- Физиология сенсорных систем

Содержание
- 2. У позвоночных животных в процессе эволюции возникло большое разнообразие сенсорных систем (анализаторов). Благодаря этому ЦНС получает
- 3. Классификация рецепторов Рецепторы Экстерорецепторы расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы
- 4. Рецепторы Первично-чувствующие (первичные) - преобразуют энергию внешнего стимула в возбуждение непосредственно на мембране рецептора Вторично-чувствующие (вторичные)
- 5. Примером может служить тельце Паччини (рис. 6.1 А)- Рецептор представляет собой немиелинизированное окончание аксона, одетого соединительнотканной
- 6. Во вторично-чувствующем рецепторе внешний стимул также вызывает генерацию рецепторного потенциала (рис. 6.1,Б). Афферентное волокно образует на
- 8. Слуховые, вестибулярные и другие типы рецепторов представлены волосковыми клетками, которые являются типичными вторичными рецепторами. Поверхность основания
- 9. Апикальная (верхняя) поверхность волосковой клетки находится в эндолимфе с высоким содержанием калия (рис. 6.2). Пучок волосков
- 10. Волосковый рецептор обладает направленной чувствительностью: смещение пучка в сторону киноцилии деполяризует клетку, а смещение в противоположную
- 11. Важным понятием сенсорной физиологии является порог. Различают абсолютный и дифференциальный (разностный) пороги. Абсолютным порогом считают минимальную
- 12. Это преобразование в подавляющем большинстве рецепторов носит нелинейный характер. Для ряда рецепторов было установлено, что передаточная
- 13. Другой широко распространенный способ кодирования в ЦНС получил название «кодирование номером линии». Этот тип кодирования хорошо
- 14. Большое число рецепторов спонтанно разряжаются в отсутствие каких-либо раздражителей. Например, рецепторы вестибулярного аппарата постоянно активны, что
- 15. При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это явление называется адаптацией. В адаптации
- 17. Отдельное афферентное нервное волокно 1-го порядка собирает информацию с более или менее широкой области поля рецепторов.
- 18. Одним из важных принципов взаимодействия в нейронных сетях является латеральное торможение: возбужденные элементы (рецепторы, нейроны) через
- 19. В состав сенсорной системы, кроме описанного 1-го уровня обработки информации, входит также ряд других структур головного
- 20. Кора головного мозга млекопитающих, покрывающая большие полушария конечного мозга, благодаря многочисленным складкам может иметь значительную площадь;
- 22. В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные зоны коры. В проекционных зонах
- 24. Скачать презентацию
У позвоночных животных в процессе эволюции возникло большое разнообразие сенсорных систем
У позвоночных животных в процессе эволюции возникло большое разнообразие сенсорных систем
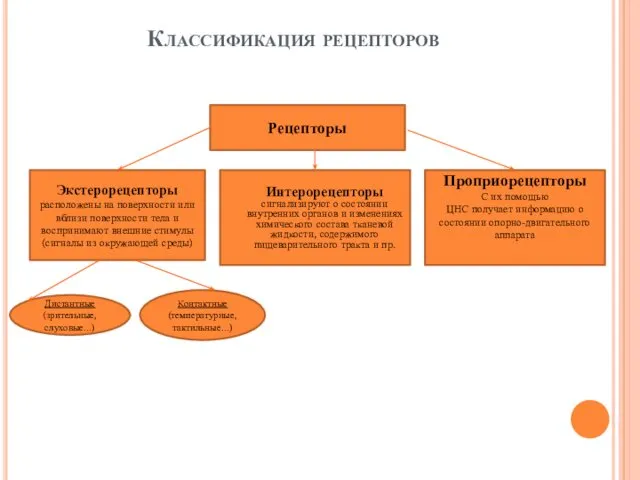
Классификация рецепторов
Рецепторы
Экстерорецепторы
расположены на поверхности или вблизи поверхности тела и воспринимают внешние
Классификация рецепторов
Рецепторы
Экстерорецепторы
расположены на поверхности или вблизи поверхности тела и воспринимают внешние

Рецепторы
Первично-чувствующие (первичные) - преобразуют энергию внешнего стимула в возбуждение непосредственно на
Рецепторы
Первично-чувствующие (первичные) - преобразуют энергию внешнего стимула в возбуждение непосредственно на
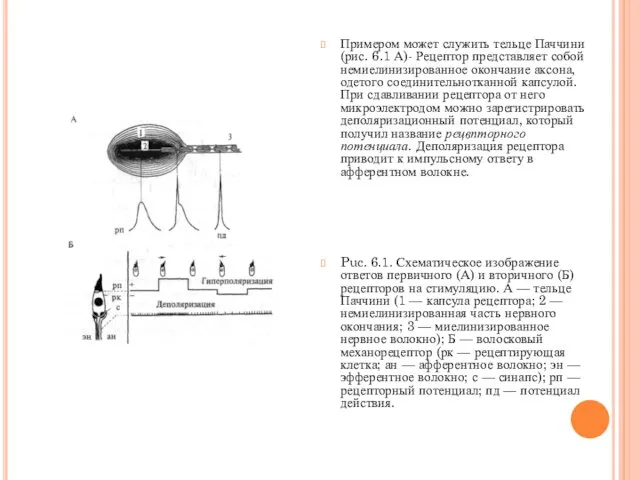
Примером может служить тельце Паччини (рис. 6.1 А)- Рецептор представляет собой
Примером может служить тельце Паччини (рис. 6.1 А)- Рецептор представляет собой
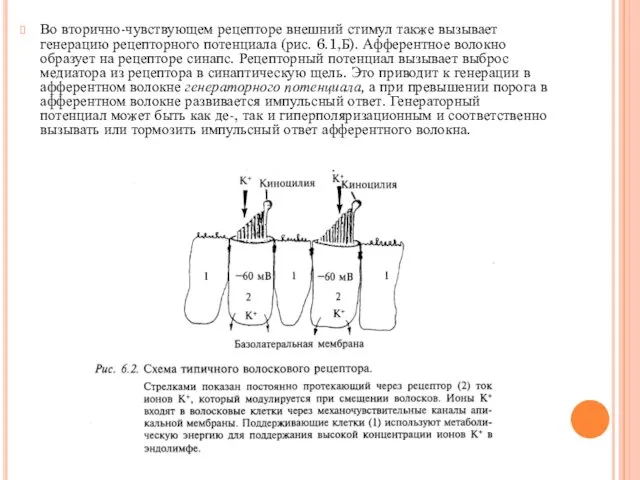
Во вторично-чувствующем рецепторе внешний стимул также вызывает генерацию рецепторного потенциала (рис.
Во вторично-чувствующем рецепторе внешний стимул также вызывает генерацию рецепторного потенциала (рис.


Слуховые, вестибулярные и другие типы рецепторов представлены волосковыми клетками, которые являются
Слуховые, вестибулярные и другие типы рецепторов представлены волосковыми клетками, которые являются
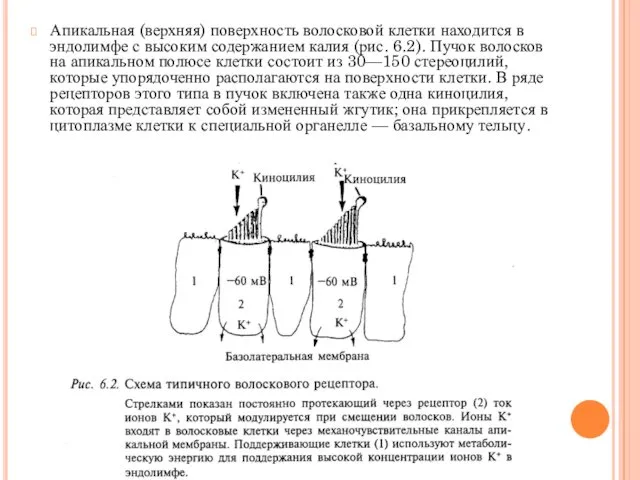
Апикальная (верхняя) поверхность волосковой клетки находится в эндолимфе с высоким содержанием
Апикальная (верхняя) поверхность волосковой клетки находится в эндолимфе с высоким содержанием

Волосковый рецептор обладает направленной чувствительностью: смещение пучка в сторону киноцилии деполяризует
Волосковый рецептор обладает направленной чувствительностью: смещение пучка в сторону киноцилии деполяризует
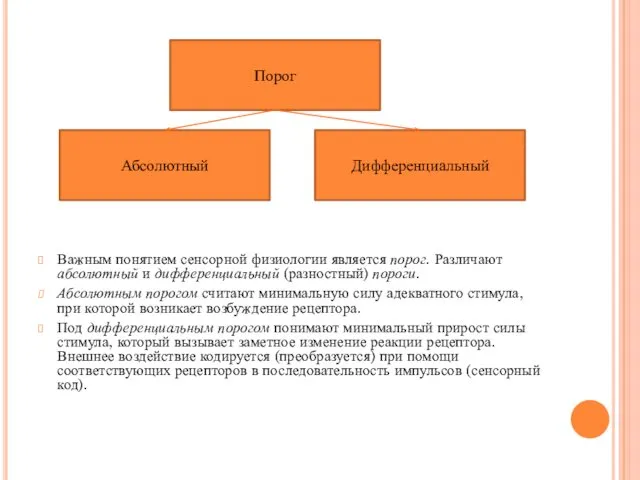
Важным понятием сенсорной физиологии является порог. Различают абсолютный и дифференциальный (разностный)
Важным понятием сенсорной физиологии является порог. Различают абсолютный и дифференциальный (разностный)
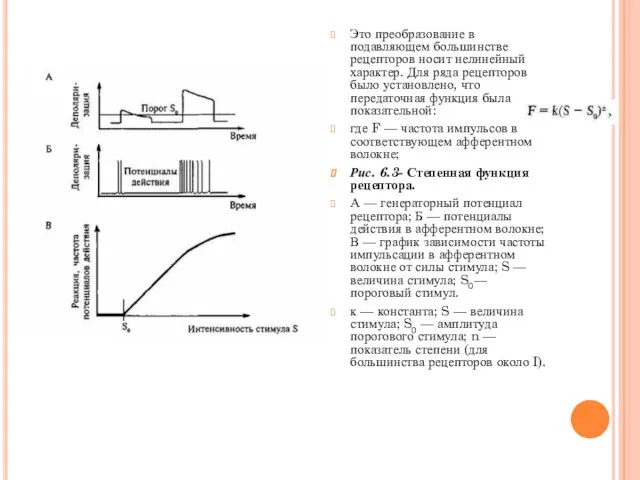
Это преобразование в подавляющем большинстве рецепторов носит нелинейный характер. Для ряда
Это преобразование в подавляющем большинстве рецепторов носит нелинейный характер. Для ряда
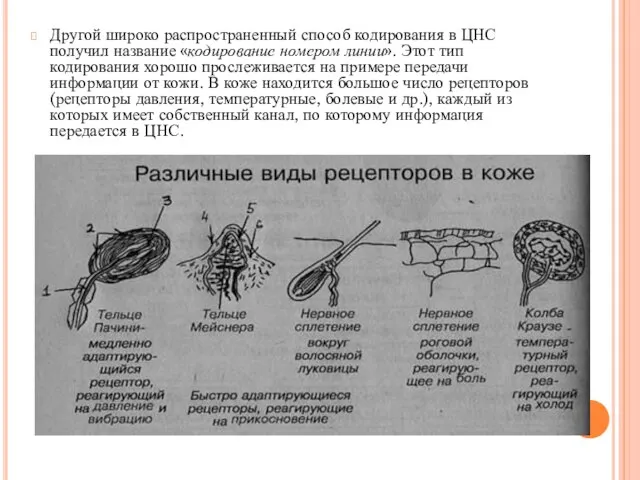
Другой широко распространенный способ кодирования в ЦНС получил название «кодирование номером
Другой широко распространенный способ кодирования в ЦНС получил название «кодирование номером
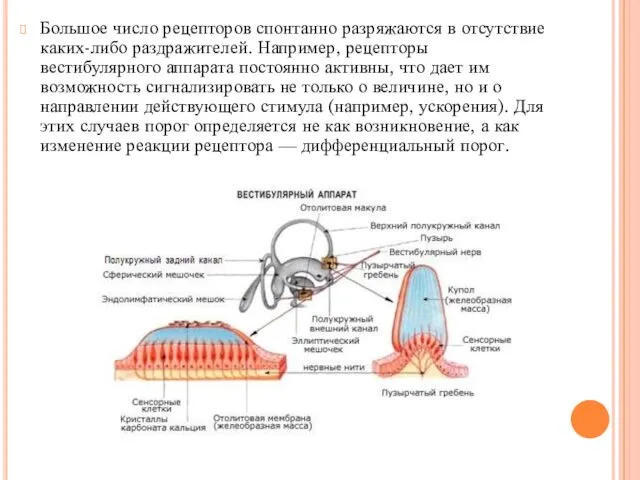
Большое число рецепторов спонтанно разряжаются в отсутствие каких-либо раздражителей. Например, рецепторы
Большое число рецепторов спонтанно разряжаются в отсутствие каких-либо раздражителей. Например, рецепторы

При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это
При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это


Отдельное афферентное нервное волокно 1-го порядка собирает информацию с более или
Отдельное афферентное нервное волокно 1-го порядка собирает информацию с более или
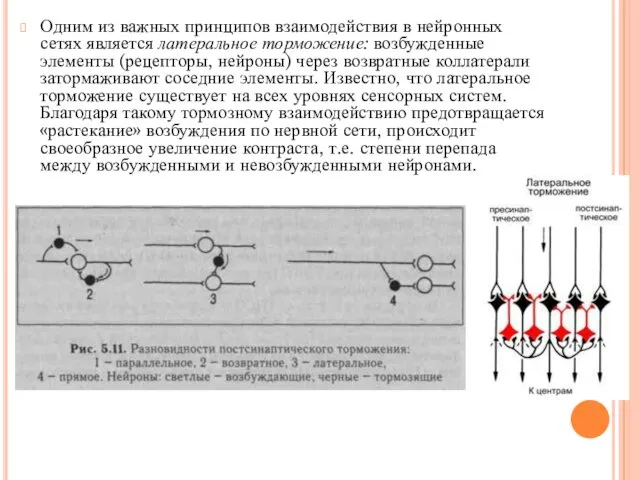
Одним из важных принципов взаимодействия в нейронных сетях является латеральное торможение:
Одним из важных принципов взаимодействия в нейронных сетях является латеральное торможение:
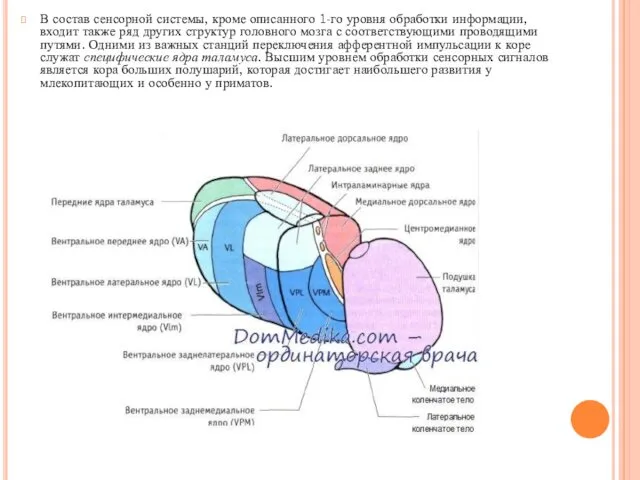
В состав сенсорной системы, кроме описанного 1-го уровня обработки информации, входит
В состав сенсорной системы, кроме описанного 1-го уровня обработки информации, входит
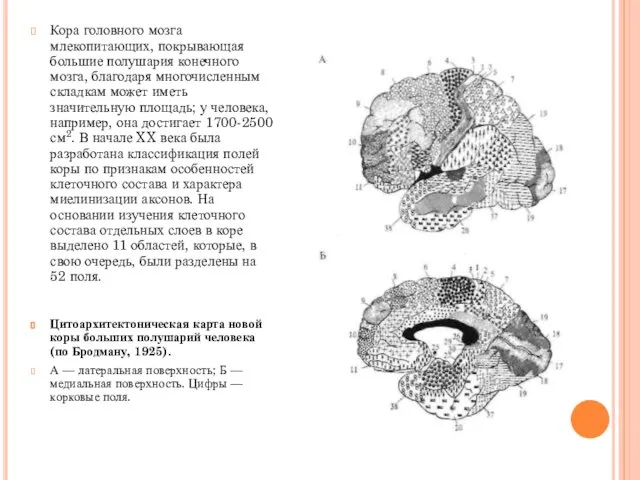
Кора головного мозга млекопитающих, покрывающая большие полушария конечного мозга, благодаря многочисленным
Кора головного мозга млекопитающих, покрывающая большие полушария конечного мозга, благодаря многочисленным

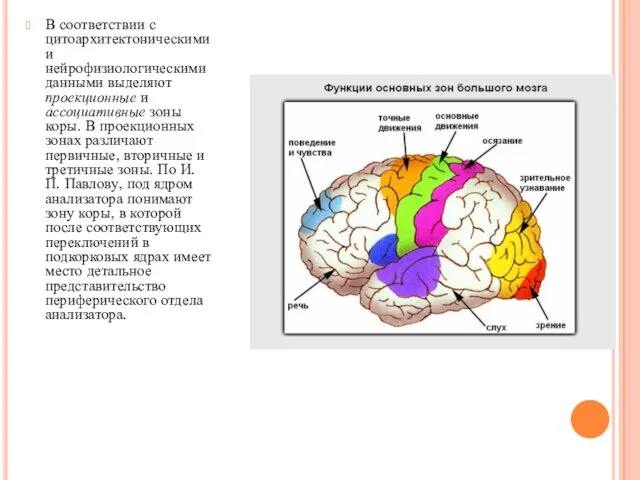
В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные
В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные
 Растения-паразиты Презентация к уроку экологии в 6 классе
Растения-паразиты Презентация к уроку экологии в 6 классе  Лекция № 15 Ценологическая геоботаника (Фитоценология) Определение фитоценоза. Теория сообществ-единиц (организмизм) Индивидуал
Лекция № 15 Ценологическая геоботаника (Фитоценология) Определение фитоценоза. Теория сообществ-единиц (организмизм) Индивидуал Презентация на тему "Турнир юных биологов" - скачать презентации по Биологии
Презентация на тему "Турнир юных биологов" - скачать презентации по Биологии Моллюски. Удивительные существа
Моллюски. Удивительные существа История
История Система опоры и движения
Система опоры и движения Эксплуатационные свойства газонных покрытий и установление качества дернины и травостоя по их параметрам
Эксплуатационные свойства газонных покрытий и установление качества дернины и травостоя по их параметрам Железы организма человека
Железы организма человека Царство Животные
Царство Животные Мейоз. Особенности мейоза. Механизм и биологическое значение митоза и мейоза
Мейоз. Особенности мейоза. Механизм и биологическое значение митоза и мейоза Дикие животные
Дикие животные Строение мышцы
Строение мышцы Выполнили: Чикунов Вячеслав, Болтовская Элина 4 в класс МОУ СОШ № 11
Выполнили: Чикунов Вячеслав, Болтовская Элина 4 в класс МОУ СОШ № 11 Как животные леса готовятся к зиме
Как животные леса готовятся к зиме Биология клетки
Биология клетки Тип Плоские черви, Круглые черви, Кольчатые черви
Тип Плоские черви, Круглые черви, Кольчатые черви Гибридизация как метод селекции
Гибридизация как метод селекции Значение птиц в природе и жизни человека
Значение птиц в природе и жизни человека Споры и Спорангии
Споры и Спорангии Сохранить планету для людей
Сохранить планету для людей РЕПТИЛИИ
РЕПТИЛИИ Фикус
Фикус  Лучше один раз увидеть?
Лучше один раз увидеть? Многообразие биогеоценозов в природе. (10 класс)
Многообразие биогеоценозов в природе. (10 класс) Загадка человека. Урок 3
Загадка человека. Урок 3 Презентация на тему "Видоизменённые надземные побеги" - скачать презентации по Биологии
Презентация на тему "Видоизменённые надземные побеги" - скачать презентации по Биологии Презентация на тему "Отдел покрытосеменные" - скачать презентации по Биологии
Презентация на тему "Отдел покрытосеменные" - скачать презентации по Биологии КРЫСЫ- соседи и враги
КРЫСЫ- соседи и враги