- Групповой отбор vs индивидуальный отбор

Содержание
- 2. Изменение состава популяции (по признаку альтруистического поведения) под действием группового (А) и индивидуального (В) отбора Остались
- 3. Каким же образом распространяются признаки выгодные для группы и не выгодные для особи? Английский генетик и
- 4. Медведица (Salt-marsh moth) Estigmene acrea
- 5. Устойчивая эволюционная или эволюционно-стабильная стратегия (ЭСС) Что это такое? ЭСС – это такая стратегия, при которой
- 6. Ситуация 1. В популяции одни «голуби». Выигрыш +50–10= +40; проигрыш = –10. Средний итог равен +15.
- 7. Заметим, что это не групповой отбор, если бы все были бы «голубями», то выигрыш популяции (15
- 8. Из всех четырех стратегий только «отпорщик» устойчив. Почему? «Голуби» неустойчивы против «ястребов», «задир». Популяцию «ястребов» бы
- 13. Joop Jukema, Theunis Piersma Permanent female mimics in a lekking shorebird // Biol. Lett. 22 June
- 14. Альтруизм на примере общественных перепончатокрылых
- 15. Уильям Гамильтон 1964. Hamilton W.D. 1964. The genetical evolution of social behaviour // J. of Theor.
- 16. Итоговая (результирующая, совокупная ) приспособленность inclusive fitness Дельта ИП= (Na-na)ra+Σ(Nr-nr)rr Na – число детей альтруиста na–
- 17. Гаплодиплодия (самки –диплойдны, самцы – гаплойдны). Коэффициент родства с дочерью 1/2 , а с сестрой –
- 18. Реципрокный (взаимный) альтруизм Trivers R.L. 1971. The evolution of reciprocal altruism. // Quarterly Rev. of Biol.
- 19. Допустим у нас есть вид птиц, который поражен клещами. Они могут снять их отовсюду кроме головы.
- 20. Признаки реципрокного альтруизма Особи не родственны друг другу. Выигрыш существенно больше потери. Необходимо индивидуальное опознавание.
- 21. Родительский вклад Trivers R.L. 1972. Parental investment and sexual selection // Sexual Selection and the Descent
- 22. Вклад ограничен (число отложенных яиц, собранного корма) Должен ли он распределяться равномерно или неравномерно? Если условия
- 23. Могут ли потомки манипулировать вкладом? Trivers R.L. 1974. Parent – offspring conflict // American Zoologist 14
- 24. Существует ли конфликт между полами? Каждый пол заинтересован сэкономить свой вклад для того, чтобы вложить его
- 25. Гипотеза «Скверного переплета» Карлайл-Доукинса (Dawkins, Carlisle, 1976) объясняющая различие в преимущественной заботе о потомстве у животных
- 26. Таким образом, самка заинтересована найти самца, который будет вкладывать в потомство Самка «изучает» самцов. Один из
- 27. Генетический выигрыш за успешно выращенного детеныша +15 Расходы на одного детеныша – 20. Длительное ухаживание –
- 28. Если мы введем 4 стратегии: «скромница» не спаривается, пока самец не предпринял долгого ухаживания, «распутница» быстро
- 29. Ситуация 1. Все самки скромницы, все самцы верные супруги. Средний выигрыш 15 – 10 – 3=+2
- 30. Связь приспособленности и соотношения полов потомства Trivers R.L., Willard D.E., 1973. Natural selection of parental ability
- 31. 1975 Эдвард Уилсон, Гарвардский Университет
- 32. Стрелкой указан год выхода книги Уилсона (по Доукинсу) Стрелкой указан год выхода книги Уилсона (по Доукинсу)
- 33. Какие существуют возражения социобиологическому подходу 1. Обратный ход времени (будущий репродуктивный успех) 2. Противоречия с генетикой
- 34. Ответы на возражения Речь идет не об эволюции, а об устойчивости Работа Ротенбьюлера (Rothenbuhler, 1964) по
- 35. Возможен ли групповой отбор? Парадокс Симпсона John S. Chuang, Olivier Rivoire, Stanislas Leibler. Simpson’s Paradox in
- 36. Межгрупповая конкуренция способствует внутригрупповой кооперации «Внутригрупповые перетягивания каната (tag-of-war) вложены в межгрупповые». Члены группы соревнуются за
- 38. Скачать презентацию
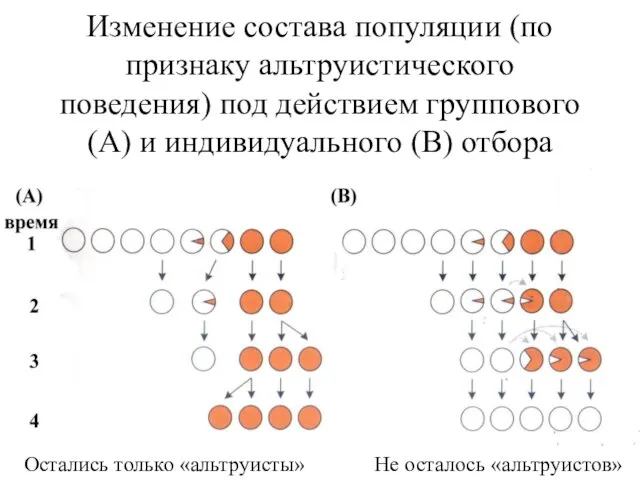
Изменение состава популяции (по признаку альтруистического поведения) под действием группового (А)
Изменение состава популяции (по признаку альтруистического поведения) под действием группового (А)

Каким же образом распространяются признаки выгодные для группы и не выгодные
Каким же образом распространяются признаки выгодные для группы и не выгодные

Медведица (Salt-marsh moth) Estigmene acrea
Медведица (Salt-marsh moth) Estigmene acrea

Устойчивая эволюционная или эволюционно-стабильная стратегия (ЭСС)
Что это такое? ЭСС –
Устойчивая эволюционная или эволюционно-стабильная стратегия (ЭСС)
Что это такое? ЭСС –

Ситуация 1. В популяции одни «голуби». Выигрыш +50–10= +40; проигрыш =
Ситуация 1. В популяции одни «голуби». Выигрыш +50–10= +40; проигрыш =

Заметим, что это не групповой отбор, если бы все были бы
Заметим, что это не групповой отбор, если бы все были бы

Из всех четырех стратегий только «отпорщик» устойчив. Почему?
«Голуби» неустойчивы против «ястребов»,
Из всех четырех стратегий только «отпорщик» устойчив. Почему?
«Голуби» неустойчивы против «ястребов»,





Joop Jukema, Theunis Piersma Permanent female mimics in a lekking shorebird
Joop Jukema, Theunis Piersma Permanent female mimics in a lekking shorebird

Альтруизм
на примере
общественных
перепончатокрылых
Альтруизм
на примере
общественных
перепончатокрылых

Уильям Гамильтон 1964.
Hamilton W.D. 1964. The genetical evolution of social
Уильям Гамильтон 1964.
Hamilton W.D. 1964. The genetical evolution of social

Итоговая (результирующая, совокупная ) приспособленность inclusive fitness
Дельта ИП= (Na-na)ra+Σ(Nr-nr)rr
Na –
Итоговая (результирующая, совокупная ) приспособленность inclusive fitness
Дельта ИП= (Na-na)ra+Σ(Nr-nr)rr
Na –

Гаплодиплодия (самки –диплойдны, самцы – гаплойдны). Коэффициент родства с дочерью 1/2
Гаплодиплодия (самки –диплойдны, самцы – гаплойдны). Коэффициент родства с дочерью 1/2

Реципрокный (взаимный) альтруизм
Trivers R.L. 1971. The evolution of reciprocal altruism. //
Реципрокный (взаимный) альтруизм
Trivers R.L. 1971. The evolution of reciprocal altruism. //

Допустим у нас есть вид птиц, который поражен клещами. Они могут
Допустим у нас есть вид птиц, который поражен клещами. Они могут

Признаки реципрокного альтруизма
Особи не родственны друг другу.
Выигрыш существенно больше потери.
Необходимо индивидуальное
Признаки реципрокного альтруизма
Особи не родственны друг другу.
Выигрыш существенно больше потери.
Необходимо индивидуальное

Родительский вклад
Trivers R.L. 1972. Parental investment and sexual selection // Sexual
Родительский вклад
Trivers R.L. 1972. Parental investment and sexual selection // Sexual

Вклад ограничен (число отложенных яиц, собранного корма)
Должен ли он распределяться равномерно
Вклад ограничен (число отложенных яиц, собранного корма)
Должен ли он распределяться равномерно

Могут ли потомки манипулировать вкладом?
Trivers R.L. 1974. Parent – offspring conflict
Могут ли потомки манипулировать вкладом?
Trivers R.L. 1974. Parent – offspring conflict

Существует ли конфликт между полами?
Каждый пол заинтересован сэкономить свой вклад
Существует ли конфликт между полами?
Каждый пол заинтересован сэкономить свой вклад
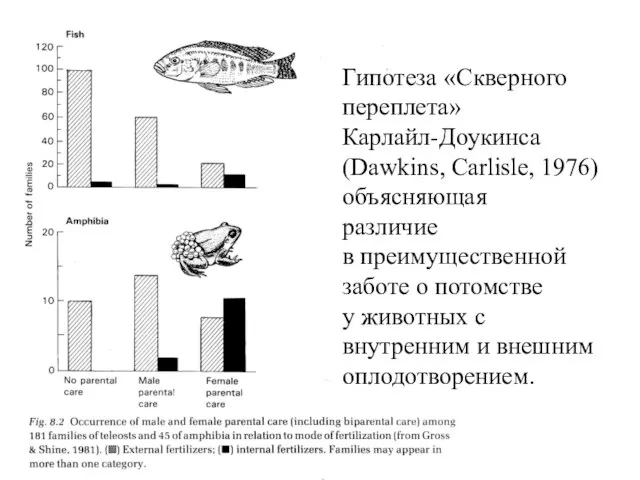
Гипотеза «Скверного
переплета»
Карлайл-Доукинса
(Dawkins, Carlisle, 1976)
объясняющая
различие
в преимущественной
заботе
Гипотеза «Скверного
переплета»
Карлайл-Доукинса
(Dawkins, Carlisle, 1976)
объясняющая
различие
в преимущественной
заботе

Таким образом, самка заинтересована найти самца, который будет вкладывать в потомство
Самка
Самка

Генетический выигрыш за успешно выращенного детеныша +15
Расходы на одного детеныша –
Генетический выигрыш за успешно выращенного детеныша +15
Расходы на одного детеныша –

Если мы введем 4 стратегии:
«скромница» не спаривается, пока самец не предпринял
Если мы введем 4 стратегии:
«скромница» не спаривается, пока самец не предпринял

Ситуация 1. Все самки скромницы, все самцы верные супруги. Средний выигрыш
Ситуация 1. Все самки скромницы, все самцы верные супруги. Средний выигрыш
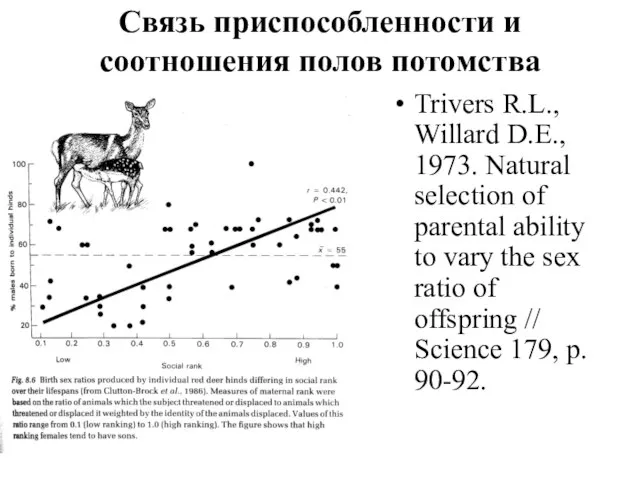
Связь приспособленности и соотношения полов потомства
Trivers R.L., Willard D.E., 1973. Natural
Связь приспособленности и соотношения полов потомства
Trivers R.L., Willard D.E., 1973. Natural

1975
Эдвард Уилсон,
Гарвардский Университет
1975
Эдвард Уилсон,
Гарвардский Университет
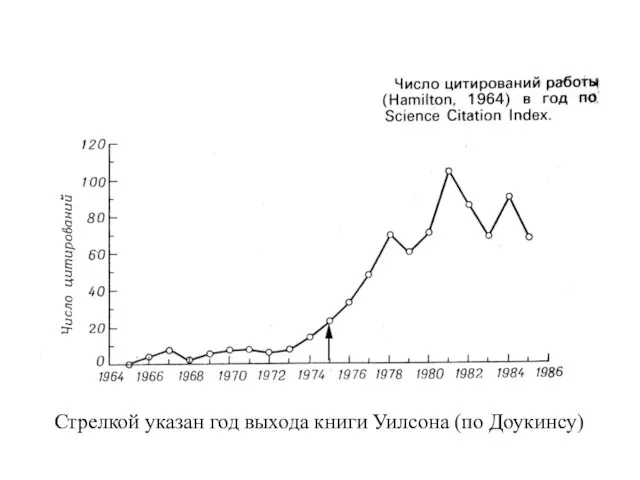
Стрелкой указан год выхода книги Уилсона (по Доукинсу)
Стрелкой указан год выхода
Стрелкой указан год выхода книги Уилсона (по Доукинсу)
Стрелкой указан год выхода

Какие существуют возражения социобиологическому подходу
1. Обратный ход времени (будущий репродуктивный успех)
2.
Какие существуют возражения социобиологическому подходу
1. Обратный ход времени (будущий репродуктивный успех)
2.

Ответы на возражения
Речь идет не об эволюции, а об устойчивости
Работа Ротенбьюлера
Ответы на возражения
Речь идет не об эволюции, а об устойчивости
Работа Ротенбьюлера
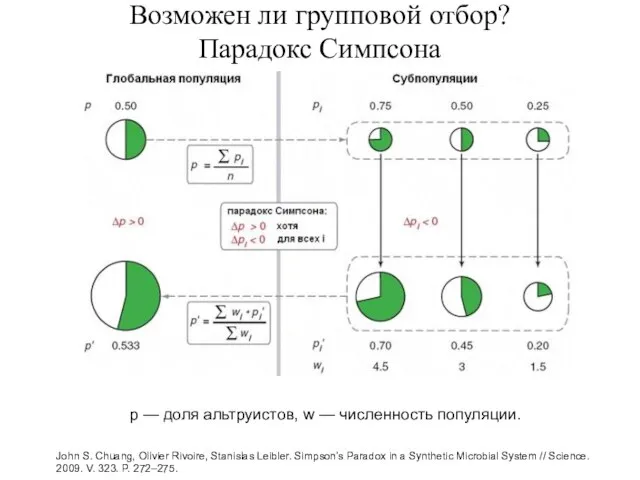
Возможен ли групповой отбор?
Парадокс Симпсона
John S. Chuang, Olivier Rivoire, Stanislas Leibler.
Возможен ли групповой отбор?
Парадокс Симпсона
John S. Chuang, Olivier Rivoire, Stanislas Leibler.
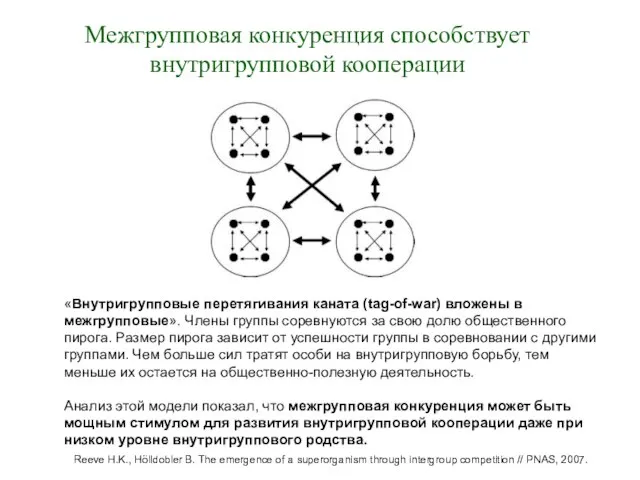
Межгрупповая конкуренция способствует внутригрупповой кооперации
«Внутригрупповые перетягивания каната (tag-of-war) вложены в межгрупповые».
Межгрупповая конкуренция способствует внутригрупповой кооперации
«Внутригрупповые перетягивания каната (tag-of-war) вложены в межгрупповые».
 Возникновение и проведение нервных импульсов в миелинизированных и немиелинизированных аксонах. 9 класс
Возникновение и проведение нервных импульсов в миелинизированных и немиелинизированных аксонах. 9 класс Головной мозг и его тайны!
Головной мозг и его тайны! Презентация на тему Виды и формы иммунитета
Презентация на тему Виды и формы иммунитета  Тип Саркодовые
Тип Саркодовые Взаимоотношения микроорганизмов и растений
Взаимоотношения микроорганизмов и растений Ферменты. Общие свойства ферментов
Ферменты. Общие свойства ферментов Спинной мозг
Спинной мозг Учитель биологии Антонова Н.Л.
Учитель биологии Антонова Н.Л. Основы учения об эволюции. Эволюционная теория Ч. Дарвина
Основы учения об эволюции. Эволюционная теория Ч. Дарвина Аспирин: за и против
Аспирин: за и против Наше питание (3 класс)
Наше питание (3 класс) Наблюдение за прорастанием семян фасоли
Наблюдение за прорастанием семян фасоли Медузы
Медузы Строение клеток, определение, свойства, функции
Строение клеток, определение, свойства, функции Декоративные качества растений
Декоративные качества растений Краниометрическая программа. Краниометрические размеры. (Тема 3)
Краниометрическая программа. Краниометрические размеры. (Тема 3) Углеводы содержатся в клетках растительных и животных организмов
Углеводы содержатся в клетках растительных и животных организмов Кровь Кровообращение
Кровь Кровообращение  Везикула. Везикулярный транспорт
Везикула. Везикулярный транспорт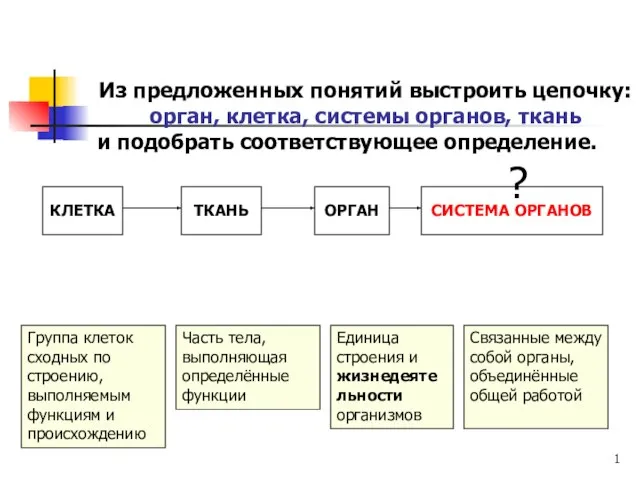 Из предложенных понятий выстроить цепочку: орган, клетка, системы органов, ткань и подобрать соответствующее определение
Из предложенных понятий выстроить цепочку: орган, клетка, системы органов, ткань и подобрать соответствующее определение Индивидуальное развитие растений
Индивидуальное развитие растений Фтор, роль в процессах жизнедеятельности
Фтор, роль в процессах жизнедеятельности Мир китов
Мир китов Презентация на тему "Орган вкуса" - скачать презентации по Биологии
Презентация на тему "Орган вкуса" - скачать презентации по Биологии Ферментативные свойства микроорганизмов
Ферментативные свойства микроорганизмов Презентация на тему "Магнолия" - скачать бесплатно презентации по Биологии
Презентация на тему "Магнолия" - скачать бесплатно презентации по Биологии Развитие жизни на Земле
Развитие жизни на Земле Охрана окружающей среды дома
Охрана окружающей среды дома