- Генетический контроль плана развития организма

Содержание
- 2. Классические опыты, показывающие роль цитоплазмы ооцита в установлении двух “organizing centers” - центров формирования полярности эмбриона
- 3. СРАВНЕНИЕ СЕГМЕНТАЦИИ ЭМБРИОНА И ИМАГО ДРОЗОФИЛЫ У эмбриона уже определены оси полярности, число и ориентация сегментов,
- 4. Становление плана строения тела и классы основных генов контролирующих эмбриональное развитие 1. Образование передне-задней и спинно-брюшной
- 5. Ранний эмбриогенез дрозофилы Ядро зиготы дрозофилы претерпевают 13 синхронных делений без цитокинеза, образуя зародыш с ~6000
- 6. Оплодотворенное яйцо Миграция ядер к периферии Синцитиальная бластодерма Деления ядер – образования синцития Синцитиальная бластодерма Клеточная
- 7. Принцип поиска мутантов по материнским генам Мутагенез Гомозиготные мутанты живут, Погибшие эмбрионы но стерильны. имеют структурные
- 8. Первая «передняя» система морфогенов, определяет области головы и груди. Вторая «задняя» система морфогенов определяет сегментацию абдоминальной
- 9. K. Kalthoff and K. Sander. 1968. Wilhelm Roux's Arch. Entw Mech. Org. 161: 129-146. W. Driever,
- 10. Дикий тип Нарушения А-градиентов Нарушения Р-градиента Эксперименты Нюссляйн-Вольхард по трансплантации на эмбрионах дрозофилы
- 11. Принцип поиска мутантов по генам сегментации Мутагенез Гетерозиготы живут Погибшие гомозиготные эмбрионы . имеют дефекты сегментации
- 12. Препарат кутикулы эмбриона дрозофилы дикого типа А Р Вид с латеральной стороны Вид с вентральной стороны
- 13. Фенотипы кутикулы эмбрионов с нарушенной сегментацией вдоль А/Р-оси В середине – дикий тип. Слева мутант по
- 14. Каким образом формируются градиенты морфогенов в яйце? Как поляризуется сам ооцит?
- 15. Строение репродуктивной системы самки дрозофилы Парный яичник Овариола гермарий овариола трофоциты ооцит 200 мкм 20 мкм
- 16. Строение гермария Обозначения: ТФ – терминальный филамент ГСК – герминативные стволовые клетки ВК – верхушечные клетки
- 17. Ранний оогенез дрозофилы Когда циста достигнет зоны 2б, одна клетка получит статус ооцита. Первое, ооцит-специфичные белки
- 18. Фьюсома происходит из сферической структуры, названной спектросома, которая характерна герминативным стволовым клеткам (ГСК). Она состоит из
- 19. Nicole C. Grieder, Margaret de Cuevas and Allan C. Spradling. «The fusome organizes the microtubule network
- 20. Следствие полярности фьюсомы Циста в зоне 2а дифференцируется в двух направлениях: 1. цитоплазматическом Во-первых, фьюсома организовывает
- 21. Передняя область гермария мутанта: спектросомы и фьюсомы без актина. ниша Стволовые клетки Гермарий Стволовые герминативные клетки
- 22. Нарушение ранних стадий оогенеза у мутантов с аномальной фьюсомой Мутант Яйцевые камеры отсутствуют. Разросшиеся гермарии заполнены
- 23. Установление A/P полярности ооцита Когда герминативная циста достигает зоны 2б, транспортируемые в ооцит вдоль фьюсомы специфические
- 24. Система PAR-aPKC вовлечена в разные типы поляризации клеток
- 25. Current Biology, Vol. 14, 520–525, March 23, 2004, Асимметричная локализация двух белков PAR6 во время ранней
- 26. Роль кадгерина в формировании А/Р полярных осей Локализация ооцита в заднем отделе яйцевой камеры происходит благодаря
- 27. Регуляторы цитоплазматической дифференцировки обозначены красным, регуляторы ядерной дифференцировки обозначены синим цветом. Вверху представлена 4-клеточная циста (красная
- 28. Фьюсома устанавливает оси полярности очень рано, в зоне 1 гермария Затем в зоне 3 гермария белки
- 29. Ранний оогенез у дрозофилы и ксенопуса У дрозофилы и у ксенопуса ооцит наследует передне-задние оси симметрии
- 30. Я - тоже муха: Мой краток век. А чем ты, муха, Не человек? ВИЛЬЯМ БЛЕЙК. "Муха"
- 31. 1995 Нобелевская премия по физиологии и медицине «За открытия, касающиеся генетического контроля на ранней стадии эмбрионального
- 32. Первая «передняя» система морфогенов, определяет области головы и груди. Вторая «задняя» система морфогенов определяет сегментацию абдоминальной
- 33. Вдоль АР оси индивидуальные области формируются независимо. Если одновременно убрать функции двух АР систем, то останутся
- 34. Общие свойства четырёх систем Продукт одного гена каждой системы локализован в специфической области свежеотложенного яйца и
- 35. Формирование А/Р оси ооцита Drosophila (А) Ооцит перемещается в задний отдел яйцевой камеры, а фолликулярные клетки
- 36. Локализация продуктов генов gurken, bicoid и oskar в ооците мРНК Белок gurken-мРНК накапливается между ядром ооцита
- 37. Антериорная система – морфогенетический градиент А-система является самой простой. Только один ген фактора транскрипции bicoid (bcd)
- 38. Экспериментальная демонстрация морфогенетической индукции головных структур геном bicoid «Да у него две головы: одна спереди, другая
- 39. Формирование градиента в заднем отделе эмбриона Центральный компонент Р-системы – продукт гена nanos. Сначала перемещаются мРНК
- 40. Модель распределения градиентов продуктов материнских генов вдоль передне-задней оси Два других материнских гена hunchback и caudal
- 41. Схема двойной репрессии продуктов генов caudal и hunchback вдоль передне-задней оси эмбриона Передний полюс Задний полюс
- 42. Терминальная система – локальная активация рецептора Главную роль в установлении АР-полярности играют герминативные клетки – трофоциты.
- 43. Схема активации рецептора Torso Белок Torso-like экспрессируется фолликулярными клетками на полюсах ооцита. мРНК Torso экспрессируется вокруг
- 44. Активация рецептора Torso на полюсах яйца происходит в результате сигнальной трансдукции Активация рецептора Torso на полюсах
- 45. Цель Т-системы: инактивация транскрипционного супрессора «терминальных» зиготических генов huckebein (hkb) и tailless (tll) Torso инактивирует комплекс
- 46. Механизм Torso-сигнальной трансдукции Willis. 2005. Functions and Mechanisms of Receptor Tyrosine Kinase Torso Signaling: Lessons From
- 48. Скачать презентацию

Классические опыты, показывающие роль цитоплазмы ооцита в установлении двух “organizing centers”
Классические опыты, показывающие роль цитоплазмы ооцита в установлении двух “organizing centers”
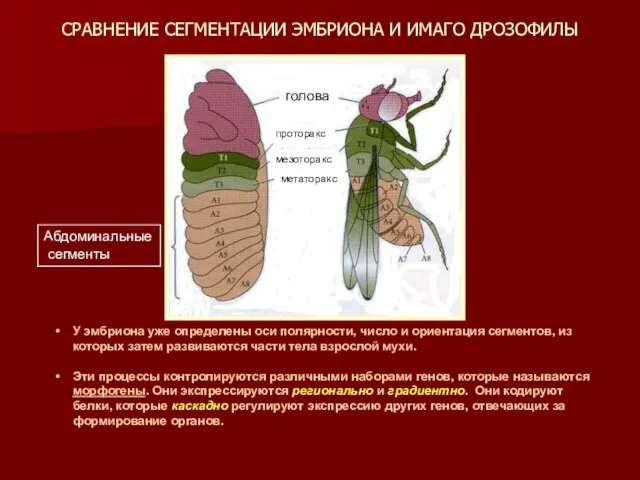
СРАВНЕНИЕ СЕГМЕНТАЦИИ ЭМБРИОНА И ИМАГО ДРОЗОФИЛЫ
У эмбриона уже определены оси полярности,
СРАВНЕНИЕ СЕГМЕНТАЦИИ ЭМБРИОНА И ИМАГО ДРОЗОФИЛЫ
У эмбриона уже определены оси полярности,
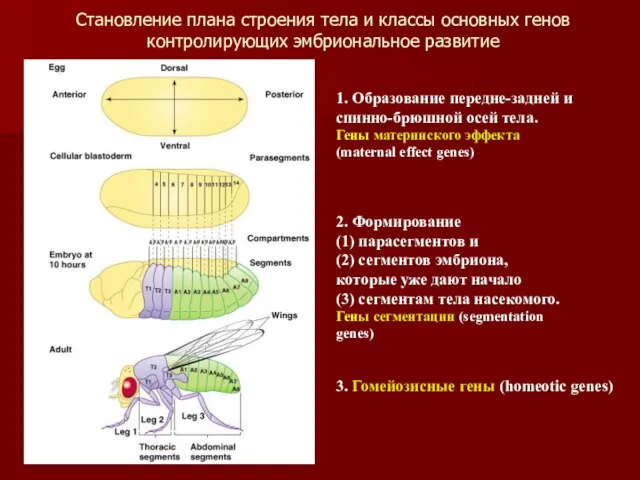
Становление плана строения тела и классы основных генов
контролирующих эмбриональное развитие
1. Образование
Становление плана строения тела и классы основных генов
контролирующих эмбриональное развитие
1. Образование
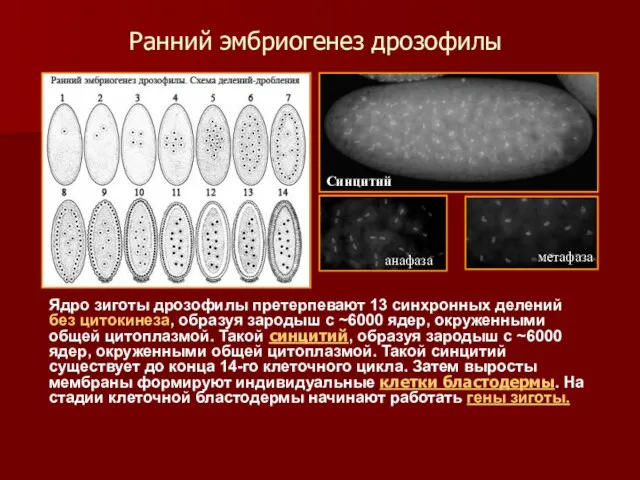
Ранний эмбриогенез дрозофилы
Ядро зиготы дрозофилы претерпевают 13 синхронных делений без цитокинеза,
Ранний эмбриогенез дрозофилы
Ядро зиготы дрозофилы претерпевают 13 синхронных делений без цитокинеза,
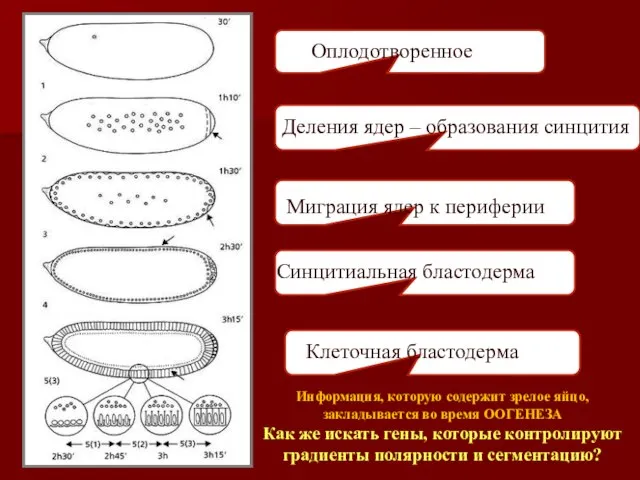
Оплодотворенное яйцо
Миграция ядер к периферии
Синцитиальная бластодерма
Деления ядер – образования синцития
Синцитиальная бластодерма
Клеточная
Оплодотворенное яйцо
Миграция ядер к периферии
Синцитиальная бластодерма
Деления ядер – образования синцития
Синцитиальная бластодерма
Клеточная
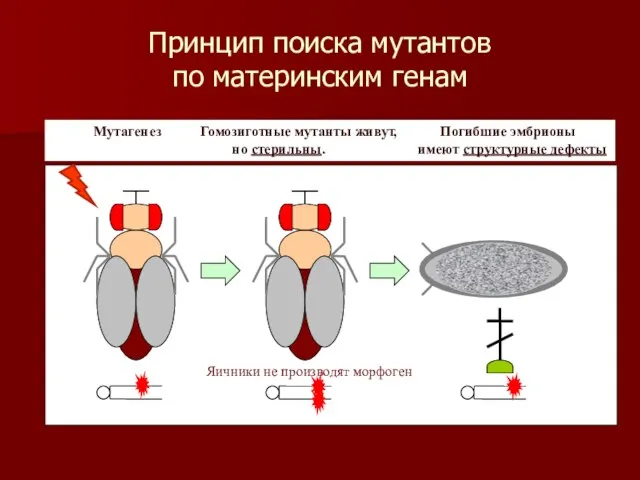
Принцип поиска мутантов
по материнским генам
Мутагенез Гомозиготные мутанты живут, Погибшие
Принцип поиска мутантов
по материнским генам
Мутагенез Гомозиготные мутанты живут, Погибшие
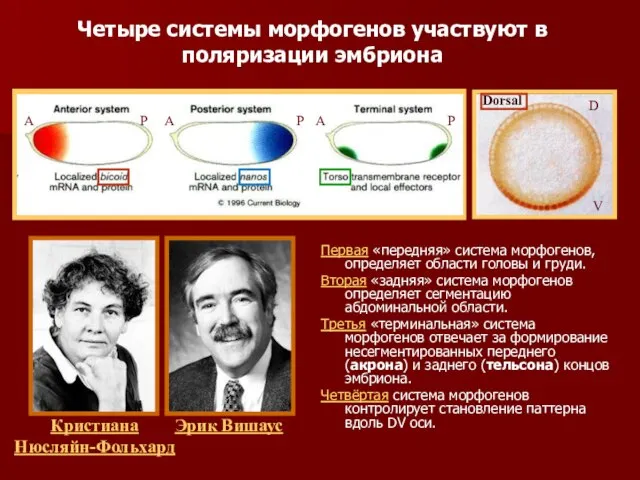
Первая «передняя» система морфогенов, определяет области головы и груди.
Вторая «задняя»
Вторая «задняя»
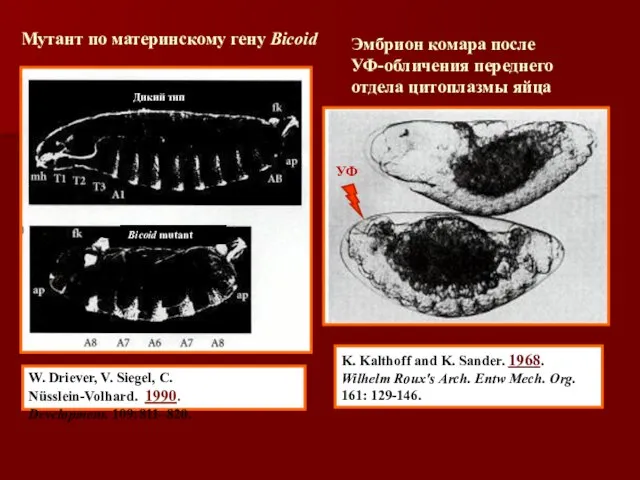
K. Kalthoff and K. Sander. 1968. Wilhelm Roux's Arch. Entw Mech.
K. Kalthoff and K. Sander. 1968. Wilhelm Roux's Arch. Entw Mech.
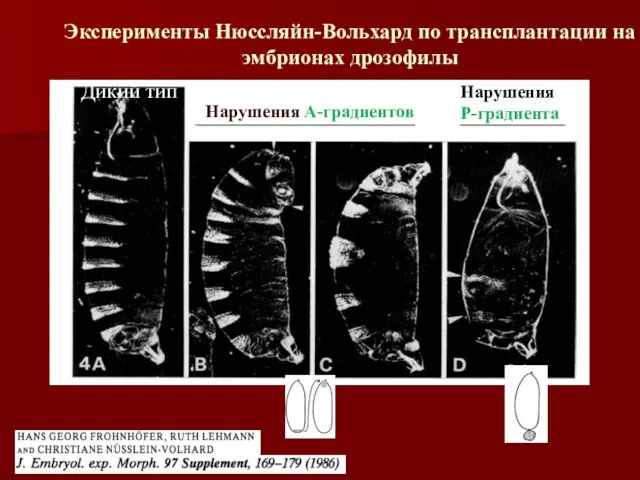
Дикий тип
Нарушения А-градиентов
Нарушения
Р-градиента
Эксперименты Нюссляйн-Вольхард по трансплантации на эмбрионах дрозофилы
Дикий тип
Нарушения А-градиентов
Нарушения
Р-градиента
Эксперименты Нюссляйн-Вольхард по трансплантации на эмбрионах дрозофилы
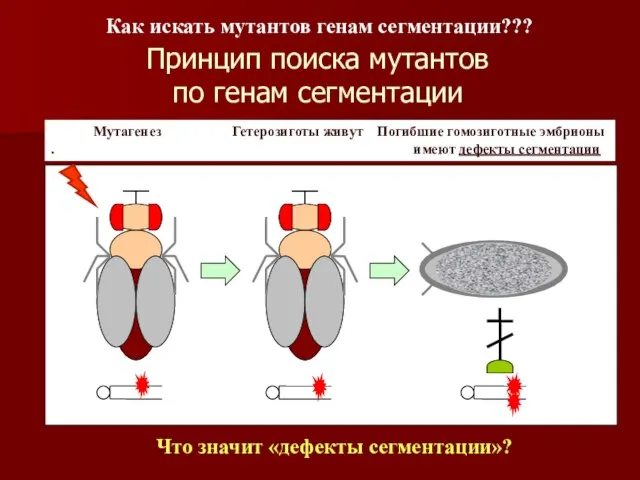
Принцип поиска мутантов
по генам сегментации
Мутагенез Гетерозиготы живут Погибшие гомозиготные
Принцип поиска мутантов
по генам сегментации
Мутагенез Гетерозиготы живут Погибшие гомозиготные
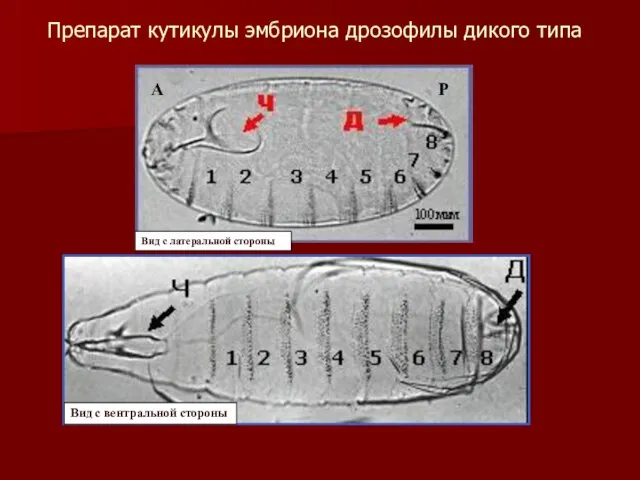
Препарат кутикулы эмбриона дрозофилы дикого типа
А
Р
Вид с латеральной стороны
Вид с вентральной
Препарат кутикулы эмбриона дрозофилы дикого типа
А
Р
Вид с латеральной стороны
Вид с вентральной
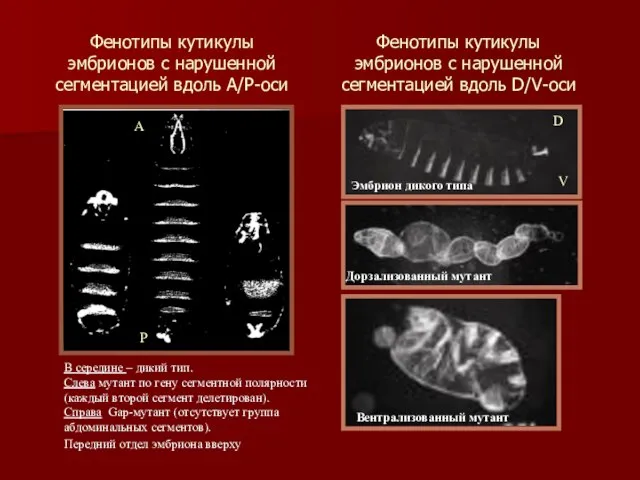
Фенотипы кутикулы эмбрионов с нарушенной сегментацией вдоль А/Р-оси
В середине – дикий
Фенотипы кутикулы эмбрионов с нарушенной сегментацией вдоль А/Р-оси
В середине – дикий

Каким образом формируются градиенты морфогенов в яйце?
Как поляризуется сам ооцит?
Каким образом формируются градиенты морфогенов в яйце?
Как поляризуется сам ооцит?
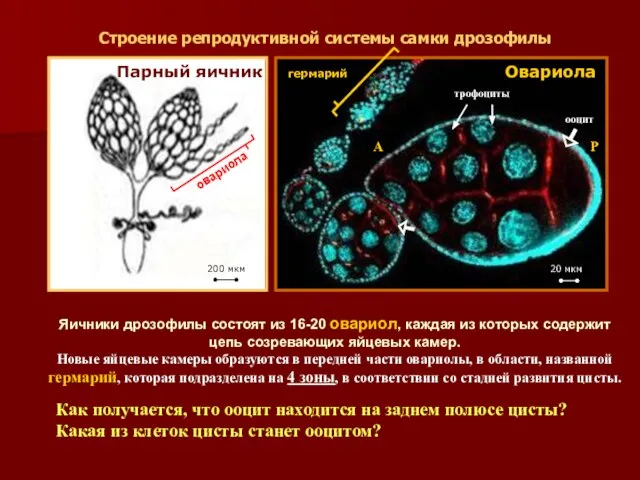
Строение репродуктивной системы самки дрозофилы
Парный яичник
Овариола
гермарий
овариола
трофоциты
ооцит
200 мкм
20 мкм
Яичники дрозофилы состоят из
Строение репродуктивной системы самки дрозофилы
Парный яичник
Овариола
гермарий
овариола
трофоциты
ооцит
200 мкм
20 мкм
Яичники дрозофилы состоят из
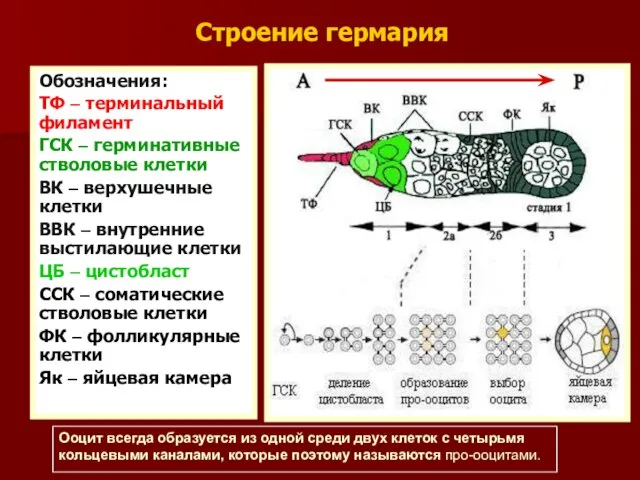
Строение гермария
Обозначения:
ТФ – терминальный филамент
ГСК – герминативные стволовые клетки
ВК –
Строение гермария
Обозначения:
ТФ – терминальный филамент
ГСК – герминативные стволовые клетки
ВК –
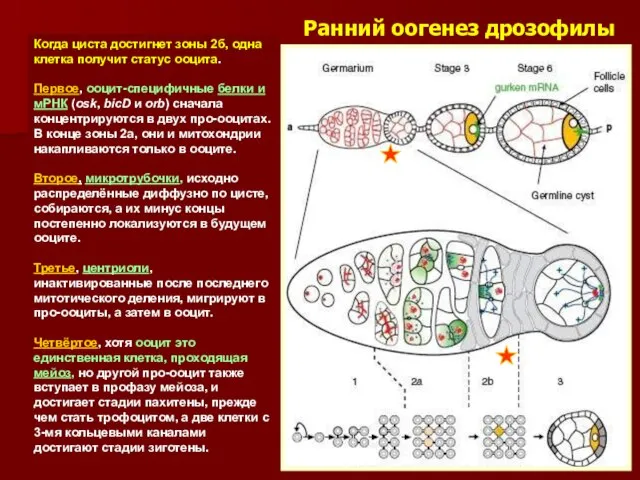
Ранний оогенез дрозофилы
Когда циста достигнет зоны 2б, одна клетка получит статус
Ранний оогенез дрозофилы
Когда циста достигнет зоны 2б, одна клетка получит статус
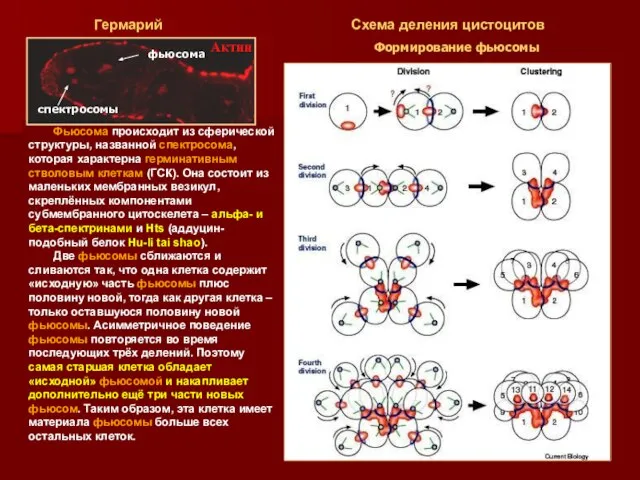
Фьюсома происходит из сферической структуры, названной спектросома, которая характерна герминативным стволовым
Фьюсома происходит из сферической структуры, названной спектросома, которая характерна герминативным стволовым
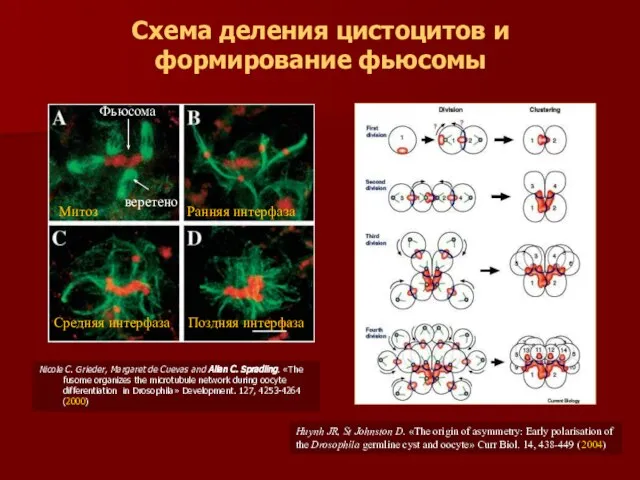
Nicole C. Grieder, Margaret de Cuevas and Allan C. Spradling. «The
Nicole C. Grieder, Margaret de Cuevas and Allan C. Spradling. «The

Следствие полярности фьюсомы
Циста в зоне 2а дифференцируется в двух направлениях:
Следствие полярности фьюсомы
Циста в зоне 2а дифференцируется в двух направлениях:
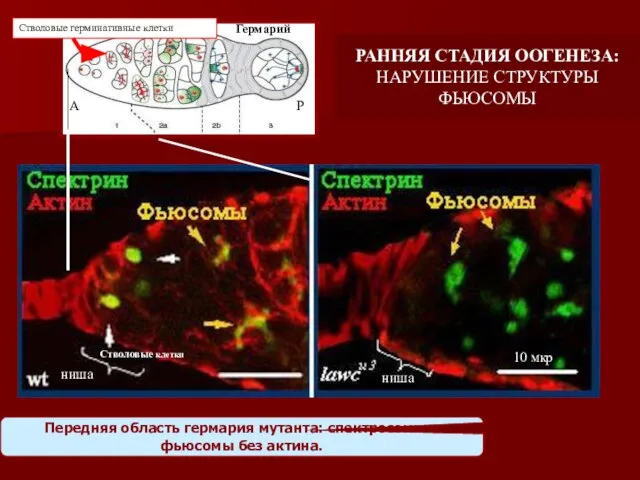
Передняя область гермария мутанта: спектросомы и фьюсомы без актина.
ниша
Стволовые клетки
Гермарий
Стволовые герминативные
Передняя область гермария мутанта: спектросомы и фьюсомы без актина.
ниша
Стволовые клетки
Гермарий
Стволовые герминативные
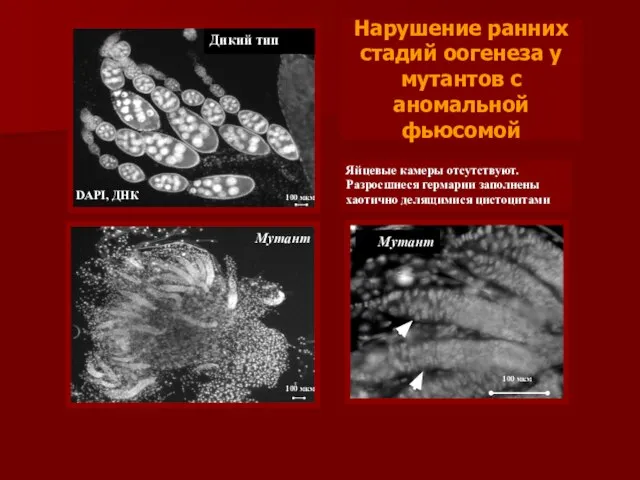
Нарушение ранних стадий оогенеза у мутантов с аномальной фьюсомой
Мутант
Яйцевые камеры
Нарушение ранних стадий оогенеза у мутантов с аномальной фьюсомой
Мутант
Яйцевые камеры

Установление A/P полярности ооцита
Когда герминативная циста достигает зоны 2б, транспортируемые
Установление A/P полярности ооцита
Когда герминативная циста достигает зоны 2б, транспортируемые
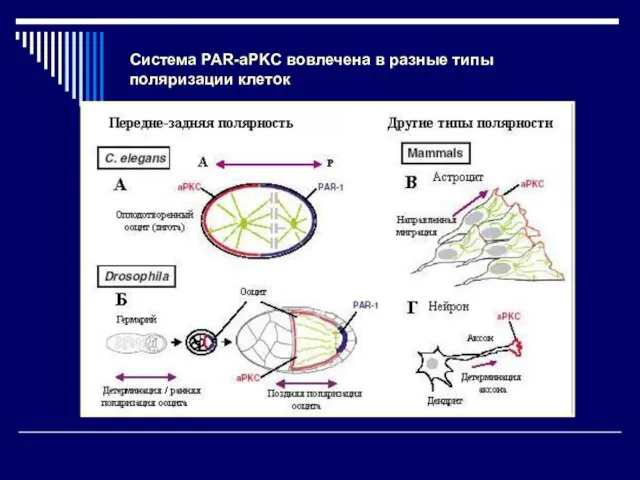
Система PAR-aPKC вовлечена в разные типы
поляризации клеток
Система PAR-aPKC вовлечена в разные типы
поляризации клеток

Current Biology, Vol. 14, 520–525, March 23, 2004,
Асимметричная локализация двух белков
Current Biology, Vol. 14, 520–525, March 23, 2004,
Асимметричная локализация двух белков

Роль кадгерина в формировании А/Р полярных осей
Локализация ооцита в заднем отделе
Роль кадгерина в формировании А/Р полярных осей
Локализация ооцита в заднем отделе
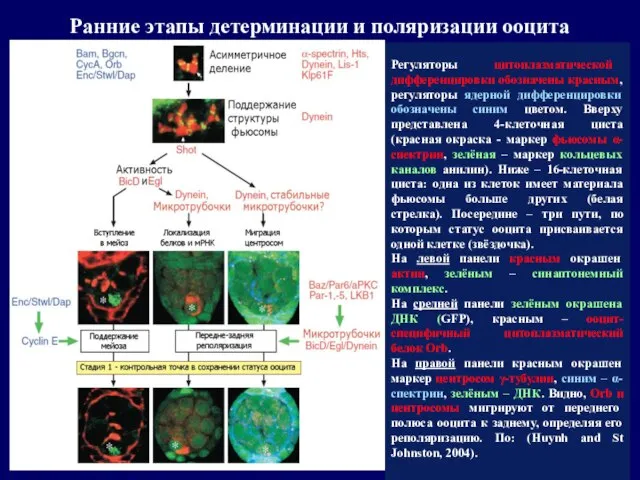
Регуляторы цитоплазматической дифференцировки обозначены красным, регуляторы ядерной дифференцировки обозначены синим цветом.
Регуляторы цитоплазматической дифференцировки обозначены красным, регуляторы ядерной дифференцировки обозначены синим цветом.

Фьюсома устанавливает оси полярности очень рано, в зоне 1 гермария
Затем в
Фьюсома устанавливает оси полярности очень рано, в зоне 1 гермария
Затем в
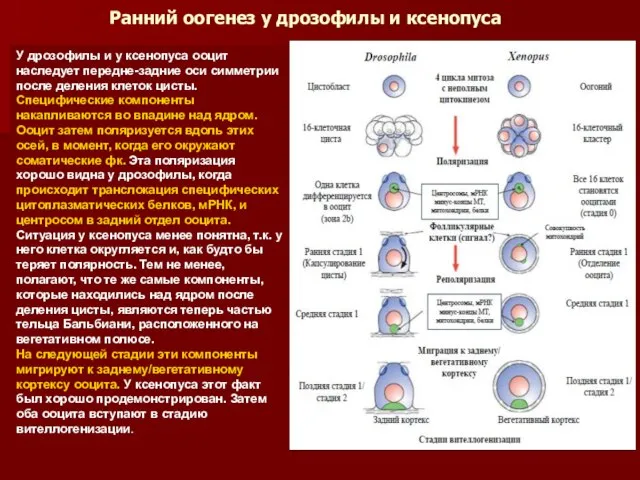
Ранний оогенез у дрозофилы и ксенопуса
У дрозофилы и у ксенопуса
Ранний оогенез у дрозофилы и ксенопуса
У дрозофилы и у ксенопуса

Я - тоже муха:
Мой краток век.
А чем ты, муха,
Не человек?
Я - тоже муха:
Мой краток век.
А чем ты, муха,
Не человек?

1995
Нобелевская премия по физиологии и медицине
«За открытия, касающиеся генетического контроля на
1995
Нобелевская премия по физиологии и медицине
«За открытия, касающиеся генетического контроля на
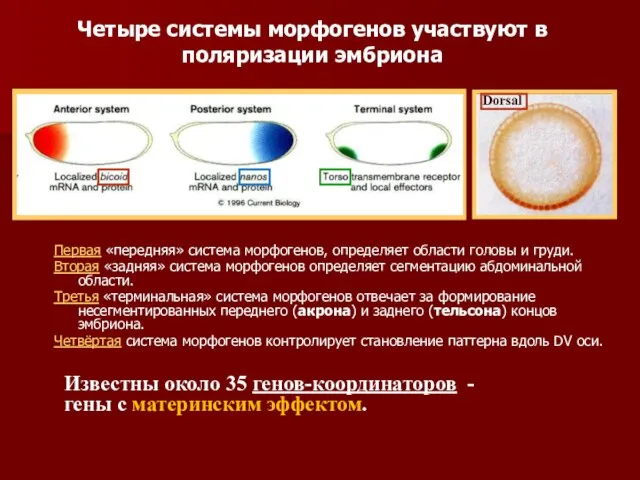
Первая «передняя» система морфогенов, определяет области головы и груди.
Вторая «задняя»
Вторая «задняя»
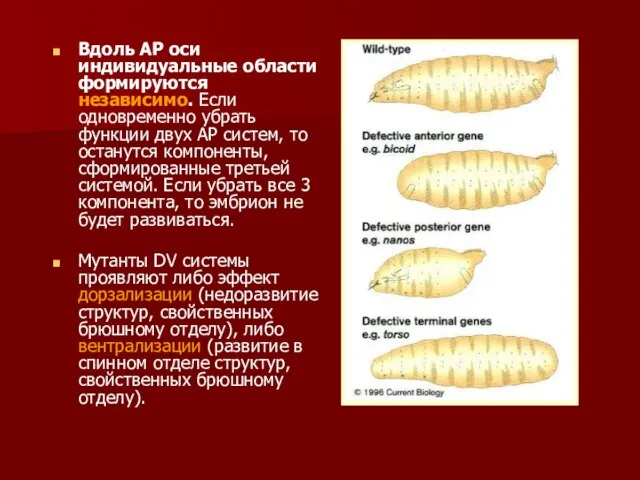
Вдоль АР оси индивидуальные области формируются независимо. Если одновременно убрать функции
Вдоль АР оси индивидуальные области формируются независимо. Если одновременно убрать функции

Общие свойства четырёх систем
Продукт одного гена каждой системы локализован в специфической
Общие свойства четырёх систем
Продукт одного гена каждой системы локализован в специфической
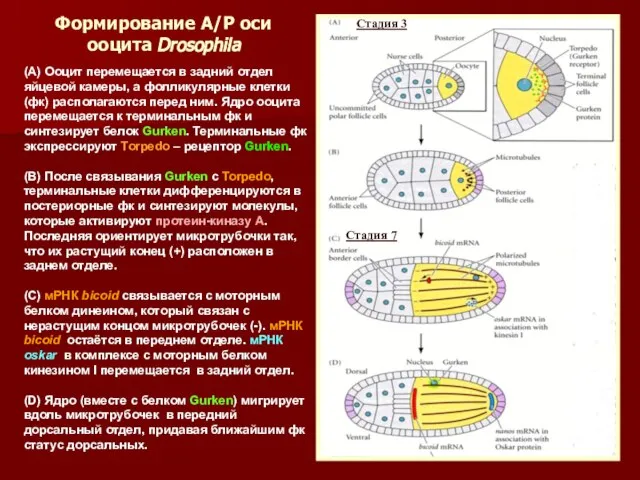
Формирование А/Р оси ооцита Drosophila
(А) Ооцит перемещается в задний отдел яйцевой
Формирование А/Р оси ооцита Drosophila
(А) Ооцит перемещается в задний отдел яйцевой
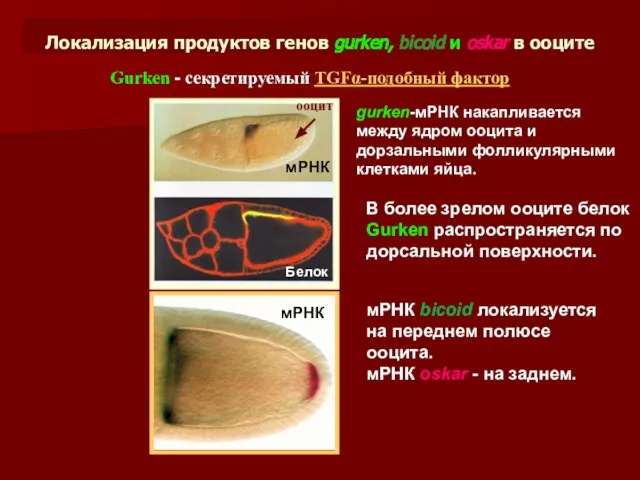
Локализация продуктов генов gurken, bicoid и oskar в ооците
мРНК
Белок
gurken-мРНК накапливается
Локализация продуктов генов gurken, bicoid и oskar в ооците
мРНК
Белок
gurken-мРНК накапливается

Антериорная система – морфогенетический градиент
А-система является самой простой. Только один ген
Антериорная система – морфогенетический градиент
А-система является самой простой. Только один ген
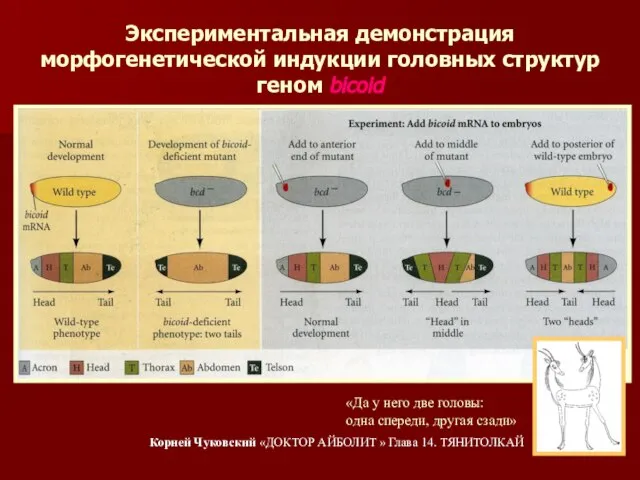
Экспериментальная демонстрация морфогенетической индукции головных структур геном bicoid
«Да у него две
Экспериментальная демонстрация морфогенетической индукции головных структур геном bicoid
«Да у него две

Формирование градиента в заднем отделе эмбриона
Центральный компонент Р-системы – продукт гена
Формирование градиента в заднем отделе эмбриона
Центральный компонент Р-системы – продукт гена
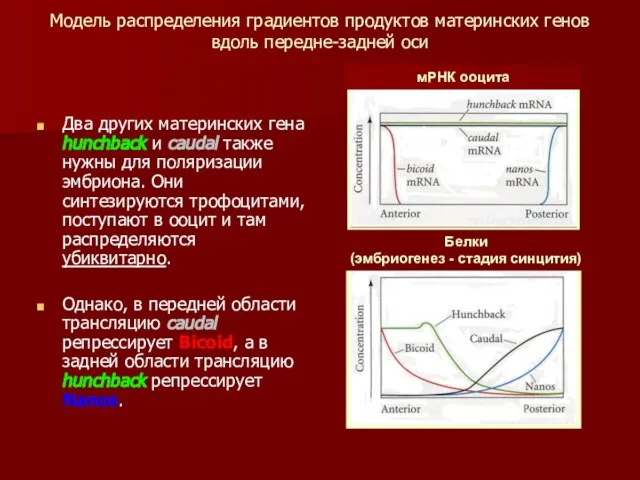
Модель распределения градиентов продуктов материнских генов вдоль передне-задней оси
Два других материнских
Модель распределения градиентов продуктов материнских генов вдоль передне-задней оси
Два других материнских
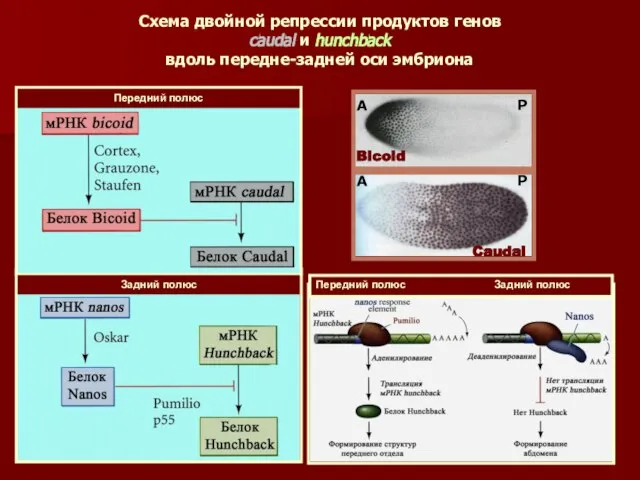
Схема двойной репрессии продуктов генов
caudal и hunchback
вдоль передне-задней оси
Схема двойной репрессии продуктов генов caudal и hunchback вдоль передне-задней оси
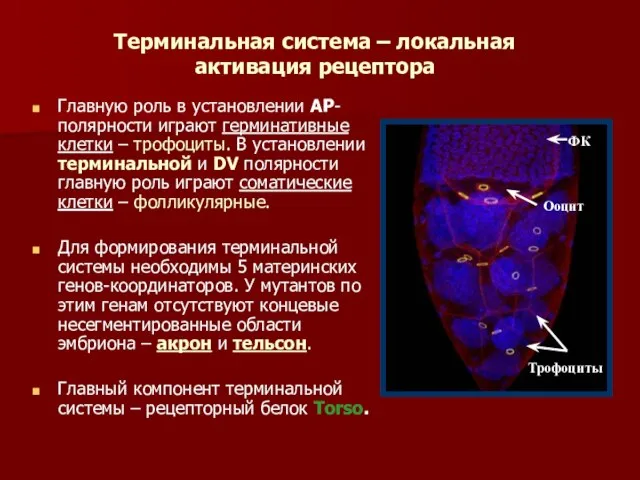
Терминальная система – локальная активация рецептора
Главную роль в установлении АР-полярности играют
Терминальная система – локальная активация рецептора
Главную роль в установлении АР-полярности играют
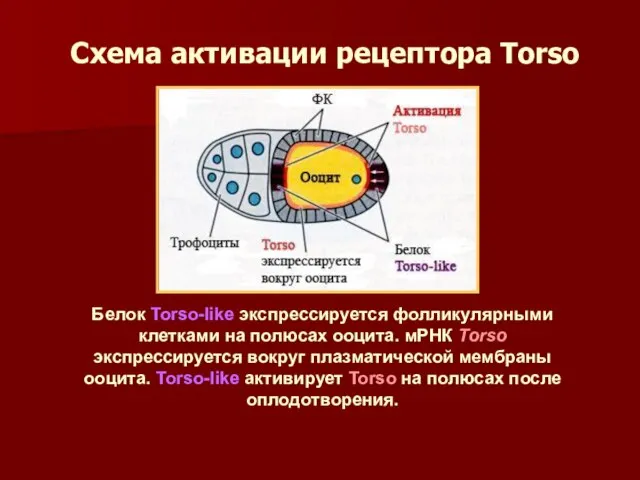
Схема активации рецептора Torso
Белок Torso-like экспрессируется фолликулярными клетками на полюсах ооцита.
Схема активации рецептора Torso
Белок Torso-like экспрессируется фолликулярными клетками на полюсах ооцита.

Активация рецептора Torso на полюсах яйца происходит в результате сигнальной трансдукции
Активация рецептора Torso на полюсах яйца происходит в результате сигнальной трансдукции
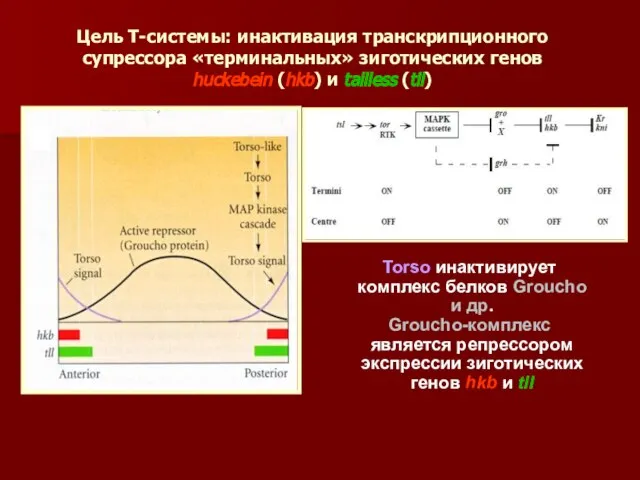
Цель Т-системы: инактивация транскрипционного супрессора «терминальных» зиготических генов
huckebein (hkb) и
Цель Т-системы: инактивация транскрипционного супрессора «терминальных» зиготических генов huckebein (hkb) и
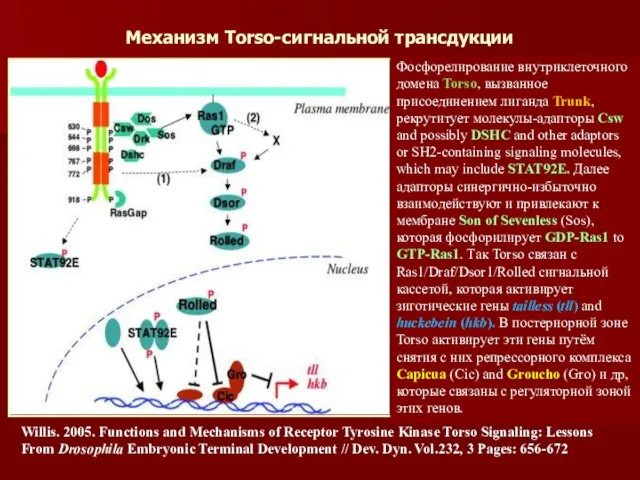
Механизм Torso-сигнальной трансдукции
Willis. 2005. Functions and Mechanisms of Receptor Tyrosine Kinase
Механизм Torso-сигнальной трансдукции
Willis. 2005. Functions and Mechanisms of Receptor Tyrosine Kinase
 Пространственная структура глобулярных белков
Пространственная структура глобулярных белков Непрямое постэмбриональное развитие
Непрямое постэмбриональное развитие Аминокислоты и белки Строение и свойства.
Аминокислоты и белки Строение и свойства. Клеточное строение организма
Клеточное строение организма Переходные формы в эволюции Составила учитель биологии Логунцова Ю.В. Школа № 14 и гимназия № 2 г.Кимры, Тверская обл.
Переходные формы в эволюции Составила учитель биологии Логунцова Ю.В. Школа № 14 и гимназия № 2 г.Кимры, Тверская обл.  ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова Оксана Викторовна
ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова Оксана Викторовна  Ұлулар
Ұлулар Жираф
Жираф Млекопитающие. Летучая мышь
Млекопитающие. Летучая мышь Тепе - теңдік талдағышы. Тактильді талдағышы. Ауырсыну
Тепе - теңдік талдағышы. Тактильді талдағышы. Ауырсыну Презентация на тему "«ПИЩЕВАРЕНИЕ»" - скачать бесплатно презентации по Биологии
Презентация на тему "«ПИЩЕВАРЕНИЕ»" - скачать бесплатно презентации по Биологии «Овсяное «молочко» Мука овсяная для детского и диабетического питания.
«Овсяное «молочко» Мука овсяная для детского и диабетического питания. Презентация на тему Зимующие птицы
Презентация на тему Зимующие птицы  Сущность жизни и свойства живого вещества
Сущность жизни и свойства живого вещества Чувство кворума и альтруизм у бактерий
Чувство кворума и альтруизм у бактерий Тема: Редкие растения
Тема: Редкие растения  Общие представления о жизни
Общие представления о жизни Презентация на тему "Губки 7 класс" - скачать презентации по Биологии
Презентация на тему "Губки 7 класс" - скачать презентации по Биологии Генетическая безопасность агробиоценозов Выполнила студентка 4 курса Факультета естествознания Селянкина Юлия
Генетическая безопасность агробиоценозов Выполнила студентка 4 курса Факультета естествознания Селянкина Юлия  Биоценозы озер
Биоценозы озер АТФ и другие органические соединения клетки
АТФ и другие органические соединения клетки Рептилії. Чому плаче крокодил?
Рептилії. Чому плаче крокодил? Imunitātes veidi
Imunitātes veidi Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии
Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии Анализаторы. Органы чувств
Анализаторы. Органы чувств ВОЗНИКНОВЕНИЕ АДАПТАЦИЙ И ИХ ОТНОСИТЕЛЬНЫЙ ХАРАКТЕР ВЫПОЛНИЛА УЧИТЕЛЬ БИОЛОГИИ МОУ «МИРНЕНСКАЯ СОШ» УЙСКОГО РАЙОНА ЧЕЛЯБИНСКОЙ
ВОЗНИКНОВЕНИЕ АДАПТАЦИЙ И ИХ ОТНОСИТЕЛЬНЫЙ ХАРАКТЕР ВЫПОЛНИЛА УЧИТЕЛЬ БИОЛОГИИ МОУ «МИРНЕНСКАЯ СОШ» УЙСКОГО РАЙОНА ЧЕЛЯБИНСКОЙ Птички-синички
Птички-синички Презентация на тему
Презентация на тему