воспроизведении участвует одна родительская особь;
2) источник генетической информации –
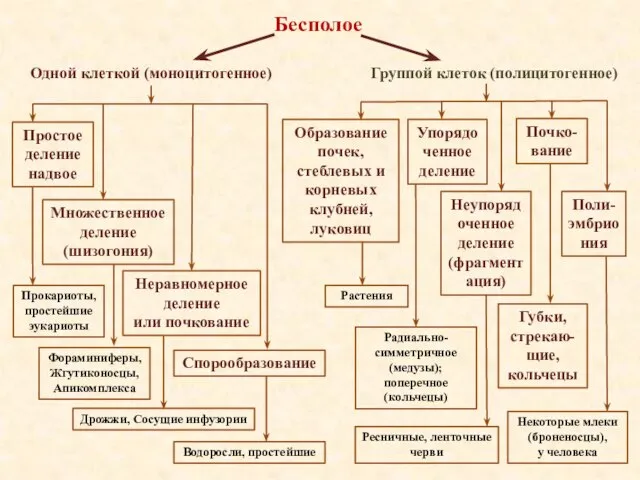
Группой клеток (полицитогенное)
Одной клеткой (моноцитогенное)
Бесполое
Простое деление
надвое
Множественное деление
(шизогония)
Неравномерное деление
или почкование
Спорообразование
Прокариоты, простейшие эукариоты
Фораминиферы,
Жгутиконосцы,
Апикомплекса
Дрожжи,
Группой клеток (полицитогенное)
Одной клеткой (моноцитогенное)
Бесполое
Простое деление
надвое
Множественное деление
(шизогония)
Неравномерное деление
или почкование
Спорообразование
Прокариоты, простейшие эукариоты
Фораминиферы,
Жгутиконосцы,
Апикомплекса
Дрожжи,
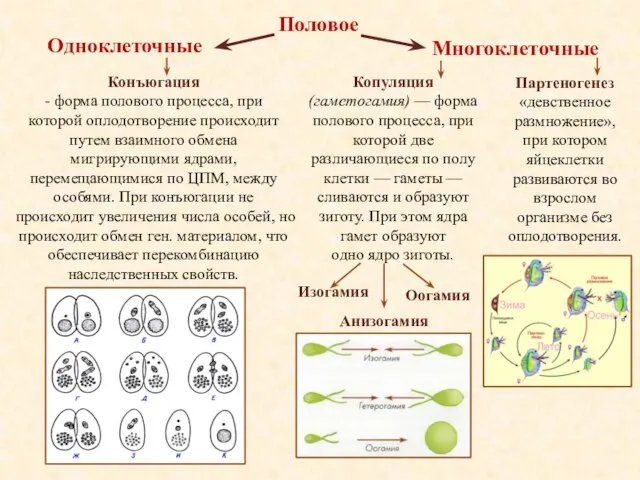
Половое
Конъюгация
- форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами,
Половое
Конъюгация
- форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами,
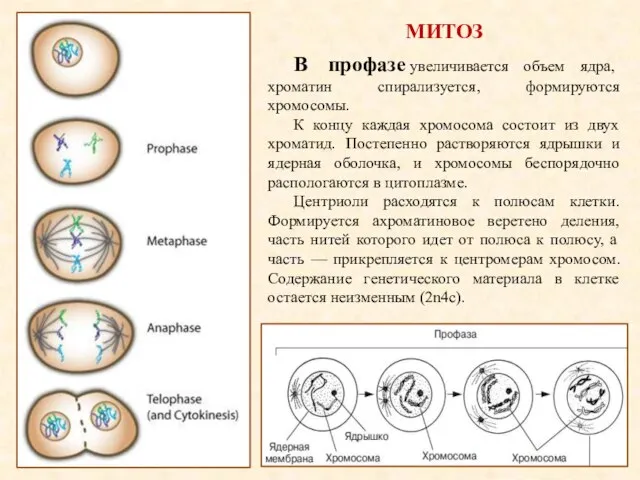
МИТОЗ
В профазе увеличивается объем ядра, хроматин спирализуется, формируются хромосомы.
К концу каждая
МИТОЗ
В профазе увеличивается объем ядра, хроматин спирализуется, формируются хромосомы.
К концу каждая
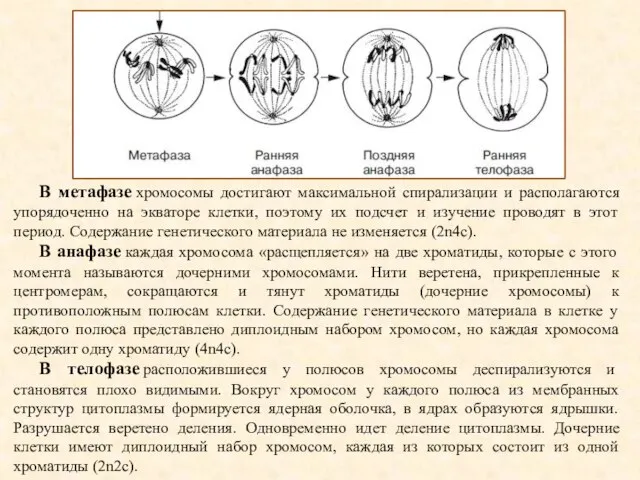
В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки,
В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки,

НЕТИПИЧНЫЕ ФОРМЫ МИТОЗА
1. Амитоз — это прямое деление ядра. При этом
НЕТИПИЧНЫЕ ФОРМЫ МИТОЗА
1. Амитоз — это прямое деление ядра. При этом
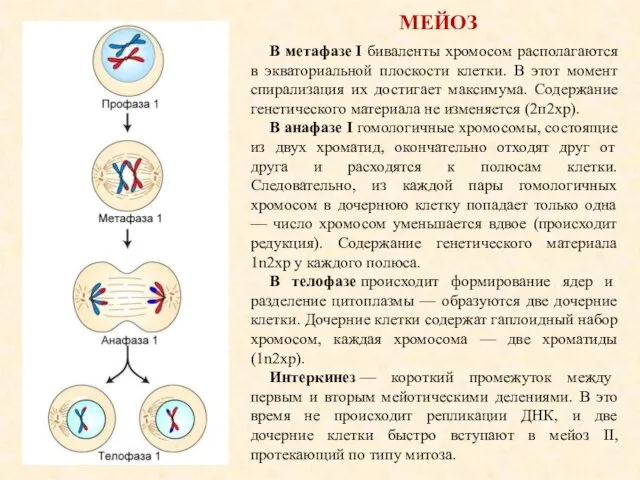
МЕЙОЗ
В метафазе I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот
МЕЙОЗ
В метафазе I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот
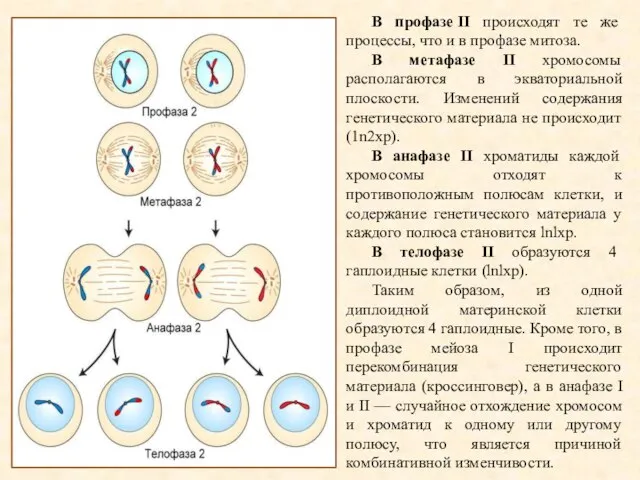
В профазе II происходят те же процессы, что и в профазе митоза.
В профазе II происходят те же процессы, что и в профазе митоза.
 Этапы развития эмбриона, онтогенез и филогенез, адаптация и симбиоз
Этапы развития эмбриона, онтогенез и филогенез, адаптация и симбиоз Медузи. Ознайомитися з будовою і різноманітністю медуз
Медузи. Ознайомитися з будовою і різноманітністю медуз Человеческие расы Презентация Расуловой Марьям 7 Б
Человеческие расы Презентация Расуловой Марьям 7 Б  Презентация на тему "Побег. Почка" - скачать презентации по Биологии
Презентация на тему "Побег. Почка" - скачать презентации по Биологии Mother s Love
Mother s Love Птицы перелётные
Птицы перелётные Презентация на тему Вирусы-неклеточная форма жизни
Презентация на тему Вирусы-неклеточная форма жизни  Структура и функции кожи человека
Структура и функции кожи человека Половое и бесполое размножение
Половое и бесполое размножение Органические вещества, входящие в состав клетки
Органические вещества, входящие в состав клетки Кордицепс однобокий. Энтомопатогенные грибы
Кордицепс однобокий. Энтомопатогенные грибы РАСТЕНИЯ СЕМЕЙСТВА ПАСЛЁНОВЫЕ, ИСПОЛЬЗУЕМЫЕ ЧЕЛОВЕКОМ Выполнила: ученица 6 класса МОУ «Долголесковская основная общеобразовател
РАСТЕНИЯ СЕМЕЙСТВА ПАСЛЁНОВЫЕ, ИСПОЛЬЗУЕМЫЕ ЧЕЛОВЕКОМ Выполнила: ученица 6 класса МОУ «Долголесковская основная общеобразовател Строение плесневого гриба-мукора
Строение плесневого гриба-мукора Ковалентная химическая связь
Ковалентная химическая связь Физиология мышц
Физиология мышц Водоросли- красные, бурые, зеленые. (Занятие 3.2)
Водоросли- красные, бурые, зеленые. (Занятие 3.2) Строение эукариотической клетки
Строение эукариотической клетки Живые системы. Лекция 6
Живые системы. Лекция 6 Мир комнатных растений
Мир комнатных растений Задачи на биосинтез белка ЕГЭ
Задачи на биосинтез белка ЕГЭ Кошки
Кошки Отряд верблюдки (raphidioptera)
Отряд верблюдки (raphidioptera) Типы соединения костей
Типы соединения костей Презентація з Медицини на тему: “кровотеча та надання першої допомоги при кровотечі”
Презентація з Медицини на тему: “кровотеча та надання першої допомоги при кровотечі”  Селекция животных
Селекция животных  Сердечно-сосудистая система человека
Сердечно-сосудистая система человека Появление и развитие жизни
Появление и развитие жизни Практическая работа №1. Анализ почвы пришкольного участка
Практическая работа №1. Анализ почвы пришкольного участка