- Эволюция филогенетических групп

Содержание
- 2. ТЕНДЕНЦИИ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ "Закон постепенного образования всего сущего" (Рулье, 1852). "Закон увеличения разнообразия организмов" (Рулье, 1852).
- 3. Правила эволюции 1. Необратимость эволюции (Л.Долло, 1893). 2. Правило прогрессирующей специализации (Ш.Депре, 1876) теломорфоз, гиперморфоз, катаморфоз,
- 4. СКОРОСТЬ ЭВОЛЮЦИИ Горотелия — средний, обычный темп эволюционного процесса (хищные млекопитающие и брюхоногие моллюски, амфибии в
- 5. НАПРАВЛЕНИЯ ЭВОЛЮЦИИ Аллогенез – в пределах одной адаптивной зоны (идиоадаптация), а- алломорфозы – изменение без сужения
- 7. ФОРМЫ ФИЛОГЕНЕЗА 1. Филетическая эволюция или ортогенез. 2. Дивергенция. 3. Конвергенция. 4. Параллелизм, а- синхронный, б-
- 8. Дивергенция
- 9. Конвергенция
- 10. Филетическая эволюция … конечности Ортогенез - прямое развитие (без дивергенции), но ортогенез признаков отнюдь не подразумевает
- 14. Синхронный параллелизм прогрессивное (компенсаторное) развитие 3-го пальца и редукция остальных редукция боковых пальцев почти без компенсации
- 15. Eohippus Merychippus Equus Macrauchenia Diadiaphorus Thoatherium
- 16. Eohippus Macrauchenia
- 17. Асинхронный параллелизм (саблезубость у кошачьих)
- 18. Асинхронный параллелизм (саблезубость у кошачьих)
- 19. Асинхронный параллелизм (саблезубость у кошачьих) Pseudoaelurus миоцен Holophoneus, олигоцен Dinictis, олигоцен Smilodon, плейстоцен
- 20. Panderichthys spp. 378 млн.лет Tulerpeton curtum 363 млн.лет Ichthyostega sp. 363 млн.лет 370 млн.лет Tiktaalik roseae
- 22. Поздняя юра мел
- 23. Нижний мел Caudipteryx zoui Coelurosaurus Microraptor gui
- 24. ранний мел поздний триас
- 25. Параллельное появление перьев и полета
- 26. Walker, C. A., Buffetaut, E., and Dyke, G. J., 2007, Large euenantiornithine birds from the Cretaceous
- 27. 1. Вытеснение менее приспособленной группы более приспособленной. 2. Дивергенция каждого таксона в пределах своей адаптивной зоны
- 28. Пример возникновения похожих наборов "жизненных форм" на разобщенных континентах При параллельном развитии сходство объясняется частью общностью
- 29. Представители планцентарных (слева) и сумчатых (справа) млекопитающих
- 30. Ambystoma macrodactylum Rana sylvatica Rana luteiventris Rana pipiens Pseudacris maculata Bufo hemiophrys Bufo boreas Spea bombifrons
- 31. Параллелизмы и их эволюционное значение Л.П.Татаринов,1987 1. Генотипические (в основе лежат гомологические мутации). 2. Эпигенетические а)
- 32. Причины параллелизмов Принципиальное сходство в строении основных типов сообществ, или биоценозов; сходство наборов экологических ниш в
- 33. Daniel M. Weinreich, Nigel F. Delaney, Mark A. DePristo, Daniel L. Hartl. Darwinian Evolution Can Follow
- 35. Система, состоящая из взаимосвязанных элементов (блоков), может существовать в различных устойчивых состояниях, переходя из одного в
- 36. Эволюционный прогресс и регресс Признаки биологического прогресса: 1- увеличение численности, 2- увеличение ареала, 3- расширение экологической
- 37. Причины эволюционного прогресса Общая тенденция - усложнение организации. В открытых системах с постоянным поступлением вещества и
- 38. … а также «преадаптации». Только часть регуляторных связей важна для адаптации в конкретный момент. Все остальные
- 39. - прогресс тормозится необходимостью сохранения целостности и нормального функционирования старой, сложившейся системы (А.П.Расницын). Принцип «ключевого ароморфоза»
- 40. Активизация функции легочного дыхания как ключевой ароморфоз синдрома дефинитивных признаков рептильного уровня организации и некоторых признаков
- 41. Наличие оперения как ключевой ароморфоз синдрома признаков уровня организации птиц. Составил А. С. Раутиан
- 42. Биохимия: (ранние этапы) появлялись и совершенствовались новые ферментые системы, новые метаболические пути. Нарастала роль регуляторных систем
- 44. Скачать презентацию

ТЕНДЕНЦИИ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
"Закон постепенного образования всего сущего" (Рулье, 1852).
"Закон
ТЕНДЕНЦИИ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
"Закон постепенного образования всего сущего" (Рулье, 1852).
"Закон

Правила эволюции
1. Необратимость эволюции (Л.Долло, 1893).
2. Правило прогрессирующей специализации (Ш.Депре,
Правила эволюции
1. Необратимость эволюции (Л.Долло, 1893).
2. Правило прогрессирующей специализации (Ш.Депре,

СКОРОСТЬ ЭВОЛЮЦИИ
Горотелия — средний, обычный темп эволюционного процесса (хищные млекопитающие
СКОРОСТЬ ЭВОЛЮЦИИ
Горотелия — средний, обычный темп эволюционного процесса (хищные млекопитающие

НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
Аллогенез – в пределах одной адаптивной зоны (идиоадаптация), а- алломорфозы
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
Аллогенез – в пределах одной адаптивной зоны (идиоадаптация), а- алломорфозы


ФОРМЫ ФИЛОГЕНЕЗА
1. Филетическая эволюция или ортогенез.
2. Дивергенция.
3. Конвергенция.
4. Параллелизм,
ФОРМЫ ФИЛОГЕНЕЗА
1. Филетическая эволюция или ортогенез.
2. Дивергенция.
3. Конвергенция.
4. Параллелизм,

Дивергенция
Дивергенция
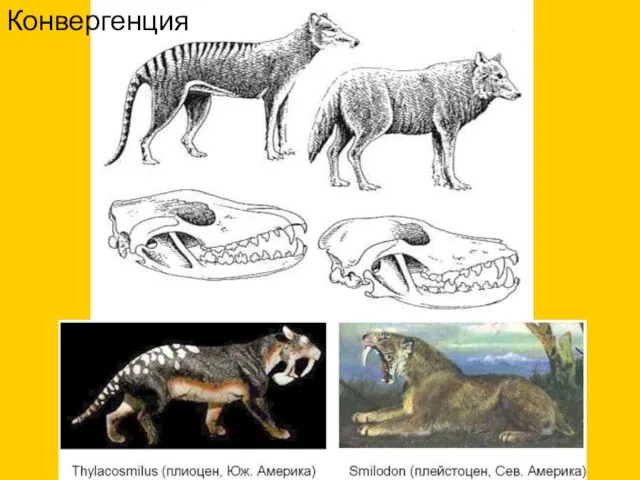
Конвергенция
Конвергенция
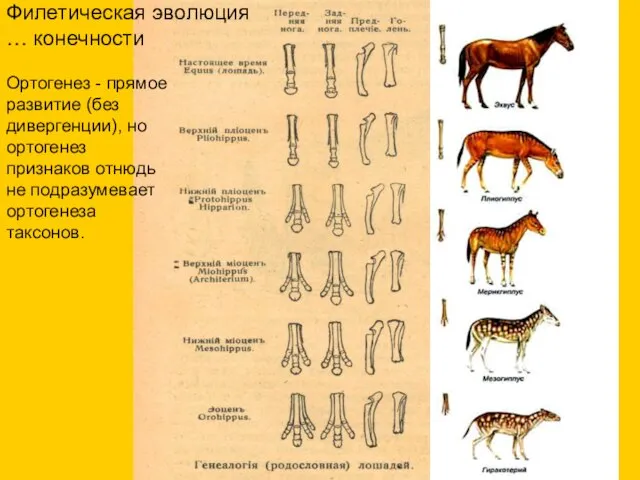
Филетическая эволюция
… конечности
Ортогенез - прямое развитие (без дивергенции), но ортогенез
Филетическая эволюция
… конечности
Ортогенез - прямое развитие (без дивергенции), но ортогенез
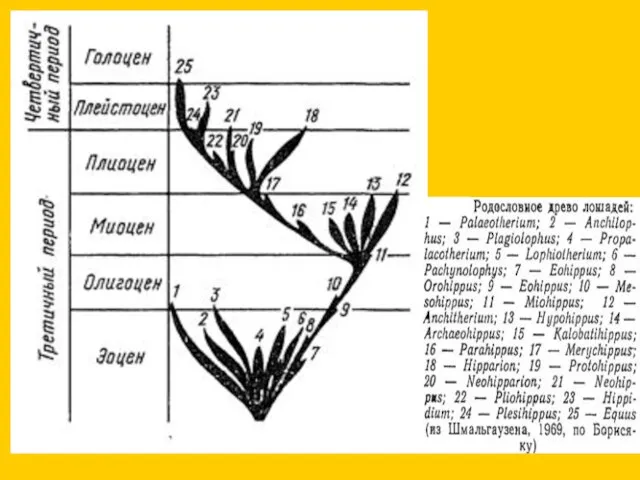

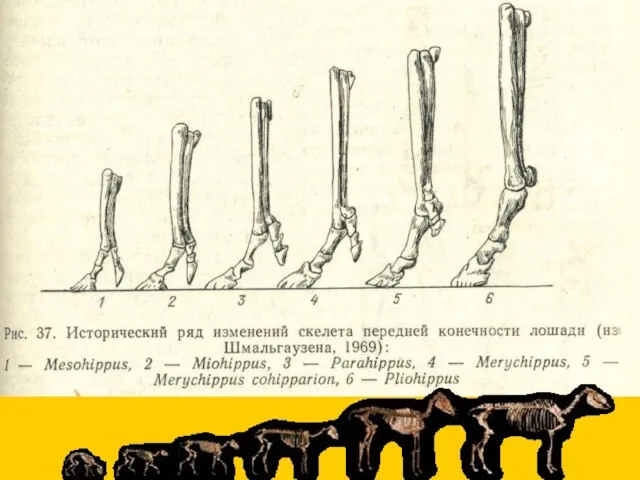
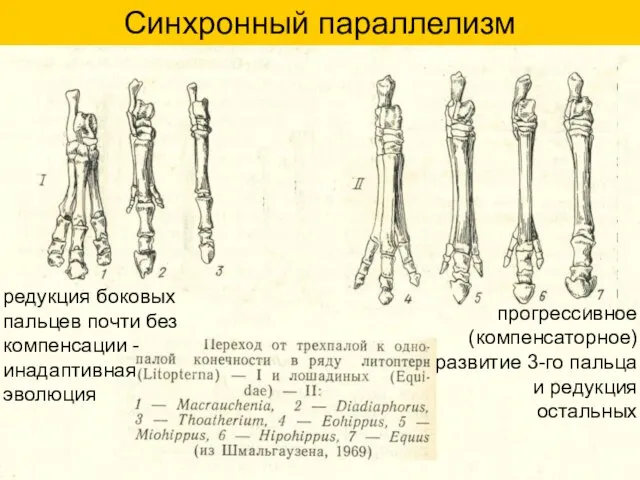
Синхронный параллелизм
прогрессивное (компенсаторное) развитие 3-го пальца и редукция остальных
редукция боковых пальцев
Синхронный параллелизм
прогрессивное (компенсаторное) развитие 3-го пальца и редукция остальных
редукция боковых пальцев
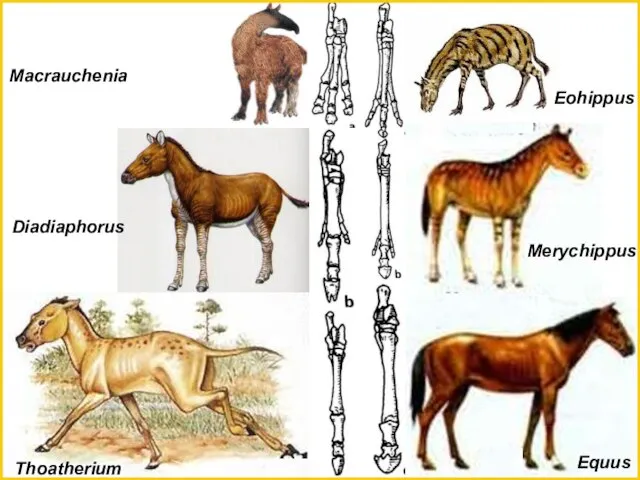
Eohippus
Merychippus
Equus
Macrauchenia
Diadiaphorus
Thoatherium
Merychippus
Equus
Macrauchenia
Diadiaphorus
Thoatherium
Eohippus
Macrauchenia
Eohippus
Macrauchenia

Асинхронный параллелизм
(саблезубость у кошачьих)
Асинхронный параллелизм
(саблезубость у кошачьих)
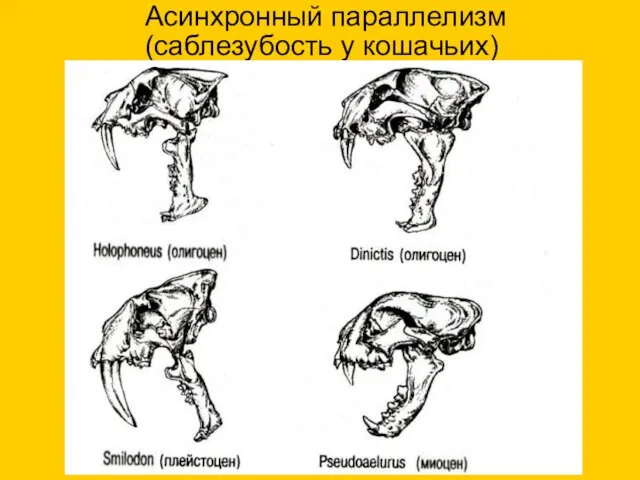
Асинхронный параллелизм
(саблезубость у кошачьих)
Асинхронный параллелизм
(саблезубость у кошачьих)

Асинхронный параллелизм
(саблезубость у кошачьих)
Pseudoaelurus миоцен
Holophoneus, олигоцен
Dinictis, олигоцен
Smilodon, плейстоцен
Асинхронный параллелизм
(саблезубость у кошачьих)
Pseudoaelurus миоцен
Holophoneus, олигоцен
Dinictis, олигоцен
Smilodon, плейстоцен
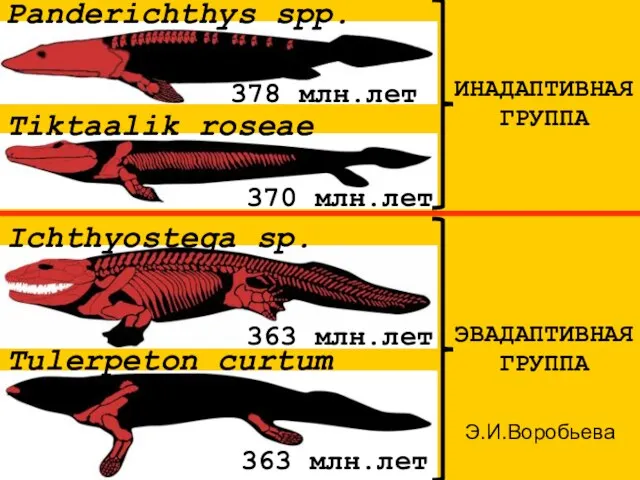
Panderichthys spp.
378 млн.лет
Tulerpeton curtum
363 млн.лет
Ichthyostega sp.
363 млн.лет
370 млн.лет
Tiktaalik roseae
ИНАДАПТИВНАЯ
ГРУППА
ЭВАДАПТИВНАЯ
ГРУППА
Э.И.Воробьева
Panderichthys spp.
378 млн.лет
Tulerpeton curtum
363 млн.лет
Ichthyostega sp.
363 млн.лет
370 млн.лет
Tiktaalik roseae
ИНАДАПТИВНАЯ
ГРУППА
ЭВАДАПТИВНАЯ
ГРУППА
Э.И.Воробьева

Поздняя юра
мел
Поздняя юра
мел
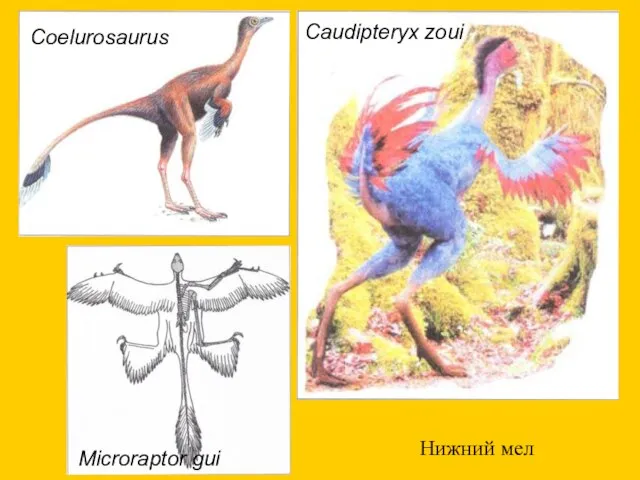
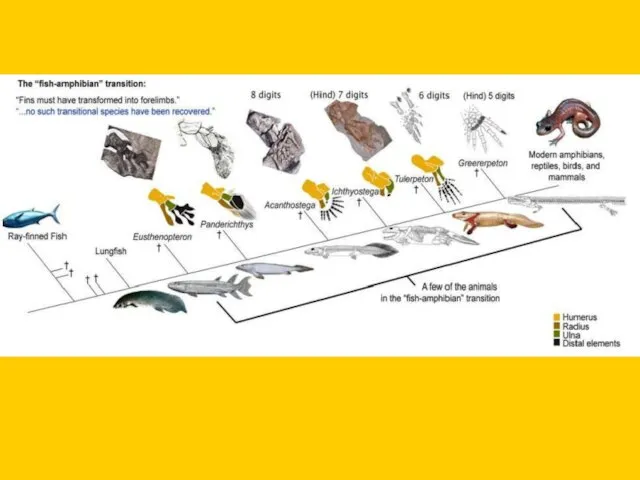
Нижний мел
Caudipteryx zoui
Coelurosaurus
Microraptor gui
Нижний мел
Caudipteryx zoui
Coelurosaurus
Microraptor gui
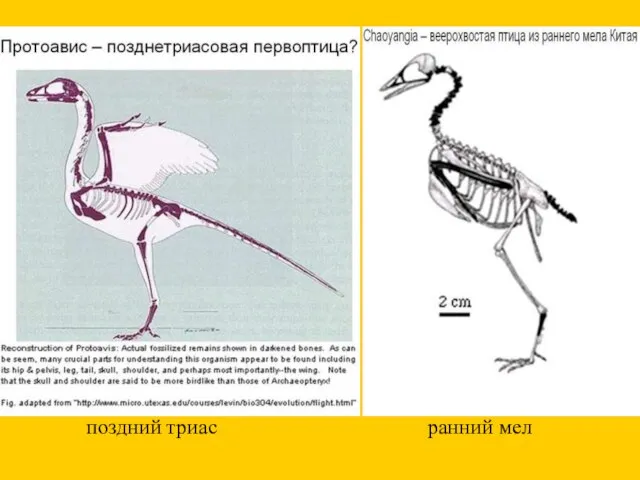
ранний мел
поздний триас
ранний мел
поздний триас
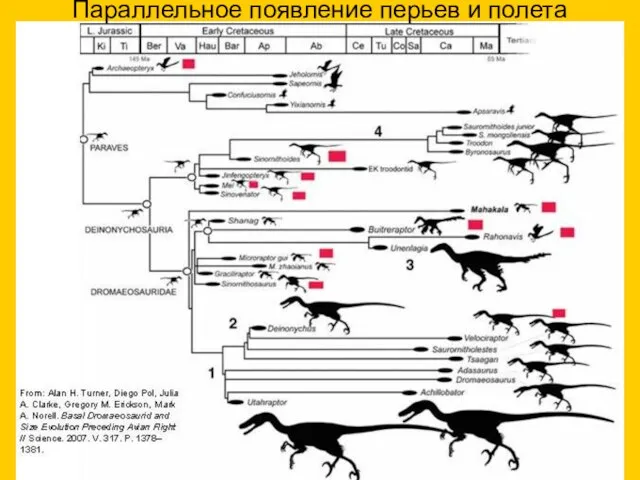
Параллельное появление перьев и полета
Параллельное появление перьев и полета
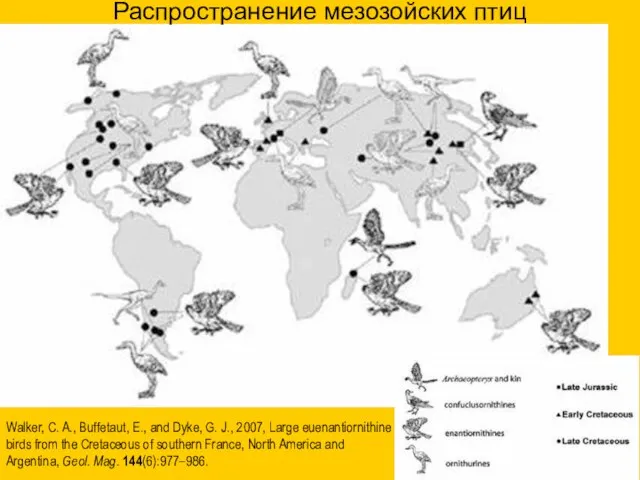
Walker, C. A., Buffetaut, E., and Dyke, G. J., 2007, Large
Walker, C. A., Buffetaut, E., and Dyke, G. J., 2007, Large

1. Вытеснение менее приспособленной группы более приспособленной.
2. Дивергенция каждого таксона
1. Вытеснение менее приспособленной группы более приспособленной.
2. Дивергенция каждого таксона

Пример возникновения похожих наборов "жизненных форм" на разобщенных континентах
При параллельном развитии
Пример возникновения похожих наборов "жизненных форм" на разобщенных континентах
При параллельном развитии

Представители планцентарных (слева) и сумчатых (справа) млекопитающих
Представители планцентарных (слева) и сумчатых (справа) млекопитающих

Ambystoma macrodactylum
Rana sylvatica
Rana luteiventris
Rana pipiens
Pseudacris maculata
Bufo hemiophrys
Bufo boreas
Spea bombifrons
Salamandrella keyserlingii
Rana
Ambystoma macrodactylum
Rana sylvatica
Rana luteiventris
Rana pipiens
Pseudacris maculata
Bufo hemiophrys
Bufo boreas
Spea bombifrons
Salamandrella keyserlingii
Rana

Параллелизмы и их эволюционное значение
Л.П.Татаринов,1987
1. Генотипические (в основе лежат
Параллелизмы и их эволюционное значение
Л.П.Татаринов,1987
1. Генотипические (в основе лежат

Причины параллелизмов
Принципиальное сходство в строении основных типов сообществ, или биоценозов;
Причины параллелизмов
Принципиальное сходство в строении основных типов сообществ, или биоценозов;

Daniel M. Weinreich, Nigel F. Delaney, Mark A. DePristo, Daniel L.
Daniel M. Weinreich, Nigel F. Delaney, Mark A. DePristo, Daniel L.


Система, состоящая из взаимосвязанных элементов (блоков), может существовать в различных
Система, состоящая из взаимосвязанных элементов (блоков), может существовать в различных

Эволюционный прогресс и регресс
Признаки биологического прогресса:
1- увеличение численности,
2- увеличение
Эволюционный прогресс и регресс
Признаки биологического прогресса:
1- увеличение численности,
2- увеличение

Причины эволюционного прогресса
Общая тенденция - усложнение организации.
В открытых системах с постоянным
Причины эволюционного прогресса
Общая тенденция - усложнение организации.
В открытых системах с постоянным

… а также «преадаптации». Только часть регуляторных связей важна для адаптации
… а также «преадаптации». Только часть регуляторных связей важна для адаптации

- прогресс тормозится необходимостью сохранения целостности и нормального функционирования старой,
- прогресс тормозится необходимостью сохранения целостности и нормального функционирования старой,
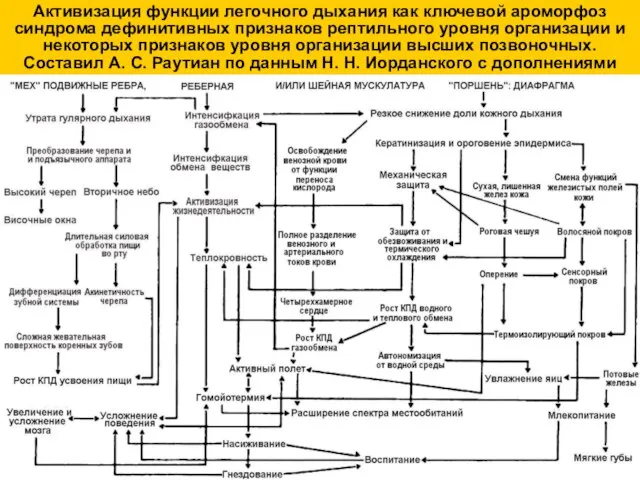
Активизация функции легочного дыхания как ключевой ароморфоз синдрома дефинитивных признаков рептильного
Активизация функции легочного дыхания как ключевой ароморфоз синдрома дефинитивных признаков рептильного

Наличие оперения как ключевой ароморфоз синдрома признаков уровня организации птиц. Составил
Наличие оперения как ключевой ароморфоз синдрома признаков уровня организации птиц. Составил

Биохимия: (ранние этапы) появлялись и совершенствовались новые ферментые системы, новые
Биохимия: (ранние этапы) появлялись и совершенствовались новые ферментые системы, новые
 Как называются эти пары хромосом?
Как называются эти пары хромосом? Kharari_Sapiens
Kharari_Sapiens Австралопитеки
Австралопитеки Презентация на тему Организм и его свойства
Презентация на тему Организм и его свойства  Жизненный цикл клетки
Жизненный цикл клетки Нервная ткань
Нервная ткань Фотосинтез
Фотосинтез Клетка. Учение о тканях. Виды тканей
Клетка. Учение о тканях. Виды тканей Анатомия, физиология и гигиена человека Анатомия - наука о форме и строении живых организмов
Анатомия, физиология и гигиена человека Анатомия - наука о форме и строении живых организмов Тип Кольчатые черви. Класс Малощетинковые
Тип Кольчатые черви. Класс Малощетинковые Питание растений и грибов
Питание растений и грибов Обобщающее повторение
Обобщающее повторение Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. (Часть 3)
Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. (Часть 3) Процессы торможения в ЦНС
Процессы торможения в ЦНС Функциональная анатомия ствола головного мозга
Функциональная анатомия ствола головного мозга Подцарство простейшие. Амебовые, жгутиконосцы
Подцарство простейшие. Амебовые, жгутиконосцы Бионика и биоинженерия
Бионика и биоинженерия Презентация на тему Биология – наука о живой природе
Презентация на тему Биология – наука о живой природе Роль гормонів у фізичному, психічному і статевому розвитку
Роль гормонів у фізичному, психічному і статевому розвитку Гаметогенез, оплодотворение
Гаметогенез, оплодотворение Презентация по биологии Почвенная среда обитания
Презентация по биологии Почвенная среда обитания  Грибы-паразиты. Головня
Грибы-паразиты. Головня Фотосинтез
Фотосинтез Дослідження росту вегетативних органів рослин
Дослідження росту вегетативних органів рослин Міні – проект на тему: Опале листя. Це користь чи шкода?
Міні – проект на тему: Опале листя. Це користь чи шкода? Овогенез. Сперматогенез
Овогенез. Сперматогенез Витамины
Витамины Презентация на тему "Современное представление о механизмах и закономерностях эволюции" - скачать презентации по Биологии
Презентация на тему "Современное представление о механизмах и закономерностях эволюции" - скачать презентации по Биологии