- Физиология нервов и нервно-мышечных синапсов

Содержание
- 2. АКСОННЫЙ ТРАНСПОРТ НЕРВЫ ЧЕЛОВЕКА «ТЕКУТ», КАК РЕКИ
- 3. В НЕРВАХ «ТЕЧЕТ» АКСОПЛАЗМА АКСОПЛАЗМА ТРАНСПОРТИРУЕТ : БЕЛКИ, ПЕПТИДЫ, ГЛЮКОЗУ, АТФ ВЕЗИКУЛЫ, МЕДИАТОРНЫЕ СИСТЕМЫ, ФАКТОРЫ РОСТА
- 4. ВИДЫ ТРАНСПОРТА АНТЕРОГРАДНЫЙ (ПРЯМОЙ): БЫСТРЫЙ – 400 мм/день МЕДЛЕННЫЙ – 1-4 мм/день РЕТРОГРАДНЫЙ (ОБРАТНЫЙ)- 200-300 мм/день
- 6. Антероградный транспорт
- 7. Транспорт в аксоне осуществляют: Микротрубочки Фибриллярные белки: семейства кинезинов и динеинов АТФ
- 8. Работа моторных белков Головки кинезина и динеина
- 9. Механизм транспорта
- 10. Ретроградный транспорт переносит: Вирусы, бактерии, токсины, Лекарства от терминалей аксона к телу нейрона
- 11. Трафик лекарств
- 12. Ретроградный транспорт используется в научных целях Для топической диагностики (локализации функций)
- 13. Введение радиометки Синтез радиоактивных белков Радиоактивные белки двигаются по аксону
- 14. Нервно-мышечная трансмиссия Передача возбуждения с нерва на мышцу
- 15. ПЕРЕДАЧА ВОЗБУЖДЕНИЯ С НЕРВА НА МЫШЦУ ПРОИСХОДИТ С ПОМОЩЬЮ СИНАПСА Ч. Шеррингтон в 1897 году ввел
- 16. Физиологические свойства химических синапсов Одностороннее проведение ПД. Обязательное участие медиаторов. Количество медиатора пропорционально частоте ПД. Передача
- 17. Дизайн синапса 1 - Шванновские клетки 2 - ПД моторного нерва 3 - Вход ионов Са
- 18. Внутриклеточная регистрация МП, ПКП и ПД мышечного волокна
- 19. Функция синапса вне возбуждения В покое существует связь между пре- и постсинаптическими мембранами – в минимальных
- 20. МП, МПКП и ПД концевой пластинки мышечного волокна
- 21. Этапы нервно-мышечной трансмиссии. 1. Синапс в покое.
- 22. 2. Работа синапса при возбуждении: ПД моторного нерва, экзоцитоз везикул с ацетилхолином в синаптическую щель
- 23. 3. Диффузия ацетилхолина в синаптической щели,связь ацетилхолина с холинорецептором, возникновение ПКП постсинаптической мембраны
- 24. 4. Разрушение ацетилхолина (холинэстераза), захват ацетила и холина пресинаптической мембраной
- 25. Ультраструктура нервно-мышечного синапса
- 26. Действие миорелаксантов на синапс
- 27. Ботокс – локальный миорелаксант Большой Мол. вес (900 кДа), поэтому не распространяется в окружающие ткани. Блокирует
- 28. Действие ботокса на синапс – 1 этап Связывание Молекула токсина тяжелой цепью связывается с окончанием двигательного
- 29. 2 этап Интернализа-ция Молекула поглощается нервным окончанием и расщепляется на лёгкую и тяжёлые цепи
- 30. 3 этап Блокирование Лёгкая цепь расщепляет транспортные белки. Ацетилхолин не выделяется. Мышца расслабляется
- 31. 4 этап Спраутинг Развиваются дополнительные отростки аксона
- 32. 5 этап Восстановление Формируются новые нервно-мышечные синапсы. Восстанавливаются транспортные белки. Мышца вновь способна к сокращению
- 33. Рецепторы – это чувствительные органы, способные реагировать на различные виды энергии (электрической, механической , химической, тепловой,
- 34. 3 функциональные роли рецепторов: Сенсор (датчик) энергии раздражителя – воспринимает, чувствует любой вид энергии стимула. Трансдуктор
- 35. Разные стимулы вызывают в рецепторах трансдукцию
- 36. Схема трансдукции и трансмиссии
- 37. 2 вида рецепторов Первичные – это нервные клетки с периферически-чувствительным и центрально-проводящим отростками. Вторичные – это
- 38. Сенсорная трансдукция, как правило, приводит к возникновению рецепторного потенциала -РП. Это значит, что стимул воздействует на
- 39. Схема возникновения РП
- 40. Схема возникновения ГП
- 41. Что объединяет мышцу, нерв и рецептор? Наличие возбудимой мембраны
- 42. Физиология нервных волокон (аксонов) Нервы являются отростками нервных клеток. Нервы выполняют роль проводников. Нервы обладают кабельными
- 43. Схема нервного ствола
- 44. ФУНКЦИИ НЕРВОВ Информационная - передача ПД от тела нейрона на периферию и обратно. Трофическая – трафик
- 45. Схема эксперимента Эрлангера и Гассера
- 46. Эрлангер и Гассер показали, что ПД в нервных стволах проводится с разной скоростью
- 47. Классификация нервных волокон по Эрлангеру и Гассеру А – волокна делятся на 4 подгруппы, все покрыты
- 48. Нервные волокна разного диаметра Аά Аβ АΔ С
- 49. Классификация Ллойд - Ханта
- 50. Законы проведения ПД по нерву ПД по нерву распространяется в обе стороны. ПД распространяется в обе
- 51. Продолжение ПД распространяется изолированно. Нерв практически не утомляем. В разных по диаметру нервных волокнах ПД распространяется
- 52. Схема миелинизированного волокна
- 53. Проведение ПД происходит: По миелиновому волокну – прыжками, сальтаторно. По безмиелиновому волокну – непрерывно. Механизм передачи
- 54. МП нервных волокон в покое
- 55. Передача по безмиелиновому волокну Местный (локальный) непрерывный ток от + к – деполяризует соседний участок, причем
- 56. Этапы проведение возбуждения по безмякотному волокну
- 57. Передача по миелиновому волокну ПД распространяется прыжками (сальто), т.к.миелин – хороший изолятор, а в перехватах Ранвье
- 58. Механизм возникновения пассивных токов
- 59. Механизм сальтаторного проведения ПД
- 61. Скачать презентацию

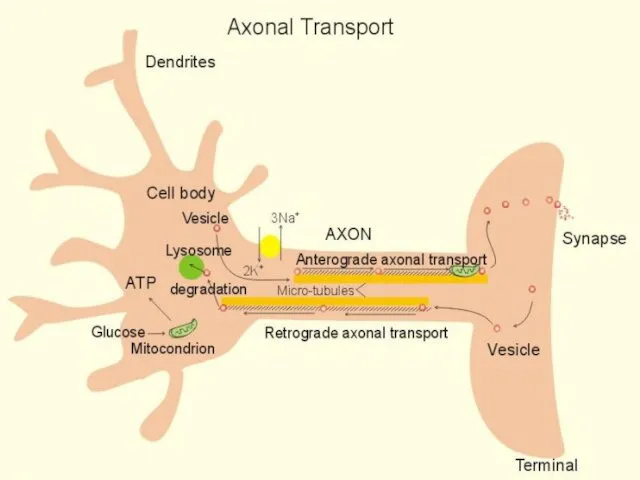
АКСОННЫЙ ТРАНСПОРТ
НЕРВЫ ЧЕЛОВЕКА «ТЕКУТ», КАК РЕКИ
АКСОННЫЙ ТРАНСПОРТ
НЕРВЫ ЧЕЛОВЕКА «ТЕКУТ», КАК РЕКИ

В НЕРВАХ «ТЕЧЕТ» АКСОПЛАЗМА
АКСОПЛАЗМА ТРАНСПОРТИРУЕТ :
БЕЛКИ, ПЕПТИДЫ, ГЛЮКОЗУ, АТФ
ВЕЗИКУЛЫ,
В НЕРВАХ «ТЕЧЕТ» АКСОПЛАЗМА
АКСОПЛАЗМА ТРАНСПОРТИРУЕТ :
БЕЛКИ, ПЕПТИДЫ, ГЛЮКОЗУ, АТФ
ВЕЗИКУЛЫ,

ВИДЫ ТРАНСПОРТА
АНТЕРОГРАДНЫЙ (ПРЯМОЙ):
БЫСТРЫЙ – 400 мм/день
МЕДЛЕННЫЙ – 1-4 мм/день
РЕТРОГРАДНЫЙ
ВИДЫ ТРАНСПОРТА
АНТЕРОГРАДНЫЙ (ПРЯМОЙ):
БЫСТРЫЙ – 400 мм/день
МЕДЛЕННЫЙ – 1-4 мм/день
РЕТРОГРАДНЫЙ
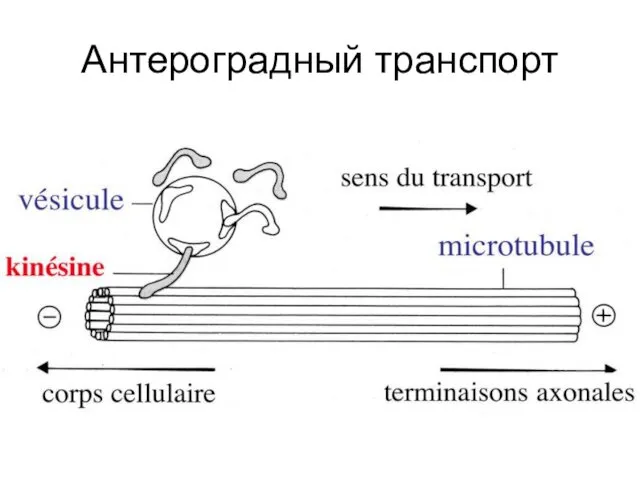
Антероградный транспорт
Антероградный транспорт
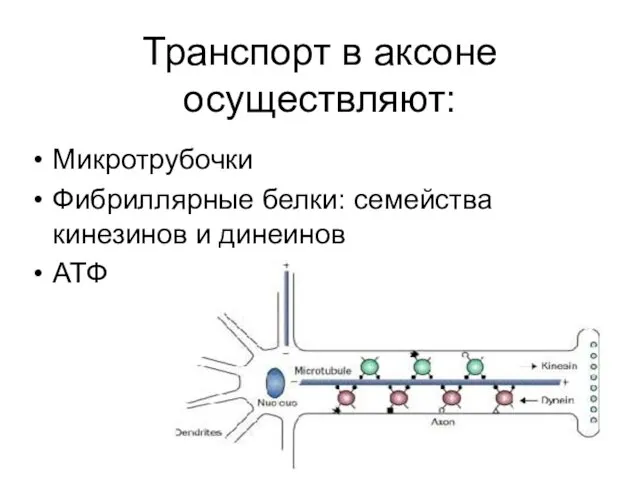
Транспорт в аксоне осуществляют:
Микротрубочки
Фибриллярные белки: семейства кинезинов и динеинов
АТФ
Транспорт в аксоне осуществляют:
Микротрубочки
Фибриллярные белки: семейства кинезинов и динеинов
АТФ
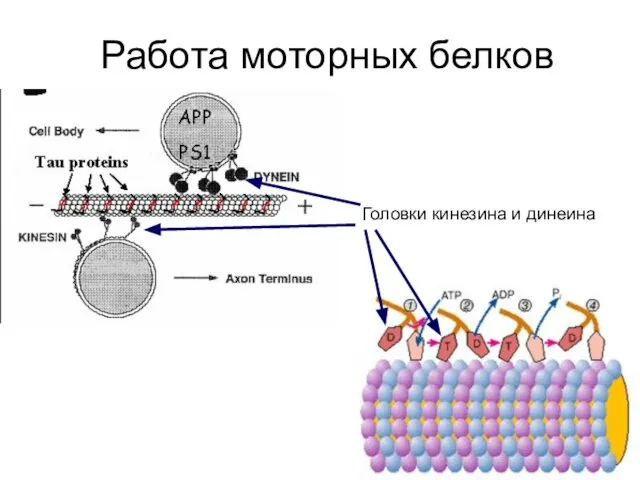
Работа моторных белков
Головки кинезина и динеина
Работа моторных белков
Головки кинезина и динеина
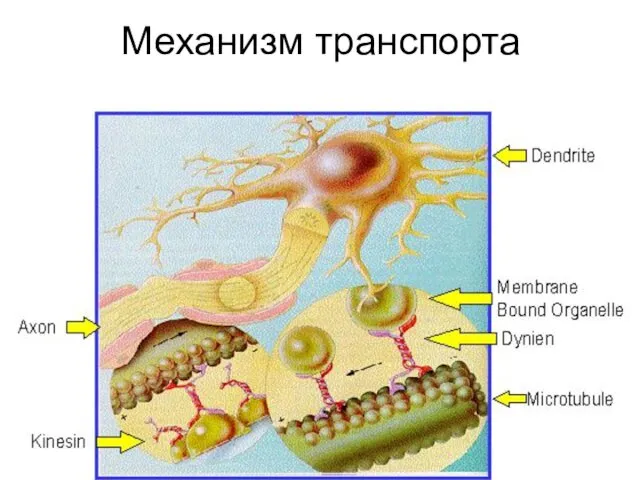
Механизм транспорта
Механизм транспорта

Ретроградный транспорт переносит:
Вирусы, бактерии, токсины,
Лекарства от терминалей аксона к телу нейрона
Ретроградный транспорт переносит:
Вирусы, бактерии, токсины,
Лекарства от терминалей аксона к телу нейрона
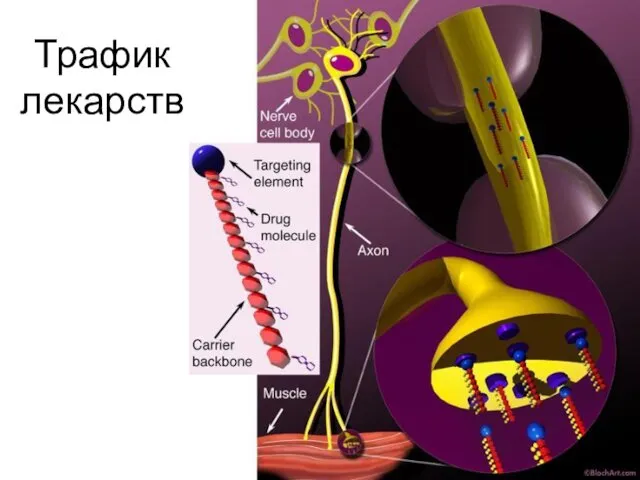
Трафик лекарств
Трафик лекарств

Ретроградный транспорт используется в научных целях
Для топической диагностики (локализации функций)
Ретроградный транспорт используется в научных целях
Для топической диагностики (локализации функций)
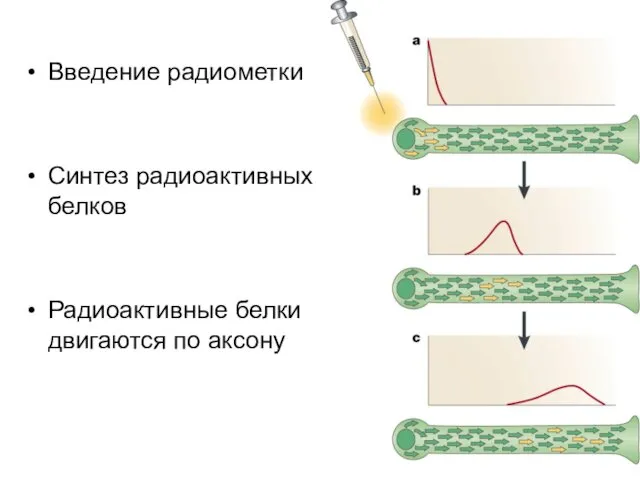
Введение радиометки
Синтез радиоактивных белков
Радиоактивные белки двигаются по аксону
Синтез радиоактивных белков
Радиоактивные белки двигаются по аксону
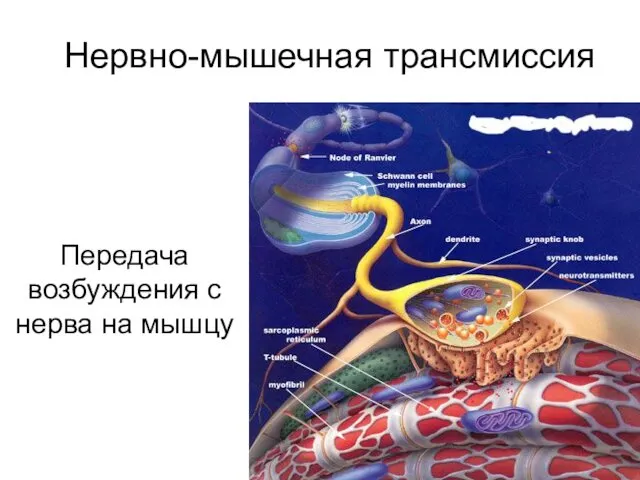
Нервно-мышечная трансмиссия
Передача возбуждения с нерва на мышцу
Нервно-мышечная трансмиссия
Передача возбуждения с нерва на мышцу

ПЕРЕДАЧА ВОЗБУЖДЕНИЯ С НЕРВА НА МЫШЦУ ПРОИСХОДИТ С ПОМОЩЬЮ СИНАПСА
Ч. Шеррингтон
ПЕРЕДАЧА ВОЗБУЖДЕНИЯ С НЕРВА НА МЫШЦУ ПРОИСХОДИТ С ПОМОЩЬЮ СИНАПСА
Ч. Шеррингтон

Физиологические свойства химических синапсов
Одностороннее проведение ПД.
Обязательное участие медиаторов.
Количество медиатора пропорционально частоте
Физиологические свойства химических синапсов
Одностороннее проведение ПД.
Обязательное участие медиаторов.
Количество медиатора пропорционально частоте
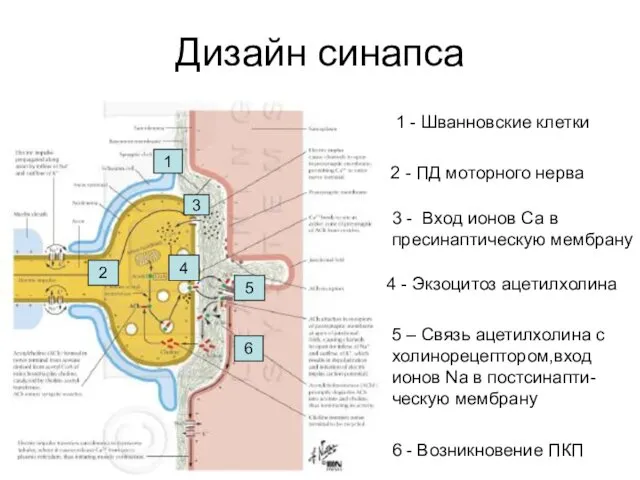
Дизайн синапса
1 - Шванновские клетки
2 - ПД моторного нерва
3 - Вход
Дизайн синапса
1 - Шванновские клетки
2 - ПД моторного нерва
3 - Вход
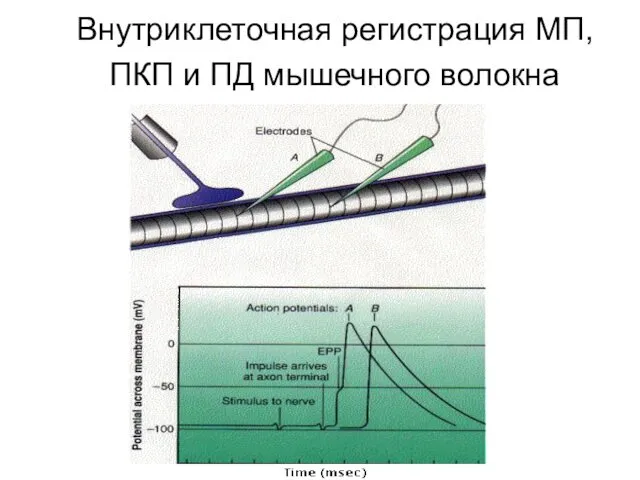
Внутриклеточная регистрация МП, ПКП и ПД мышечного волокна
Внутриклеточная регистрация МП, ПКП и ПД мышечного волокна

Функция синапса вне возбуждения
В покое существует связь между пре- и постсинаптическими
Функция синапса вне возбуждения
В покое существует связь между пре- и постсинаптическими
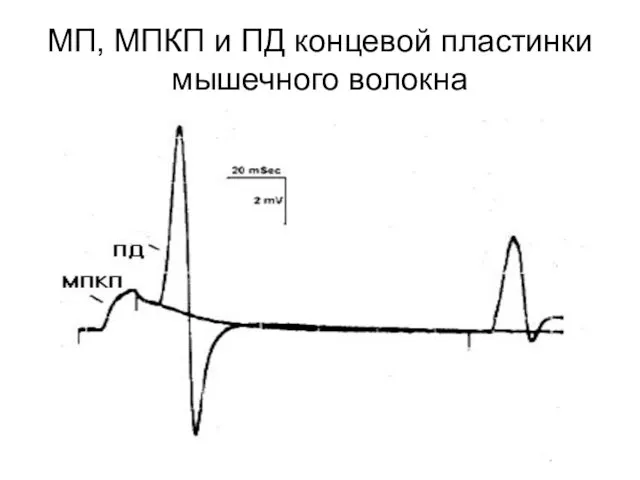
МП, МПКП и ПД концевой пластинки мышечного волокна
МП, МПКП и ПД концевой пластинки мышечного волокна
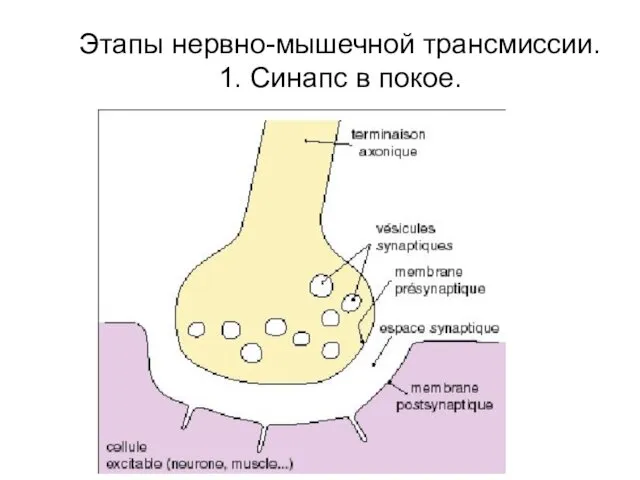
Этапы нервно-мышечной трансмиссии.
1. Синапс в покое.
Этапы нервно-мышечной трансмиссии.
1. Синапс в покое.
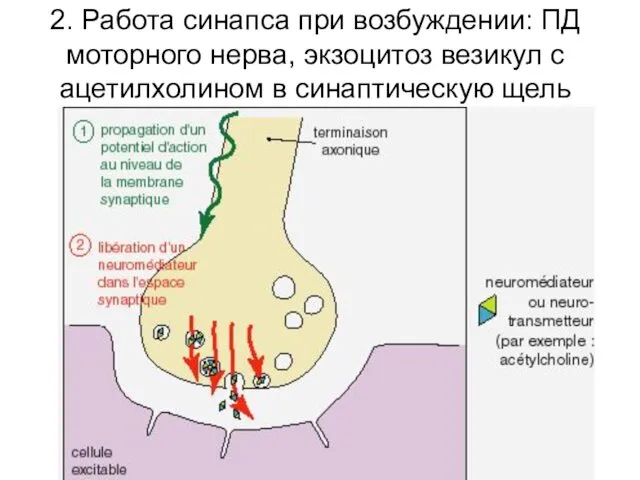
2. Работа синапса при возбуждении: ПД моторного нерва, экзоцитоз везикул с
2. Работа синапса при возбуждении: ПД моторного нерва, экзоцитоз везикул с
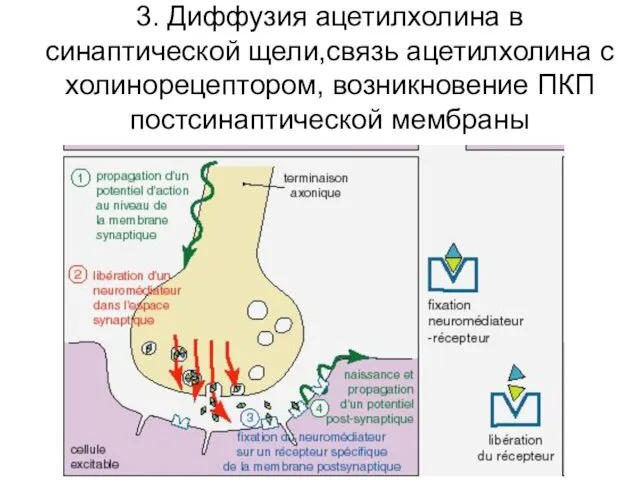
3. Диффузия ацетилхолина в синаптической щели,связь ацетилхолина с холинорецептором, возникновение ПКП
3. Диффузия ацетилхолина в синаптической щели,связь ацетилхолина с холинорецептором, возникновение ПКП
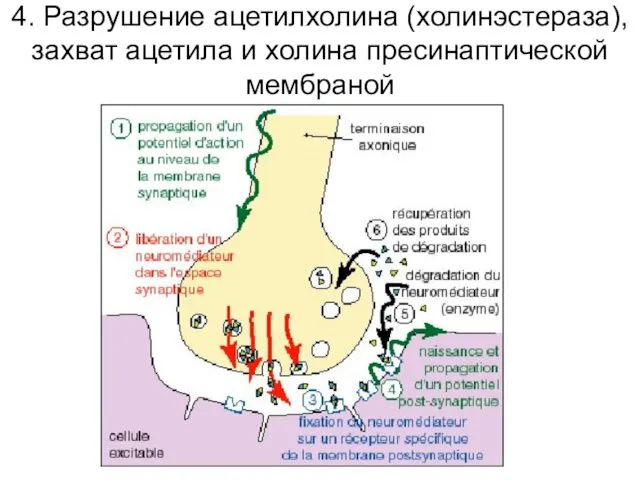
4. Разрушение ацетилхолина (холинэстераза), захват ацетила и холина пресинаптической мембраной
4. Разрушение ацетилхолина (холинэстераза), захват ацетила и холина пресинаптической мембраной
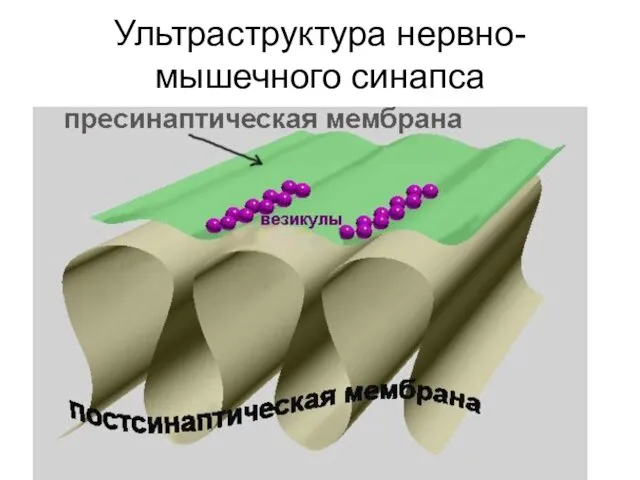
Ультраструктура нервно-мышечного синапса
Ультраструктура нервно-мышечного синапса
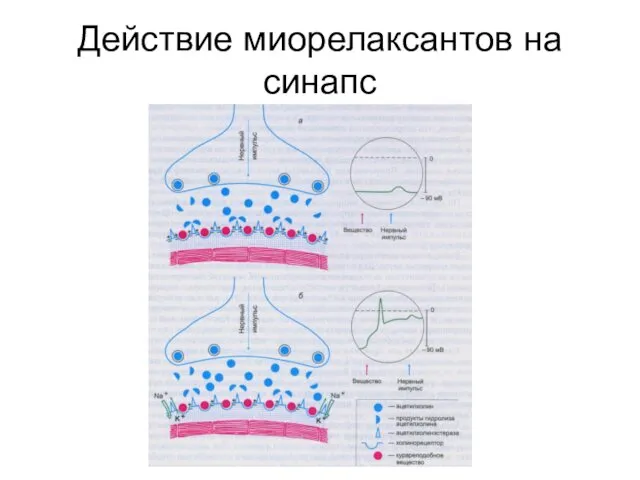
Действие миорелаксантов на синапс
Действие миорелаксантов на синапс

Ботокс – локальный миорелаксант
Большой Мол. вес (900 кДа), поэтому не распространяется
Ботокс – локальный миорелаксант
Большой Мол. вес (900 кДа), поэтому не распространяется
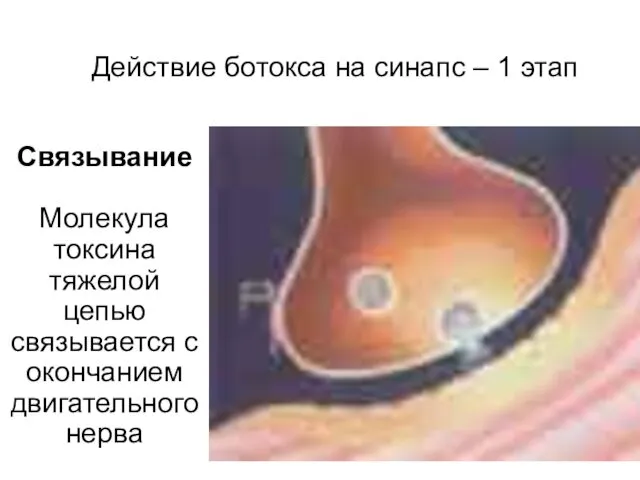
Действие ботокса на синапс – 1 этап
Связывание
Молекула токсина тяжелой цепью связывается
Действие ботокса на синапс – 1 этап
Связывание Молекула токсина тяжелой цепью связывается
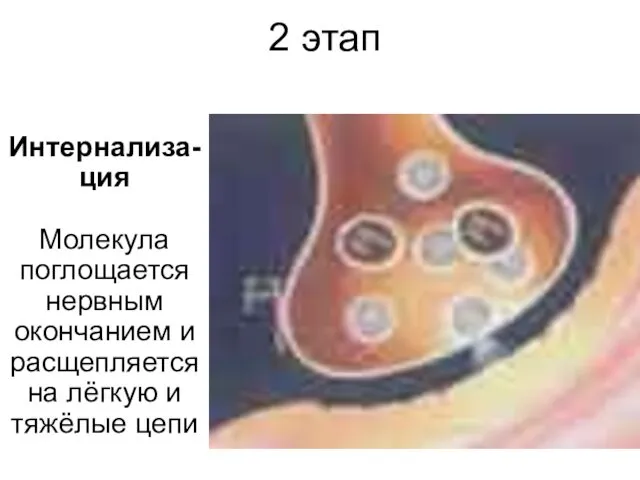
2 этап
Интернализа-ция
Молекула поглощается нервным окончанием и расщепляется на лёгкую и тяжёлые
2 этап
Интернализа-ция Молекула поглощается нервным окончанием и расщепляется на лёгкую и тяжёлые
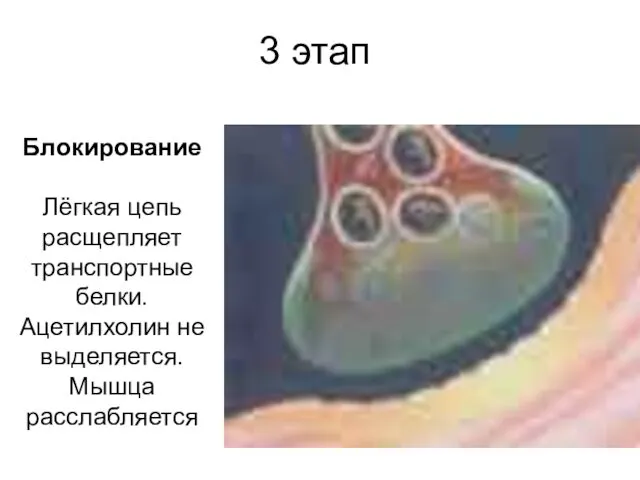
3 этап
Блокирование
Лёгкая цепь расщепляет транспортные белки. Ацетилхолин не выделяется. Мышца расслабляется
3 этап
Блокирование Лёгкая цепь расщепляет транспортные белки. Ацетилхолин не выделяется. Мышца расслабляется
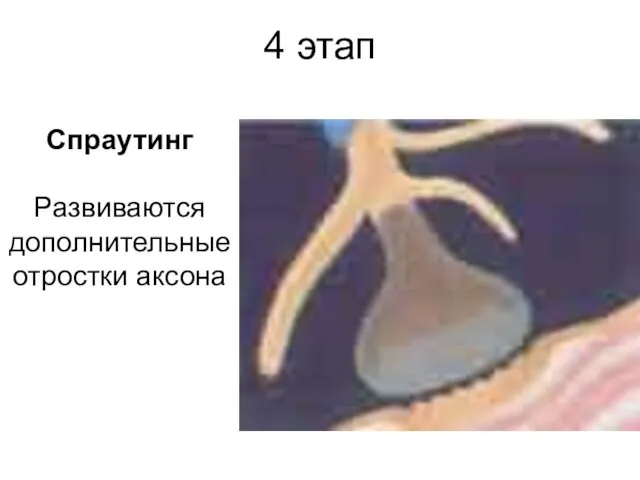
4 этап
Спраутинг
Развиваются дополнительные отростки аксона
4 этап
Спраутинг
Развиваются дополнительные отростки аксона
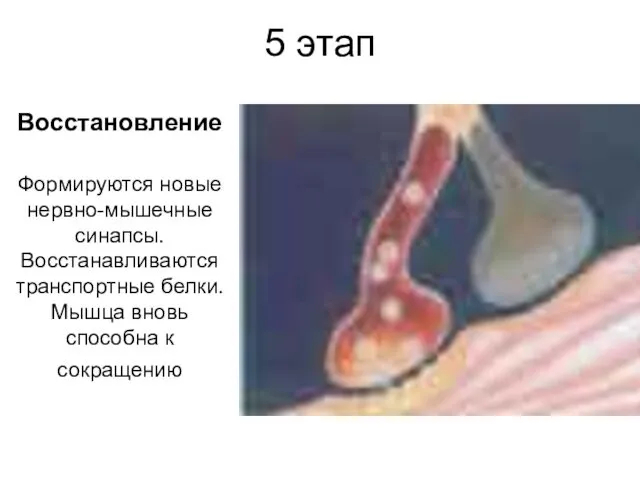
5 этап
Восстановление
Формируются новые нервно-мышечные синапсы. Восстанавливаются транспортные белки. Мышца вновь способна
5 этап
Восстановление Формируются новые нервно-мышечные синапсы. Восстанавливаются транспортные белки. Мышца вновь способна

Рецепторы – это чувствительные органы, способные реагировать на различные виды энергии
Рецепторы – это чувствительные органы, способные реагировать на различные виды энергии

3 функциональные роли рецепторов:
Сенсор (датчик) энергии раздражителя – воспринимает, чувствует любой
3 функциональные роли рецепторов:
Сенсор (датчик) энергии раздражителя – воспринимает, чувствует любой
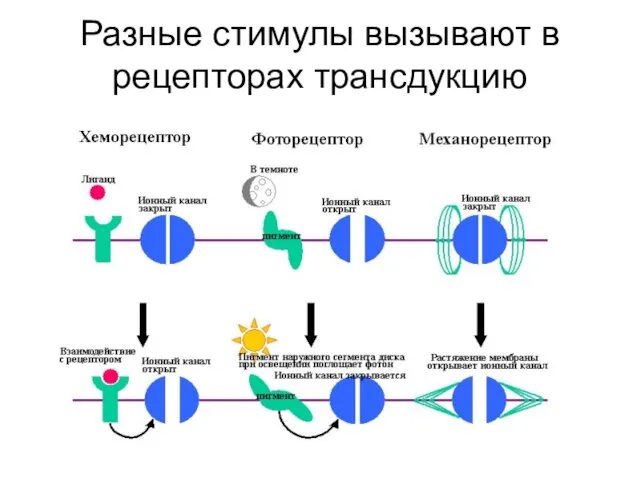
Разные стимулы вызывают в рецепторах трансдукцию
Разные стимулы вызывают в рецепторах трансдукцию
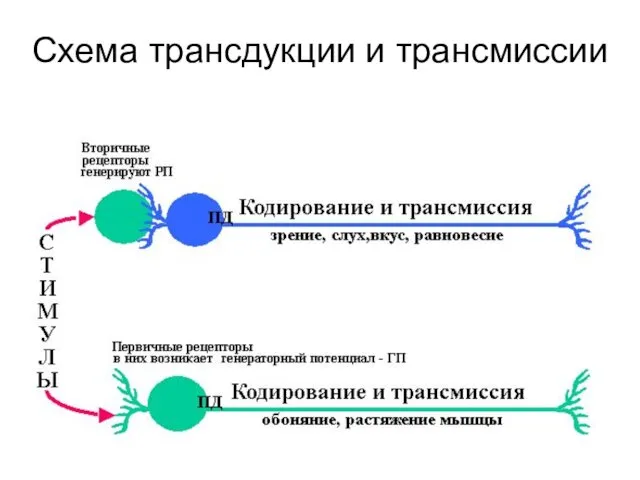
Схема трансдукции и трансмиссии
Схема трансдукции и трансмиссии

2 вида рецепторов
Первичные – это нервные клетки с периферически-чувствительным и центрально-проводящим
2 вида рецепторов
Первичные – это нервные клетки с периферически-чувствительным и центрально-проводящим

Сенсорная трансдукция, как правило, приводит к возникновению рецепторного потенциала -РП. Это
Сенсорная трансдукция, как правило, приводит к возникновению рецепторного потенциала -РП. Это
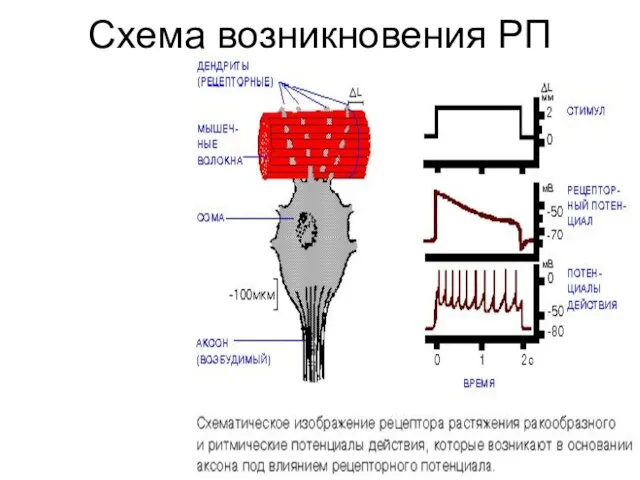
Схема возникновения РП
Схема возникновения РП
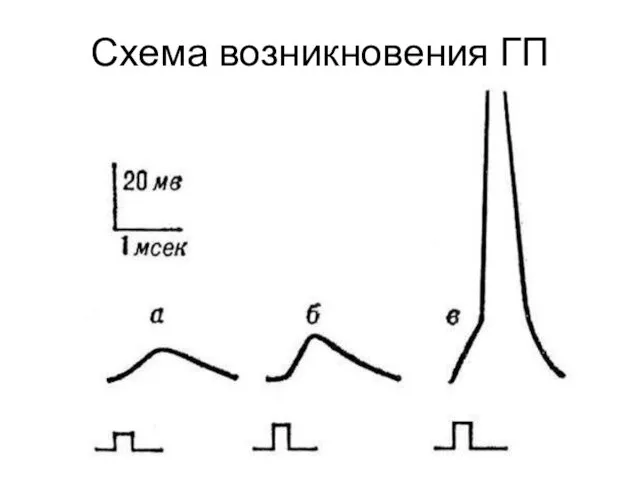
Схема возникновения ГП
Схема возникновения ГП
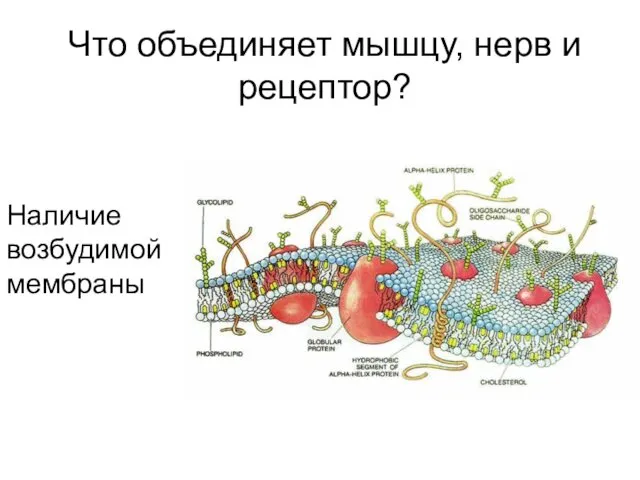
Что объединяет мышцу, нерв и рецептор?
Наличие возбудимой мембраны
Что объединяет мышцу, нерв и рецептор?
Наличие возбудимой мембраны

Физиология нервных волокон (аксонов)
Нервы являются отростками нервных клеток.
Нервы выполняют роль
Физиология нервных волокон (аксонов)
Нервы являются отростками нервных клеток.
Нервы выполняют роль
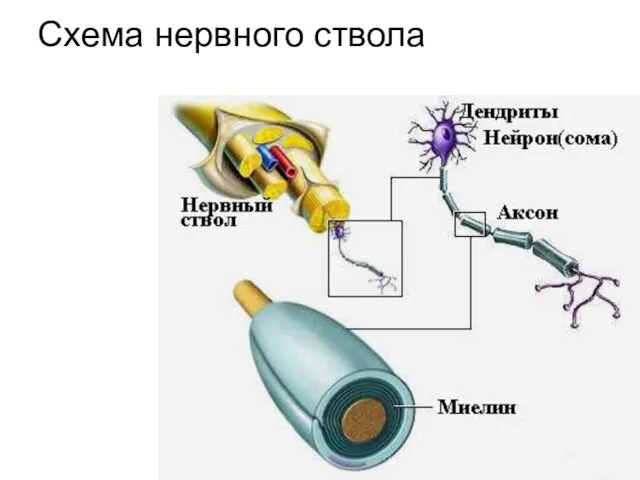
Схема нервного ствола
Схема нервного ствола

ФУНКЦИИ НЕРВОВ
Информационная - передача ПД от тела нейрона на периферию
ФУНКЦИИ НЕРВОВ
Информационная - передача ПД от тела нейрона на периферию
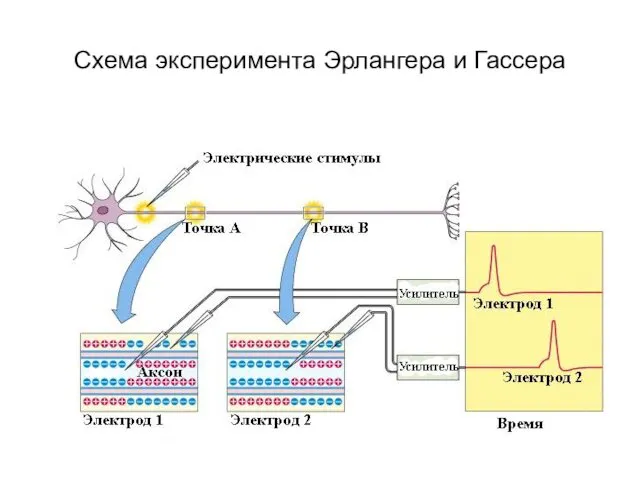
Схема эксперимента Эрлангера и Гассера
Схема эксперимента Эрлангера и Гассера
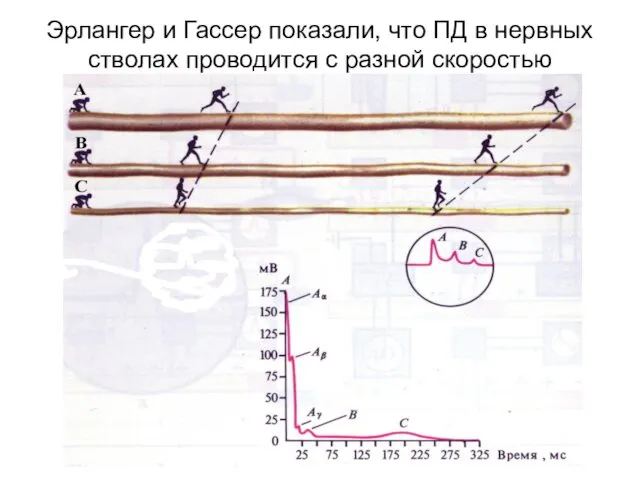
Эрлангер и Гассер показали, что ПД в нервных стволах проводится с
Эрлангер и Гассер показали, что ПД в нервных стволах проводится с

Классификация нервных волокон по Эрлангеру и Гассеру
А – волокна делятся на
Классификация нервных волокон по Эрлангеру и Гассеру
А – волокна делятся на
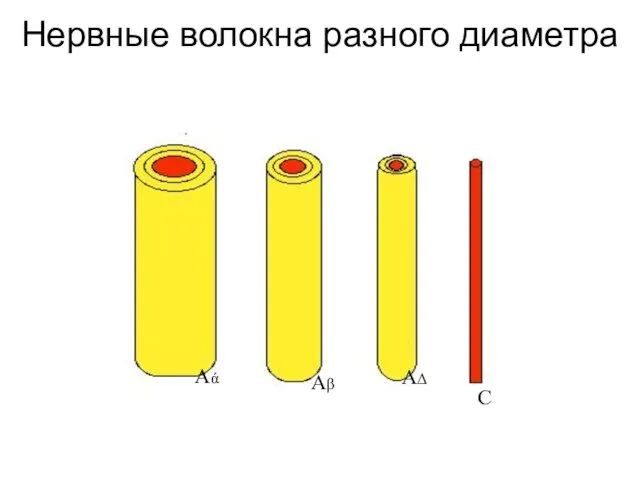
Нервные волокна разного диаметра
Аά
Аβ
АΔ
С
Нервные волокна разного диаметра
Аά
Аβ
АΔ
С
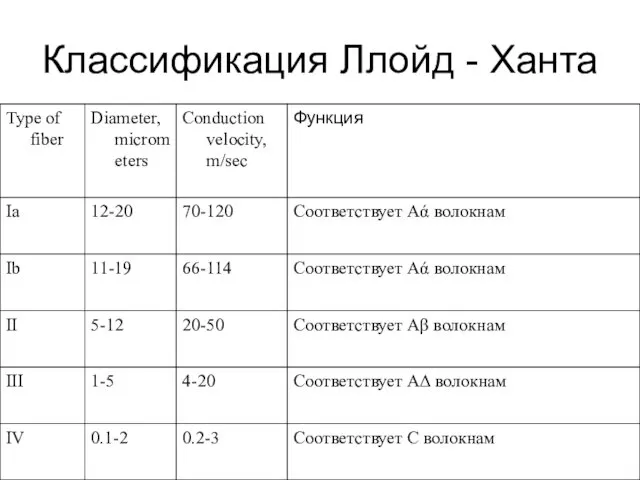
Классификация Ллойд - Ханта
Классификация Ллойд - Ханта

Законы проведения ПД по нерву
ПД по нерву распространяется в обе стороны.
ПД
Законы проведения ПД по нерву
ПД по нерву распространяется в обе стороны.
ПД

Продолжение
ПД распространяется изолированно.
Нерв практически не утомляем.
В разных по диаметру нервных волокнах
Продолжение
ПД распространяется изолированно.
Нерв практически не утомляем.
В разных по диаметру нервных волокнах
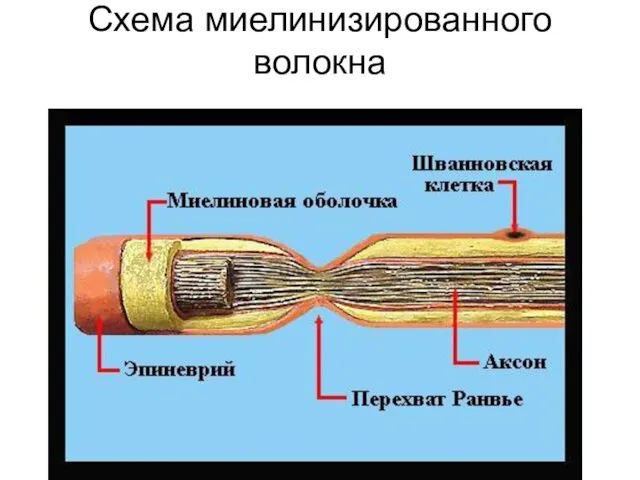
Схема миелинизированного волокна
Схема миелинизированного волокна

Проведение ПД происходит:
По миелиновому волокну – прыжками, сальтаторно.
По безмиелиновому волокну –
Проведение ПД происходит:
По миелиновому волокну – прыжками, сальтаторно.
По безмиелиновому волокну –
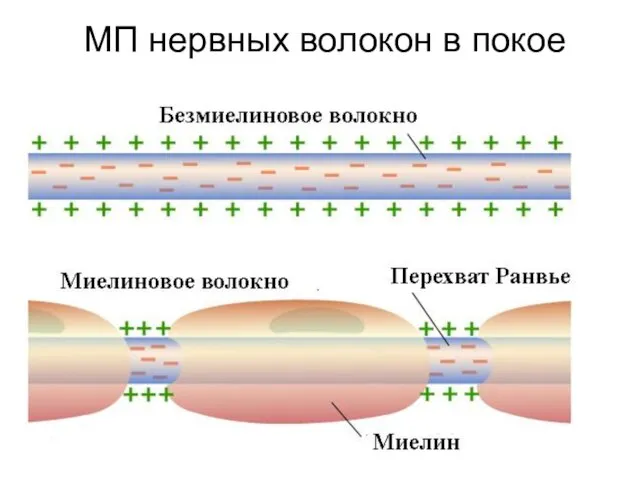
МП нервных волокон в покое
МП нервных волокон в покое
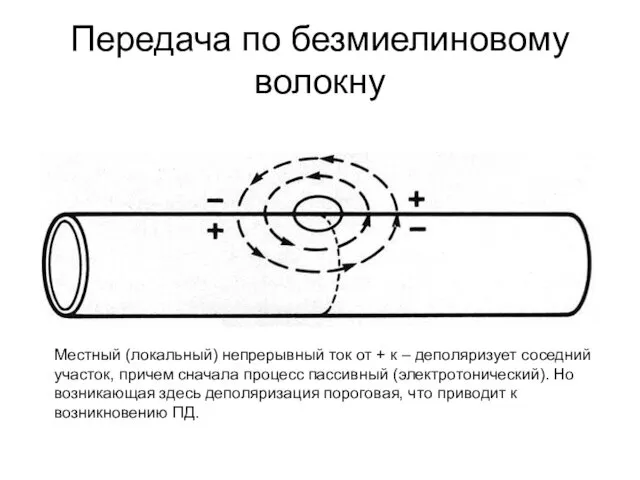
Передача по безмиелиновому волокну
Местный (локальный) непрерывный ток от + к –
Передача по безмиелиновому волокну
Местный (локальный) непрерывный ток от + к –
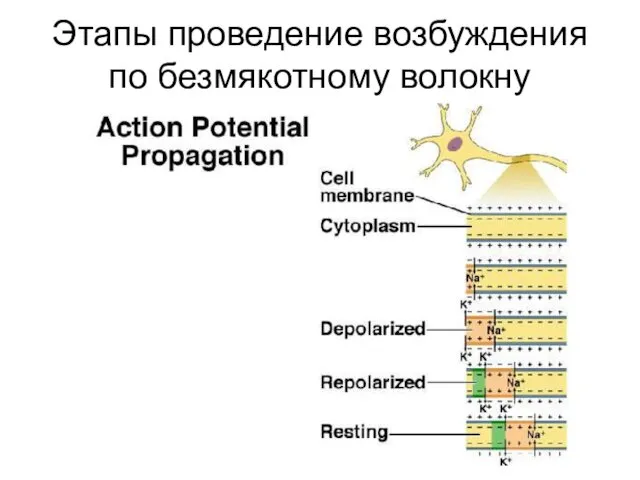
Этапы проведение возбуждения по безмякотному волокну
Этапы проведение возбуждения по безмякотному волокну
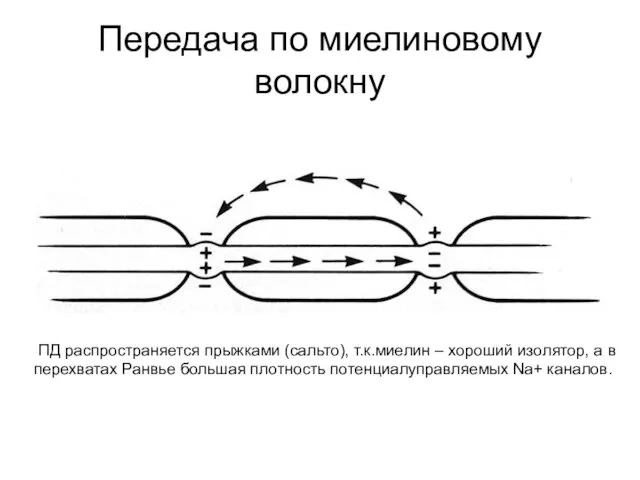
Передача по миелиновому волокну
ПД распространяется прыжками (сальто), т.к.миелин – хороший
Передача по миелиновому волокну
ПД распространяется прыжками (сальто), т.к.миелин – хороший
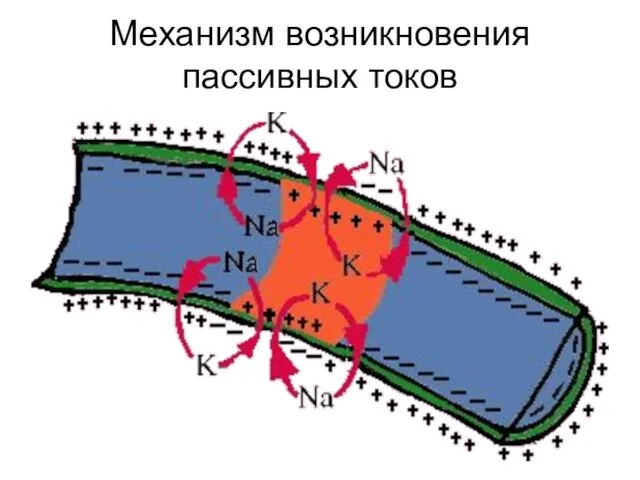
Механизм возникновения пассивных токов
Механизм возникновения пассивных токов
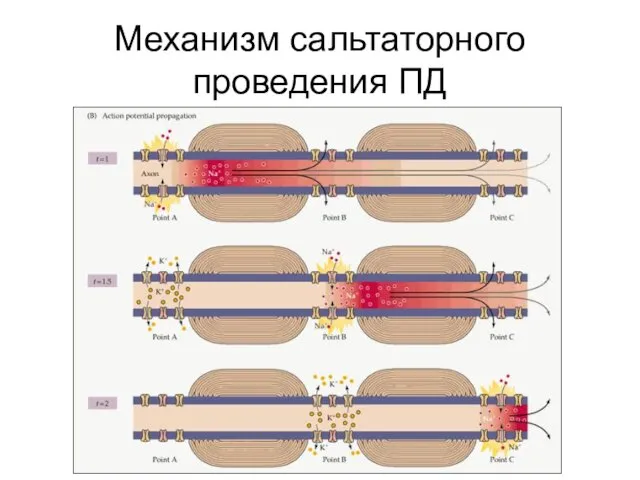
Механизм сальтаторного проведения ПД
Механизм сальтаторного проведения ПД
 Підготувала: учениця 7-А Щигельська В.
Підготувала: учениця 7-А Щигельська В. Лишайники. Разнообразие лишайников
Лишайники. Разнообразие лишайников Основные направления эволюции
Основные направления эволюции Грибы. Строение грибов
Грибы. Строение грибов Изменчивость. Изменение. Влияние
Изменчивость. Изменение. Влияние Общая характеристика грибов
Общая характеристика грибов Путешествие по эукариотической клетке: урок-игра
Путешествие по эукариотической клетке: урок-игра «Близорукость. Причины возникновения и профилактика заболевания» Автор презентации: Килязова Екатерина https://vk.com/kilyazova99
«Близорукость. Причины возникновения и профилактика заболевания» Автор презентации: Килязова Екатерина https://vk.com/kilyazova99  Строение простейших животных
Строение простейших животных ПТИЦЫ 2010-2011учебный год Потапова Полина ученица 6 «Б» класса ГОУ гимназии №42 Приморског
ПТИЦЫ 2010-2011учебный год Потапова Полина ученица 6 «Б» класса ГОУ гимназии №42 Приморског Интересное о птицах
Интересное о птицах Мезозойская эра в эволюции
Мезозойская эра в эволюции Этот прекрасный, чудесный, удивительный мир…
Этот прекрасный, чудесный, удивительный мир… Біосфера. Основні положення вчення В. Вернадського про біосферу
Біосфера. Основні положення вчення В. Вернадського про біосферу Кафедра микробиологии, вирусологии та имунологии Микробиологический контроль лекарственных средств в условиях аптечного пр
Кафедра микробиологии, вирусологии та имунологии Микробиологический контроль лекарственных средств в условиях аптечного пр Зелёная аптека
Зелёная аптека Семейство Злаки Обширное семейство злаков - одно из семейств класса однодольных. Злаки отличаются от других растений этого класса
Семейство Злаки Обширное семейство злаков - одно из семейств класса однодольных. Злаки отличаются от других растений этого класса Подотряд ящерицы
Подотряд ящерицы Использование физиологических признаков в селекции на засухоустойчивость Кудоярова Г.Р. Лаборатория физиологии растений, Инсти
Использование физиологических признаков в селекции на засухоустойчивость Кудоярова Г.Р. Лаборатория физиологии растений, Инсти Строение клетки
Строение клетки Аттестационная работа. Организация проектной деятельности на уроках окружающего мира
Аттестационная работа. Организация проектной деятельности на уроках окружающего мира Биологические особенности развития риса в Кызылординской области
Биологические особенности развития риса в Кызылординской области Обеспечение клеток энергией
Обеспечение клеток энергией Генетика. Структура генома эукариот, понятие о геномике. Локализация высокоповторенных последовательностей в хромосомах
Генетика. Структура генома эукариот, понятие о геномике. Локализация высокоповторенных последовательностей в хромосомах Ядро с сюрпризом
Ядро с сюрпризом Скелет головы и туловища
Скелет головы и туловища Основные закономерности эволюции
Основные закономерности эволюции Варакушка
Варакушка