- Генетика пола и сцепленных с полом признаков

Содержание
- 2. Хромосомная гипотеза наследственности Сразу же после переоткрытия законов Менделя ученые обратили внимание на факт удивительного сходства
- 3. Хромосомная теория наследственности Основные экспериментальные доказательства того, что гены находятся в хромосомах, были получены в исследованиях
- 4. В биологии понятие «Пол» принято определять как совокупность признаков и свойств организма, обеспечивающих функцию воспроизводства потомства
- 5. В онтогенезе пол особи может определяться: а) до оплодотворения яйцеклетки сперматозоидом (прогамное) определение пола; б) после
- 6. Прогамно До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток на быстро и медленно
- 7. Эпигамно У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза: если личинка садится на
- 8. Известно, что диплоидный набор хромосом возникает в зиготе в результате слияния двух гаплоидных гамет в процессе
- 9. Из этого следует принцип аллельного (парного) состояния гена у диплоидных организмов, который не соблюдается лишь для
- 10. Гетерогаметность пола достигается цитологической и генетической неравноценностью пары половых хромосом
- 11. Половая дискриминация? Нет – компенсация дозы активных генов! Чтобы сбалансировать действие генов Х хромосомы у мужчин
- 12. Типы хромосомного определения пола
- 13. Гапло-диплоидия Помимо этого у некоторых видов насекомых (пчелы, осы, муравьи) существует особый т.н. гапло-диплоидный тип определения
- 14. Функции половых хромосом Помимо основной своей задачи определения пола Х и У хромосомы выполняют и другие
- 15. «Гены находятся в хромосомах» Изучение механизма наследования признаков сцепленных с полом явилось экспериментальным доказательством хромосомной теории
- 16. Вернемся к опытам Т.Моргана, выполненным на дрозофиле В Х-хромосоме этого вида известно более 500 структурных генов.
- 17. В частности, Морган заметил, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих
- 18. Мутации глаз у Drozophila melanogaster Мутации окраски и формы глаз у плодовой мушки дрозофилы 1 -
- 19. Так, при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые
- 20. В реципрокном скрещивании красноглазого самца с белоглазой самкой в F1 наблюдалось расщепление 1:1, причем белыми глазами
- 21. Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т.Морган смог, только предположив, что ген, отвечающий за
- 22. Сцепленное с полом наследование при нерасхождении Х-хромосом Продолжая эксперименты по скрещиванию дрозофил, аспирант Т.Моргана Кальвин Бриджес
- 23. Сцепленное с полом наследование при нерасхождении Х-хромосом Р Хw Хw x Хw+ У
- 24. Балансовая теория определения пола у дрозофилы В 1921 году К. Бриджес обнаружил несколько фертильных самок, имевших
- 25. Балансовая теория определения пола у дрозофилы Р 3X + 3A х ХУ + 2А
- 26. К. Бриджес пришел к выводу, что пол определяется балансом числа половых хромосом и набором аутосом, а
- 27. Балансовая теория оказалась неприменимой к человеку и высшим растениям. У человека Y-хромосома играет другую роль. Известно,
- 28. Синдром Шерешевского-Тернера Особь X0 развивается главным образом по женскому типу, но имеет отличия от нормы, известные
- 29. Синдром Клайнфельтера Синдром Клайнфельтера встречается у одного из 400-600 новорождённых мальчиков и проявляется в недоразвитии первичных
- 30. Гинандроморфизм Еще одно доказательство того, что гены находятся в хромосомах, было получено на мухах дрозофилах, называемых
- 31. Гинандроморфизм В представленном на рисунке случае муха гетерозиготна по гену окраски глаз и по гену, влияющему
- 32. Признаки сцепленные с полом у человека К настоящему времени в Х-хромосоме у человека идентифицировано немногим более
- 33. Признаки сцепленные с полом у человека В зависимости от природы гена и его локализации можно выделить
- 34. Х-сцепленный рецессивный тип наследования Х-сцепленный рецессивный тип наследования характеризуется следующими признаками: 1) больные появляются не в
- 35. Дальтонизм частичная цветовая слепота, один из видов нарушения цветового зрения. Д. впервые описан в 1794 Дж.
- 36. Гемофилия А Гемофилия А - тяжелое заболевание, обусловленное дефектом фактора VIII свертывания крови. Встречается с частотой
- 38. Механизм наследования гемофилии
- 39. Х-сцепленный доминантный тип наследования Х-сцепленный доминантный тип наследования сходен с аутосомно-доминантным, за исключением того, что мужчина
- 41. Скачать презентацию

Хромосомная гипотеза наследственности
Сразу же после переоткрытия законов Менделя ученые обратили внимание
Хромосомная гипотеза наследственности
Сразу же после переоткрытия законов Менделя ученые обратили внимание

Хромосомная теория наследственности
Основные экспериментальные доказательства того, что гены находятся в хромосомах,
Хромосомная теория наследственности
Основные экспериментальные доказательства того, что гены находятся в хромосомах,

В биологии понятие «Пол» принято определять как совокупность признаков и свойств организма, обеспечивающих

В онтогенезе пол особи может определяться:
а) до оплодотворения яйцеклетки сперматозоидом (прогамное)
В онтогенезе пол особи может определяться:
а) до оплодотворения яйцеклетки сперматозоидом (прогамное)

Прогамно
До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток
Прогамно
До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток
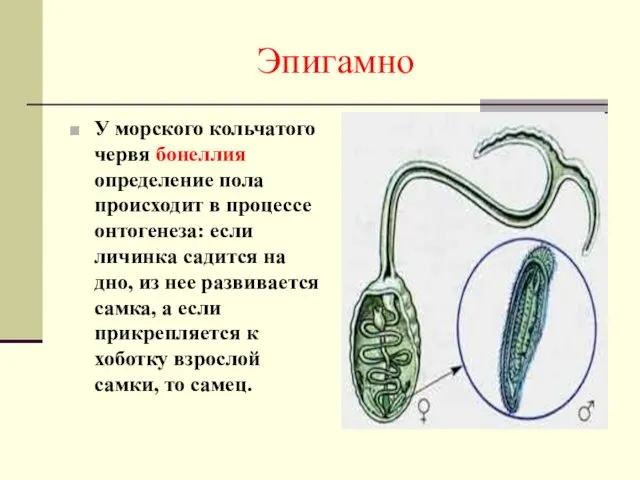
Эпигамно
У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза:
Эпигамно
У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза:

Известно, что диплоидный набор хромосом возникает в зиготе в результате слияния
Известно, что диплоидный набор хромосом возникает в зиготе в результате слияния

Из этого следует принцип аллельного (парного) состояния гена у диплоидных организмов,
Из этого следует принцип аллельного (парного) состояния гена у диплоидных организмов,

Гетерогаметность пола достигается цитологической и генетической неравноценностью пары половых хромосом
Гетерогаметность пола достигается цитологической и генетической неравноценностью пары половых хромосом

Половая дискриминация? Нет – компенсация дозы активных генов!
Чтобы сбалансировать действие генов
Половая дискриминация? Нет – компенсация дозы активных генов!
Чтобы сбалансировать действие генов

Типы хромосомного определения пола
Типы хромосомного определения пола

Гапло-диплоидия
Помимо этого у некоторых видов насекомых (пчелы, осы, муравьи) существует особый
Гапло-диплоидия
Помимо этого у некоторых видов насекомых (пчелы, осы, муравьи) существует особый

Функции половых хромосом
Помимо основной своей задачи определения пола Х и У
Функции половых хромосом
Помимо основной своей задачи определения пола Х и У

«Гены находятся в хромосомах»
Изучение механизма наследования признаков сцепленных с полом явилось
«Гены находятся в хромосомах»
Изучение механизма наследования признаков сцепленных с полом явилось

Вернемся к опытам Т.Моргана, выполненным на дрозофиле
В Х-хромосоме этого
Вернемся к опытам Т.Моргана, выполненным на дрозофиле
В Х-хромосоме этого

В частности, Морган заметил, что наследование окраски глаз у дрозофилы зависит
В частности, Морган заметил, что наследование окраски глаз у дрозофилы зависит

Мутации глаз у Drozophila melanogaster
Мутации окраски и формы глаз у плодовой
Мутации глаз у Drozophila melanogaster
Мутации окраски и формы глаз у плодовой

Так, при скрещивании белоглазого самца с красноглазой самкой в F1 были
Так, при скрещивании белоглазого самца с красноглазой самкой в F1 были

В реципрокном скрещивании красноглазого самца с белоглазой самкой в F1 наблюдалось
В реципрокном скрещивании красноглазого самца с белоглазой самкой в F1 наблюдалось

Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т.Морган смог, только
Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т.Морган смог, только

Сцепленное с полом наследование при нерасхождении Х-хромосом
Продолжая эксперименты по скрещиванию
Сцепленное с полом наследование при нерасхождении Х-хромосом
Продолжая эксперименты по скрещиванию

Сцепленное с полом наследование при нерасхождении Х-хромосом
Р Хw Хw x Хw+
Сцепленное с полом наследование при нерасхождении Х-хромосом
Р Хw Хw x Хw+

Балансовая теория определения пола у дрозофилы
В 1921 году К. Бриджес обнаружил
Балансовая теория определения пола у дрозофилы
В 1921 году К. Бриджес обнаружил

Балансовая теория определения пола у дрозофилы
Р 3X + 3A х
Балансовая теория определения пола у дрозофилы
Р 3X + 3A х

К. Бриджес пришел к выводу, что пол определяется балансом числа половых
К. Бриджес пришел к выводу, что пол определяется балансом числа половых

Балансовая теория оказалась неприменимой к человеку и высшим растениям. У человека
Балансовая теория оказалась неприменимой к человеку и высшим растениям. У человека
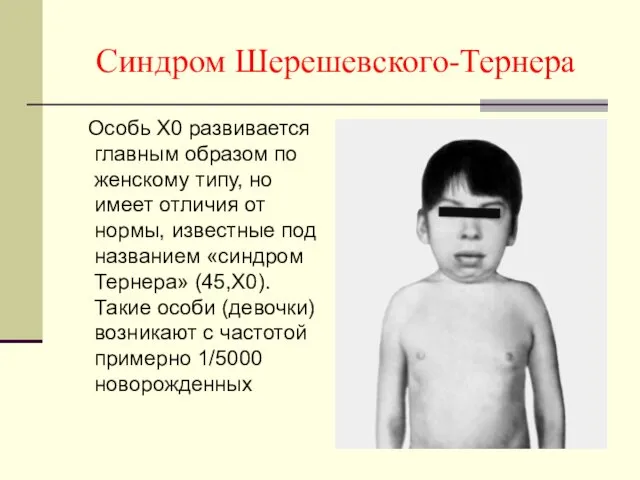
Синдром Шерешевского-Тернера
Особь X0 развивается главным образом по женскому типу, но
Синдром Шерешевского-Тернера
Особь X0 развивается главным образом по женскому типу, но

Синдром Клайнфельтера
Синдром Клайнфельтера встречается у одного из 400-600 новорождённых мальчиков
Синдром Клайнфельтера
Синдром Клайнфельтера встречается у одного из 400-600 новорождённых мальчиков

Гинандроморфизм
Еще одно доказательство того, что гены находятся в хромосомах, было получено
Гинандроморфизм
Еще одно доказательство того, что гены находятся в хромосомах, было получено

Гинандроморфизм
В представленном на рисунке случае муха гетерозиготна по гену окраски
Гинандроморфизм
В представленном на рисунке случае муха гетерозиготна по гену окраски

Признаки сцепленные с полом у человека
К настоящему времени в Х-хромосоме у
Признаки сцепленные с полом у человека
К настоящему времени в Х-хромосоме у

Признаки сцепленные с полом у человека
В зависимости от природы гена и
Признаки сцепленные с полом у человека
В зависимости от природы гена и
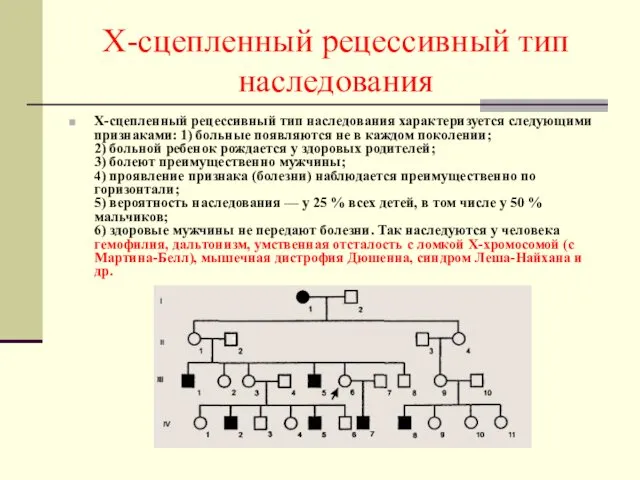
Х-сцепленный рецессивный тип наследования
Х-сцепленный рецессивный тип наследования характеризуется следующими признаками: 1)
Х-сцепленный рецессивный тип наследования
Х-сцепленный рецессивный тип наследования характеризуется следующими признаками: 1)

Дальтонизм
частичная цветовая слепота, один из видов нарушения цветового зрения.
Дальтонизм
частичная цветовая слепота, один из видов нарушения цветового зрения.

Гемофилия А
Гемофилия А - тяжелое заболевание, обусловленное дефектом фактора VIII
Гемофилия А
Гемофилия А - тяжелое заболевание, обусловленное дефектом фактора VIII
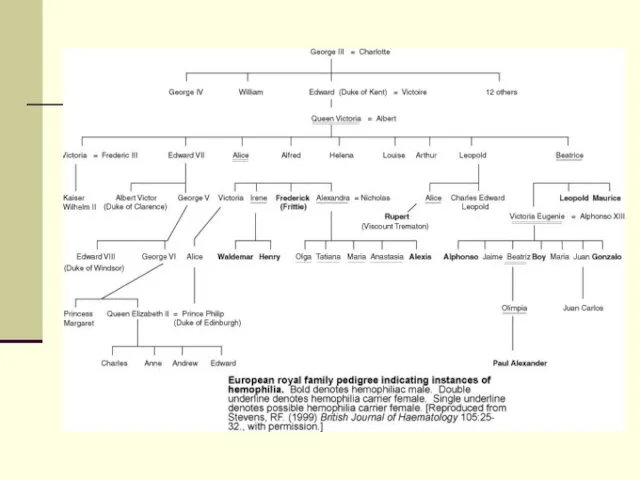

Механизм наследования гемофилии
Механизм наследования гемофилии

Х-сцепленный доминантный тип наследования
Х-сцепленный доминантный тип наследования сходен с аутосомно-доминантным,
Х-сцепленный доминантный тип наследования
Х-сцепленный доминантный тип наследования сходен с аутосомно-доминантным,
 Green algae
Green algae Тип Членистоногие
Тип Членистоногие Презентация на тему "Гемоглобин, его химический состав" - скачать презентации по Биологии
Презентация на тему "Гемоглобин, его химический состав" - скачать презентации по Биологии Презентация на тему Николай Иванович Пирогов
Презентация на тему Николай Иванович Пирогов  Птицеводство. 8 класс
Птицеводство. 8 класс Ветлужских Елена Викторовна учитель начальных классов муниципальное бюджетное общеобразовательное учреждение средняя общеобра
Ветлужских Елена Викторовна учитель начальных классов муниципальное бюджетное общеобразовательное учреждение средняя общеобра Хвостатая викторина
Хвостатая викторина Презентация на тему "Класс паукообразных" - скачать презентации по Биологии
Презентация на тему "Класс паукообразных" - скачать презентации по Биологии Embryonic development
Embryonic development Тема: Влияние температуры на живые организмы Цели: Дать понятие абиотическому фактору, тепловому балансу Проклассифицироват
Тема: Влияние температуры на живые организмы Цели: Дать понятие абиотическому фактору, тепловому балансу Проклассифицироват Индивидуальное развитие организма человека, или онтогенез
Индивидуальное развитие организма человека, или онтогенез Транспорт веществ в организме
Транспорт веществ в организме Баргузинский заповедник
Баргузинский заповедник Гомеостаз
Гомеостаз Уровни организации живой материи Презентация по биологии Подготовила учитель биологии Голубева С.В. МКОУ СОШ №4 г. Лесосибирск
Уровни организации живой материи Презентация по биологии Подготовила учитель биологии Голубева С.В. МКОУ СОШ №4 г. Лесосибирск Анато́мия челове́ка
Анато́мия челове́ка Мышцы предплечья, кисти
Мышцы предплечья, кисти В мире китов и дельфинов
В мире китов и дельфинов Царство бактерий Общая характеристика бактерий Урок по биологии учителя I квалификационной категории Халиуллиной Галии Кутдус
Царство бактерий Общая характеристика бактерий Урок по биологии учителя I квалификационной категории Халиуллиной Галии Кутдус Презентация на тему Гигиена питания
Презентация на тему Гигиена питания  Качества деревьев. Одиночные посадки
Качества деревьев. Одиночные посадки Основы генетики
Основы генетики Кожно-мышечное чувство, органы обоняния, вкуса. 8 класс
Кожно-мышечное чувство, органы обоняния, вкуса. 8 класс Общие вопросы. Задания 25, 26. Часть 2
Общие вопросы. Задания 25, 26. Часть 2 Презентация на тему "О влиянии курения и ПАВ на организм человека " - скачать презентации по Биологии
Презентация на тему "О влиянии курения и ПАВ на организм человека " - скачать презентации по Биологии Саморазвитие экосистем сукцессии
Саморазвитие экосистем сукцессии Зимующие птицы
Зимующие птицы Disease of Honeybees
Disease of Honeybees