- Импринтинг. Нарушения импринтинга как причина наследственной патологии. Семинар 7

Содержание
- 2. Уровни эпигенетической регуляции Геномный Хромосомный Генный Эпигенетические нарушения Однородительская дисомия 2) Аномалии метилирования промоторных и регуляторных
- 3. Для нормального развития организма необходим равный вклад обоих родителей. 1. Трансплантация пронуклеусов. 2. Патология у человека.
- 4. Эпигенетические изменения на геномном уровне
- 5. Проявление эпигенетической патологии на хромосомном уровне – однородительская дисомия (ОРД) Однородительская дисомия, то есть наследование обеих
- 6. Однородительская дисомия (ОРД) Однородительская дисомия - наследование обеих копий целой хромосомы или ее части от одного
- 7. материнская ОРД по хромосоме 2 => признаки дисэмбриогенеза и отставание в развитии; отцовская ОРД по длинному
- 8. Эпигенотип (импринт) - совокупность модификаций, которые по-разному маркируют родительские аллели и обеспечивают моноаллельный характер экспрессии импринтированных
- 9. Установлено, что все известные импринтированные гены содержат области различного метилирования на двух родительских хромосомах, причем эти
- 10. Метилирование у млекопитающих Поддержание структуры хроматина и стабильности хромосом Инактивация повторов и интегрированной чужеродной ДНК Формирование
- 11. Характеристика CpG-островка • >200 пн, длина большинства -0.5-3 тпн. • Относительно высокий GC-состав (>50, обычно>60%), плотное
- 12. Почти все метилирования “стирается” в раннем эмбриогенезе за счет деметилирования и/или гидроксилирования метильных групп • Паттерн
- 13. • Во время эмбрионального развития, в первичных половых клетках проходит полногеномное деметилирование, которое стирает предыдущие родительские
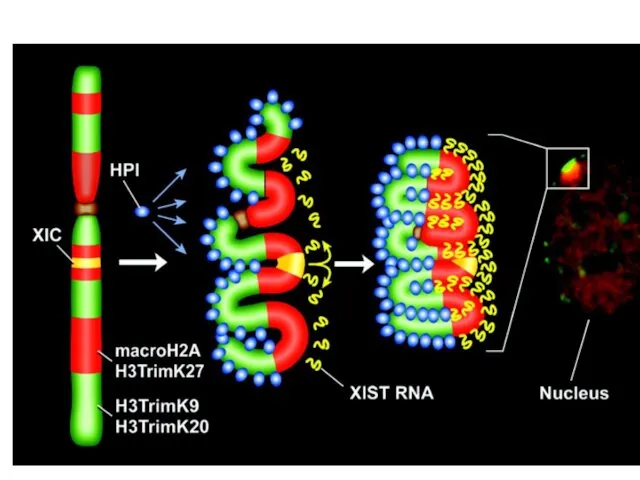
- 15. Инактивация X-хромосомы У млекопитающих компенсация дозы X-хромосом между женщинами (XX) и мужчинами (XY) достигается за счет
- 16. Инсуляторы — последовательности ДНК, особые регуляторные элементы, которые обладают способностью блокировать сигналы, исходящие от окружения. Эта
- 17. Во-вторых, инсулятор выполняет барьерную функцию для распространяющегося конденсированного хроматина. Существуют инсуляторы, выполняющие как одну из двух
- 18. Белок CTCF может блокировать энхансеры, препятствуя генной экспрессии. CTCF является первым обнаруженным белком, который необходим для
- 19. Модель X инактивация а) До инактивации Xist РНК экспрессируется в неустойчивом виде (пунктирные линии), что предполагает
- 20. Методы анализа метилирования 1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI) аналитическая чувствительность - 1: 2000
- 21. Метилчувствительная ПЦР Схема МЧ-ПЦР Анализ метилирования гена р16 методом МЧ-ПЦР в образцах ОЛ. Часть эндонуклеаз II
- 22. Метилспецифическая ПЦР
- 23. Бисульфитная модификация ДНК
- 24. Результаты метил-специфического секвенирования CpG-островка, расположенного в промотере РАК1 в культурах клеток HBL-100 и MCF-7. В результате
- 26. Бисульфитное секвенирование
- 27. Метильные ДНК-чипы
- 28. Метилирование ДНК вовлечено в широкий круг биологических процессов, которые включают регуляцию экспрессии тканеспецифичных генов, клеточную дифференцировку,
- 29. Районы локализации импринтированных генов. Розовым цветом отмечены гены, имеющие материнскую экспрессию, голубым – отцовскую, штриховкой –
- 30. ЭПИГЕНЕТИЧЕСКИЕ БОЛЕЗНИ ЧЕЛОВЕКА
- 31. Болезни импринтинга Синдромы Прадера-Вилли и Ангельмана – хромосома 15(q11.2-q13) Синдром Видеманна-Беквита - хромосома 11р15.5 Синдром Сильвера-Рассела
- 33. Ожирение, мышечная гипотония, низкий рост, гипогонадизм умственная отсталость различной степени выраженности признаки дизэмбриогенеза: долихоцефалия, гипертелоризм, эпикант,
- 34. Синдром Ангельмана (15q11-q13) - неврологические проявления: тяжелая задержка умственного и моторного развития, атаксия, гипотония, судорожная готовность,
- 35. Синдром Ангельмана
- 36. ОПРЕДЕЛЕНИЕ МИКРОДЕЛЕЦИЙ ХРОМОСОМЫ 15q11.2 ПРИ СИНДРОМАХ ПРАДЕРА-ВИЛЛИ И АНГЕЛЬМАНА МЕТОДОМ FISH (ДНК-зонд SNRPN).
- 37. Молекулярные причины СПВ и АС
- 39. Наследование мутаций центра импринтинга, приводящих к невозможности переключения импринта в герминальных клетках: (а) - СПВ: если
- 41. МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ РАЙОНА q11-q13 ХРОМОСОМЫ 15 ОТЦОВСКАЯ ХРОМОСОМА MN7 ZNF127 NDN IC SNRPN UBE3A MN7 МАТЕРИНСКАЯ
- 42. Схема функционирования центра импринтинга при синдроме Прадера-Вилли Объектом регуляции для ЦИ являются гены, которые в норме
- 44. Молекулярная диагностика СПВ и СА
- 45. Белок E6-AP, продукт гена UBE3A, входит в состав мультиферментного комплекса, обеспечивающего присоединение к цитоплазматическим белкам молекулы
- 46. Отклонение от менделевского наследования при нарушениях в ЦИ и мутациях в гене UBE3A
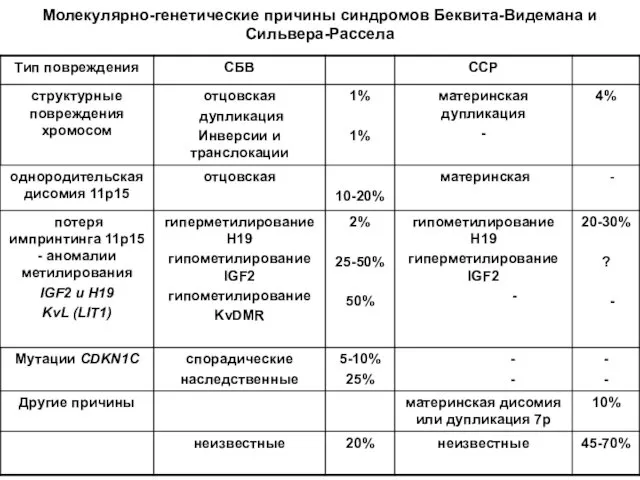
- 47. Синдром Видемана –Беквита Частота в популяции 1:10-12 тыс. Характерны пупочная грыжа, макроглоссия, гигантизм (средняя масса при
- 50. Гены H19 и IGF2 тесно сцеплены, противоположно импринтированы и их экспрессия регулируется общим энхансером, расположенным в
- 51. Гено-фенотипические корреляции при СВБ о о о о о о м м м м м о
- 52. Молекулярно-генетическая диагностика синдрома Видемана-Беквита 1 – негативный контроль; 2 – норма; 3,4,5 – пациенты с СВБ.
- 53. Пренатальная и постнатальная задержка роста; Треугольное лицо с выступающим лбом; Клинодактилия или брахидактилия; Макроцефалия; Скелетная асимметрия;
- 54. Рисунок 2а и 2б
- 55. Схема эпигенетической патологии при СРС (гипометилирование H19 на отцовской хромосоме 11)
- 56. Молекулярная диагностика синдромов Сильвера-Рассела и Беквита-Видемана (11р15) СРС БВС норма норма Н19 IGH2 Многолокусная ПЦР
- 57. 7p11.2-p13. У человека отцовская экспрессия GRB10 установлена в головном и спинном мозге, материнская – в скелетных
- 58. Молекулярно-генетические причины синдромов Беквита-Видемана и Сильвера-Рассела
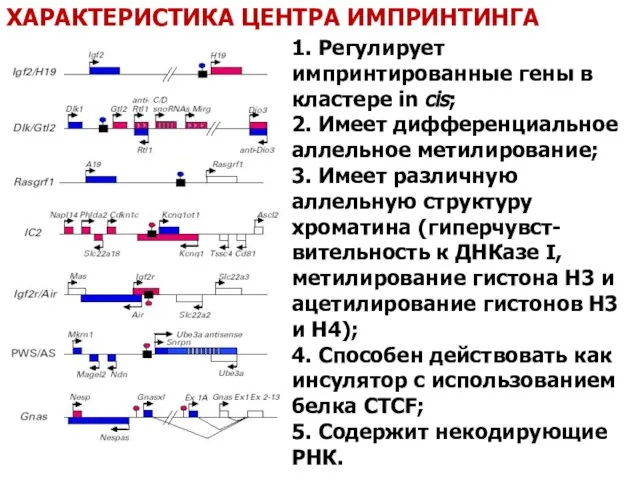
- 59. ХАРАКТЕРИСТИКА ЦЕНТРА ИМПРИНТИНГА 1. Регулирует импринтированные гены в кластере in cis; 2. Имеет дифференциальное аллельное метилирование;
- 62. Скачать презентацию

Уровни эпигенетической регуляции
Геномный
Хромосомный
Генный
Эпигенетические нарушения
Однородительская дисомия
2) Аномалии
Уровни эпигенетической регуляции
Геномный
Хромосомный
Генный
Эпигенетические нарушения
Однородительская дисомия
2) Аномалии

Для нормального развития организма необходим равный вклад обоих родителей.
1. Трансплантация
Для нормального развития организма необходим равный вклад обоих родителей.
1. Трансплантация
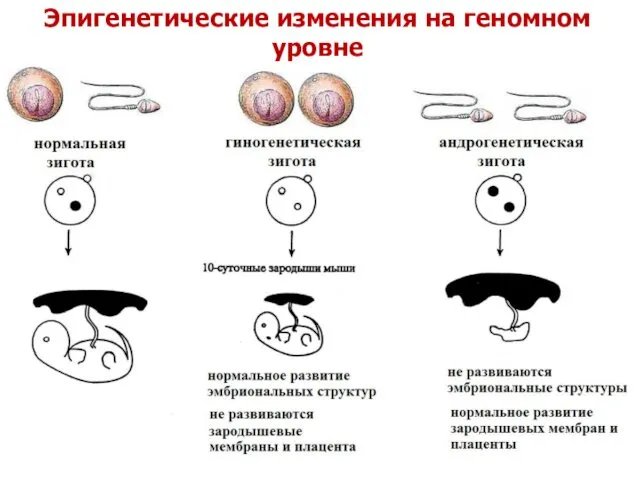
Эпигенетические изменения на геномном уровне
Эпигенетические изменения на геномном уровне

Проявление эпигенетической патологии
на хромосомном уровне – однородительская дисомия (ОРД)
Однородительская дисомия, то
Проявление эпигенетической патологии
на хромосомном уровне – однородительская дисомия (ОРД)
Однородительская дисомия, то
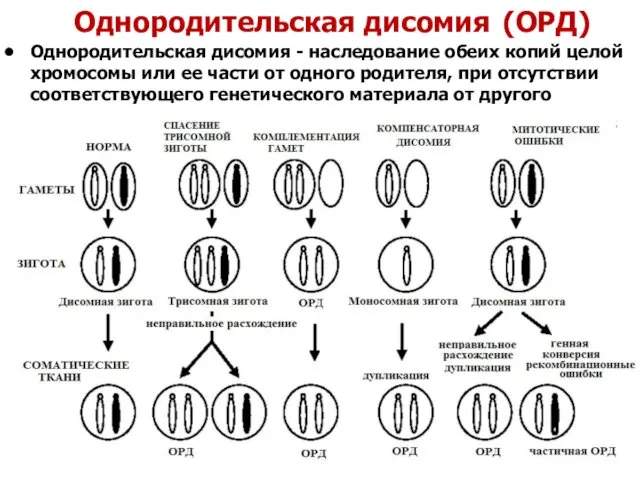
Однородительская дисомия (ОРД)
Однородительская дисомия - наследование обеих копий целой хромосомы
Однородительская дисомия (ОРД)
Однородительская дисомия - наследование обеих копий целой хромосомы

материнская ОРД по хромосоме 2 => признаки дисэмбриогенеза и отставание в
материнская ОРД по хромосоме 2 => признаки дисэмбриогенеза и отставание в

Эпигенотип (импринт) - совокупность модификаций, которые по-разному маркируют родительские аллели
Эпигенотип (импринт) - совокупность модификаций, которые по-разному маркируют родительские аллели

Установлено, что все известные импринтированные гены содержат области различного метилирования
Установлено, что все известные импринтированные гены содержат области различного метилирования
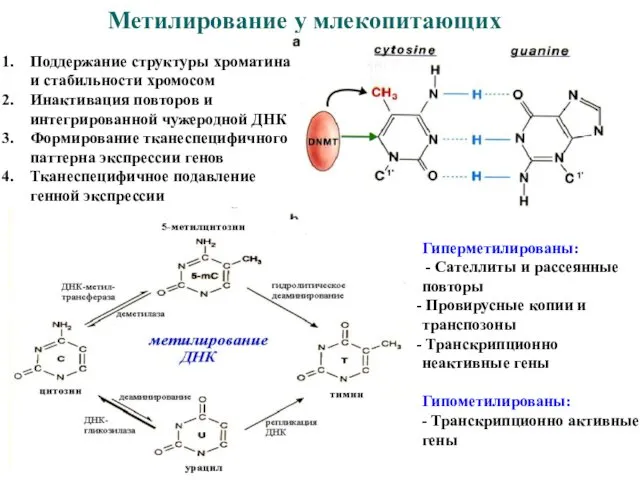
Метилирование у млекопитающих
Поддержание структуры хроматина и стабильности хромосом
Инактивация повторов и интегрированной
Метилирование у млекопитающих
Поддержание структуры хроматина и стабильности хромосом
Инактивация повторов и интегрированной

Характеристика CpG-островка
• >200 пн, длина большинства -0.5-3 тпн.
• Относительно высокий
Характеристика CpG-островка
• >200 пн, длина большинства -0.5-3 тпн.
• Относительно высокий

Почти все метилирования “стирается” в раннем эмбриогенезе за счет деметилирования и/или
Почти все метилирования “стирается” в раннем эмбриогенезе за счет деметилирования и/или
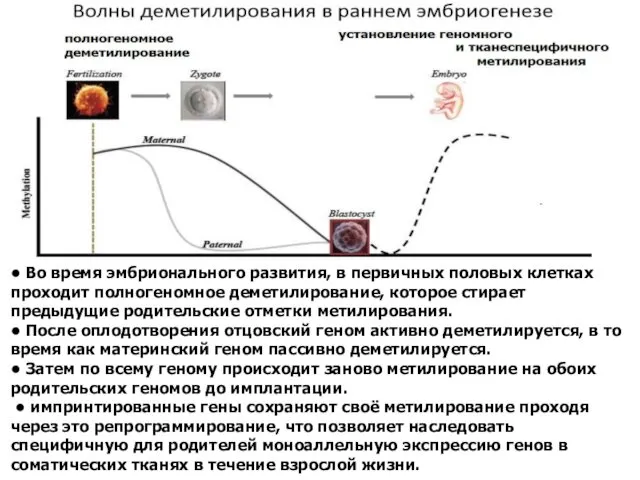
• Во время эмбрионального развития, в первичных половых клетках проходит полногеномное
• Во время эмбрионального развития, в первичных половых клетках проходит полногеномное
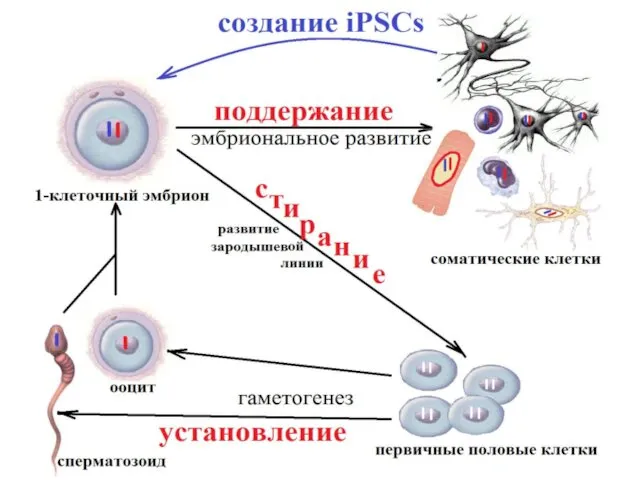

Инактивация X-хромосомы
У млекопитающих компенсация дозы X-хромосом между женщинами (XX) и мужчинами
Инактивация X-хромосомы
У млекопитающих компенсация дозы X-хромосом между женщинами (XX) и мужчинами
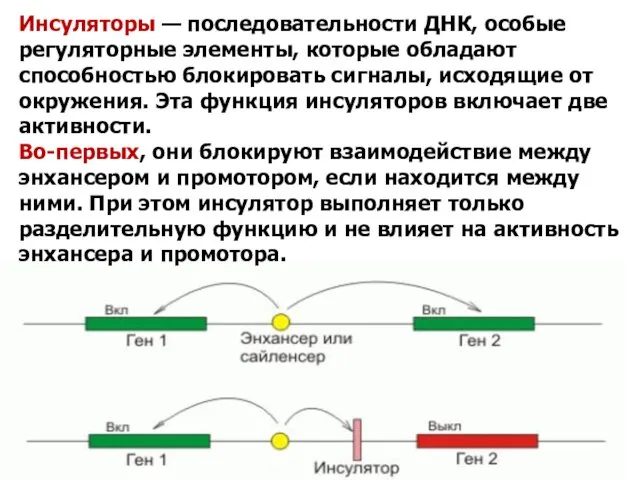
Инсуляторы — последовательности ДНК, особые регуляторные элементы, которые обладают способностью блокировать сигналы,
Инсуляторы — последовательности ДНК, особые регуляторные элементы, которые обладают способностью блокировать сигналы,
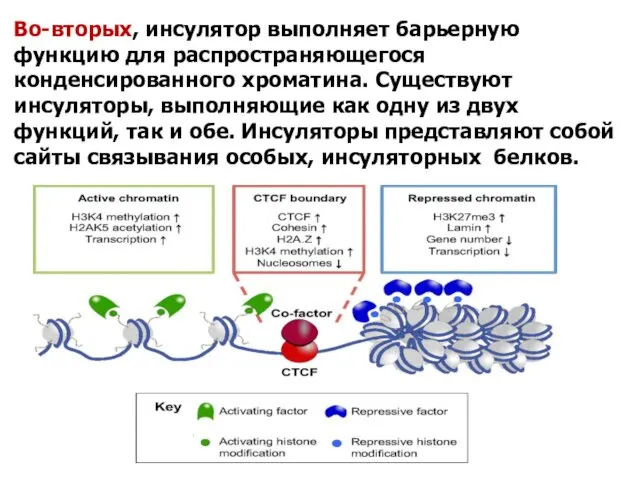
Во-вторых, инсулятор выполняет барьерную функцию для распространяющегося конденсированного хроматина. Существуют инсуляторы, выполняющие
Во-вторых, инсулятор выполняет барьерную функцию для распространяющегося конденсированного хроматина. Существуют инсуляторы, выполняющие
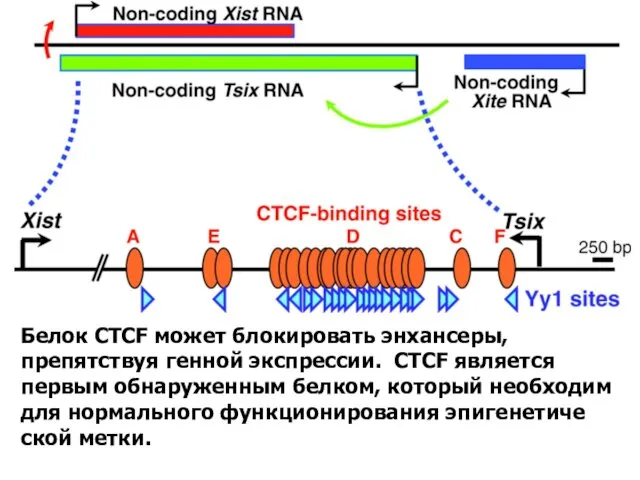
Белок CTCF может блокировать энхансеры, препятствуя генной экспрессии. CTCF является первым
Белок CTCF может блокировать энхансеры, препятствуя генной экспрессии. CTCF является первым
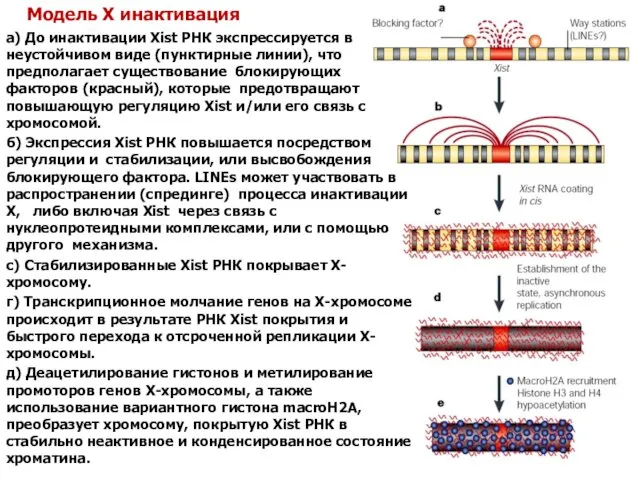
Модель X инактивация
а) До инактивации Xist РНК экспрессируется в неустойчивом виде
Модель X инактивация
а) До инактивации Xist РНК экспрессируется в неустойчивом виде

Методы анализа метилирования
1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI)
аналитическая
Методы анализа метилирования
1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI)
аналитическая
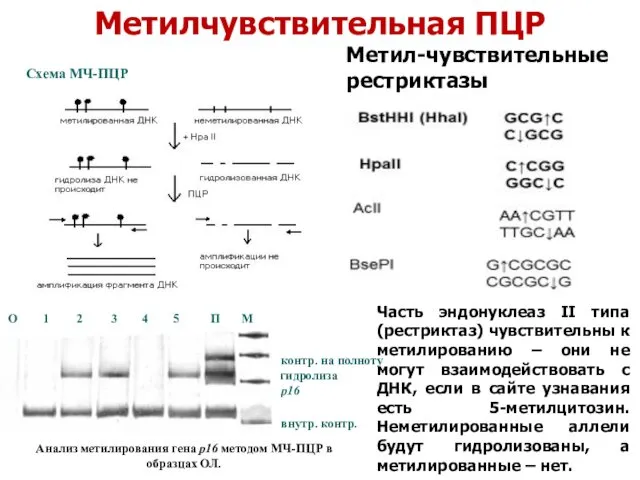
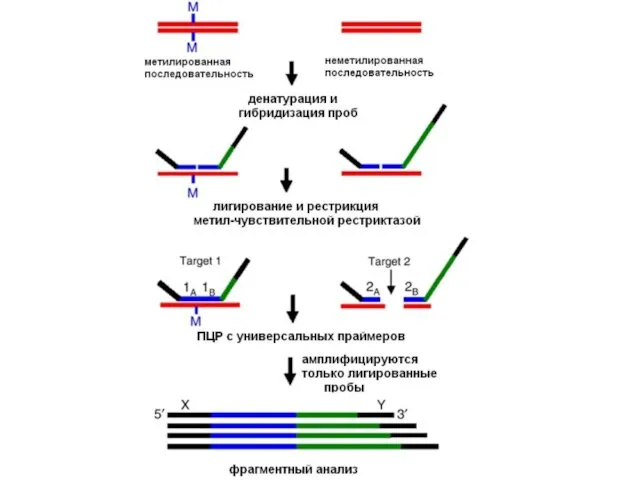
Метилчувствительная ПЦР
Схема МЧ-ПЦР
Анализ метилирования гена р16 методом МЧ-ПЦР в образцах ОЛ.
Часть
Метилчувствительная ПЦР
Схема МЧ-ПЦР
Анализ метилирования гена р16 методом МЧ-ПЦР в образцах ОЛ.
Часть
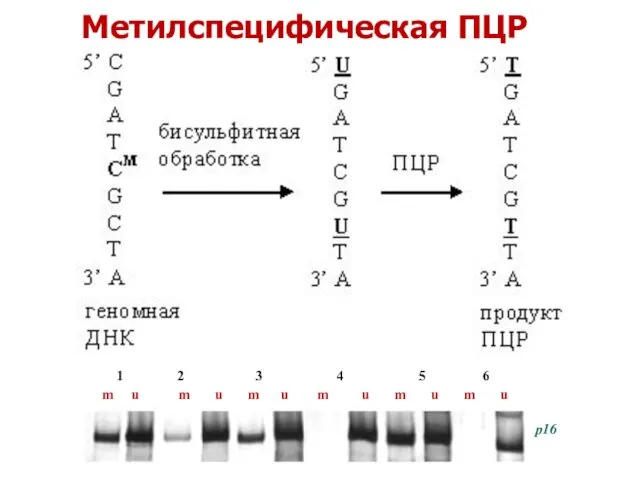
Метилспецифическая ПЦР
Метилспецифическая ПЦР
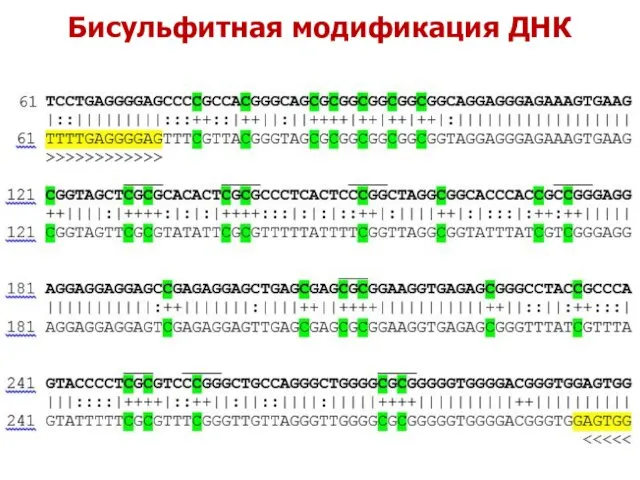
Бисульфитная модификация ДНК
Бисульфитная модификация ДНК

Результаты метил-специфического секвенирования CpG-островка, расположенного в промотере РАК1 в культурах клеток
Результаты метил-специфического секвенирования CpG-островка, расположенного в промотере РАК1 в культурах клеток
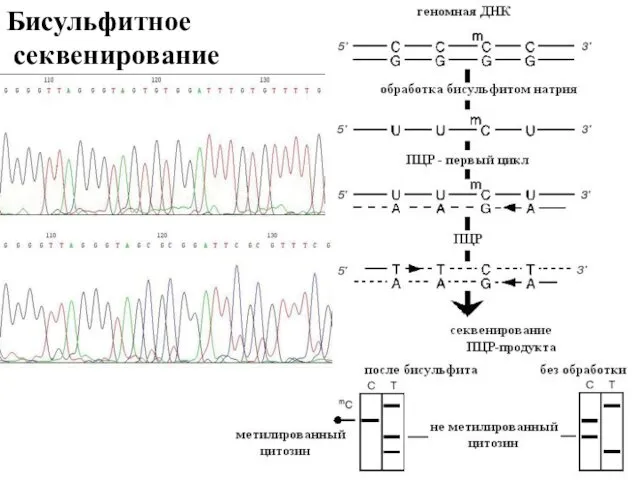
Бисульфитное
секвенирование
Бисульфитное
секвенирование
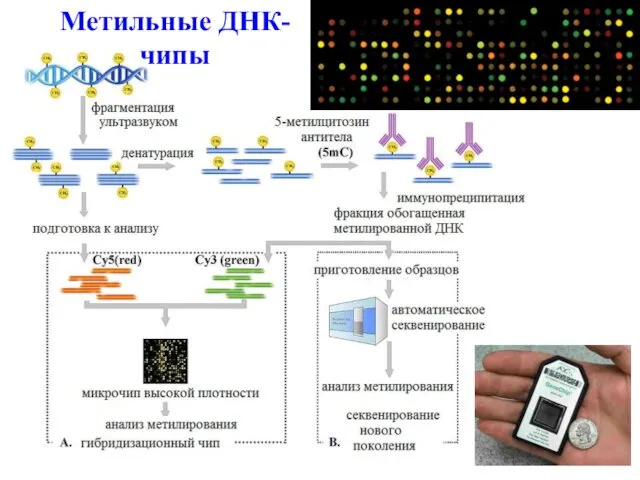
Метильные ДНК-чипы
Метильные ДНК-чипы

Метилирование ДНК вовлечено в широкий круг биологических процессов, которые включают регуляцию
Метилирование ДНК вовлечено в широкий круг биологических процессов, которые включают регуляцию
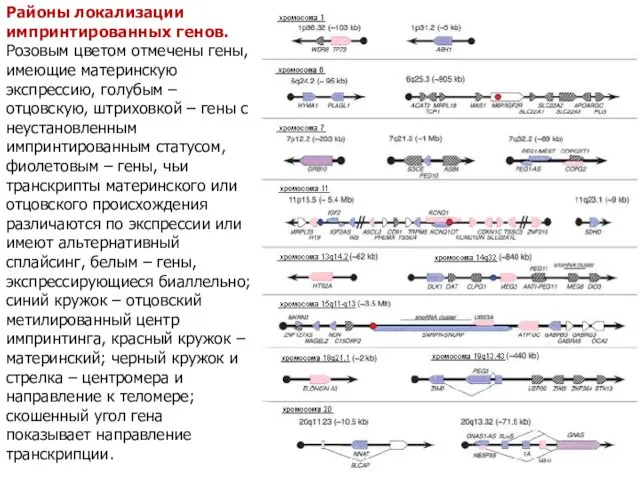
Районы локализации импринтированных генов. Розовым цветом отмечены гены, имеющие материнскую экспрессию,
Районы локализации импринтированных генов. Розовым цветом отмечены гены, имеющие материнскую экспрессию,

ЭПИГЕНЕТИЧЕСКИЕ БОЛЕЗНИ ЧЕЛОВЕКА
ЭПИГЕНЕТИЧЕСКИЕ БОЛЕЗНИ ЧЕЛОВЕКА

Болезни импринтинга
Синдромы Прадера-Вилли и Ангельмана –
хромосома 15(q11.2-q13)
Синдром Видеманна-Беквита - хромосома
Болезни импринтинга
Синдромы Прадера-Вилли и Ангельмана –
хромосома 15(q11.2-q13)
Синдром Видеманна-Беквита - хромосома

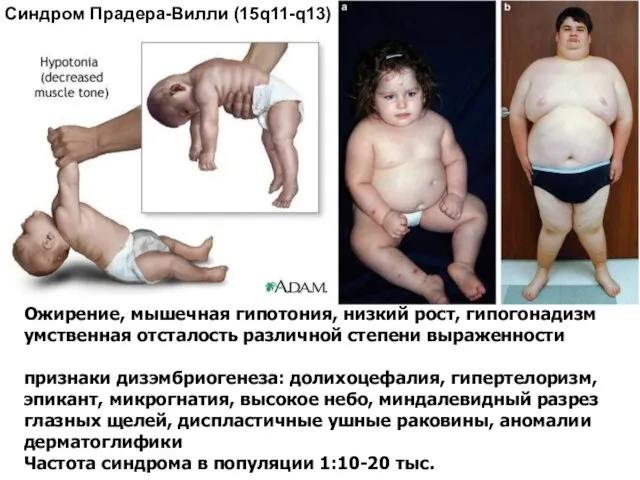
Ожирение, мышечная гипотония, низкий рост, гипогонадизм
умственная отсталость различной степени выраженности
признаки дизэмбриогенеза:
Ожирение, мышечная гипотония, низкий рост, гипогонадизм
умственная отсталость различной степени выраженности
признаки дизэмбриогенеза:

Синдром Ангельмана (15q11-q13)
- неврологические проявления: тяжелая задержка умственного и моторного развития,
Синдром Ангельмана (15q11-q13)
- неврологические проявления: тяжелая задержка умственного и моторного развития,
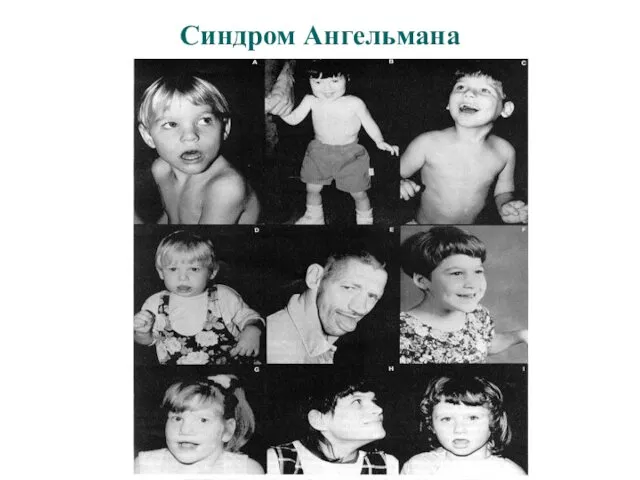
Синдром Ангельмана
Синдром Ангельмана
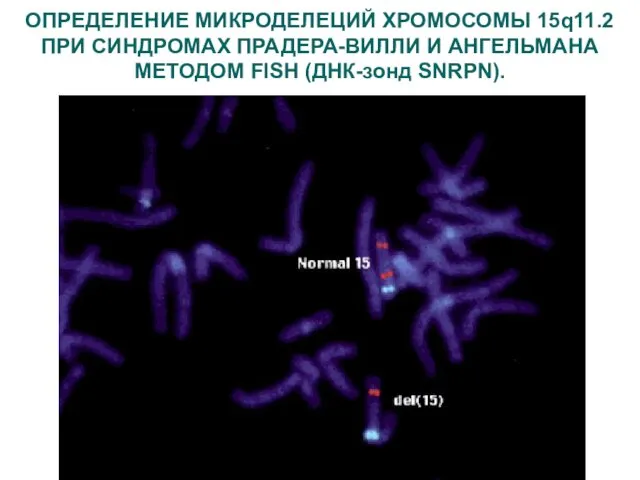
ОПРЕДЕЛЕНИЕ МИКРОДЕЛЕЦИЙ ХРОМОСОМЫ 15q11.2 ПРИ СИНДРОМАХ ПРАДЕРА-ВИЛЛИ И АНГЕЛЬМАНА МЕТОДОМ FISH
ОПРЕДЕЛЕНИЕ МИКРОДЕЛЕЦИЙ ХРОМОСОМЫ 15q11.2 ПРИ СИНДРОМАХ ПРАДЕРА-ВИЛЛИ И АНГЕЛЬМАНА МЕТОДОМ FISH

Молекулярные причины СПВ и АС
Молекулярные причины СПВ и АС
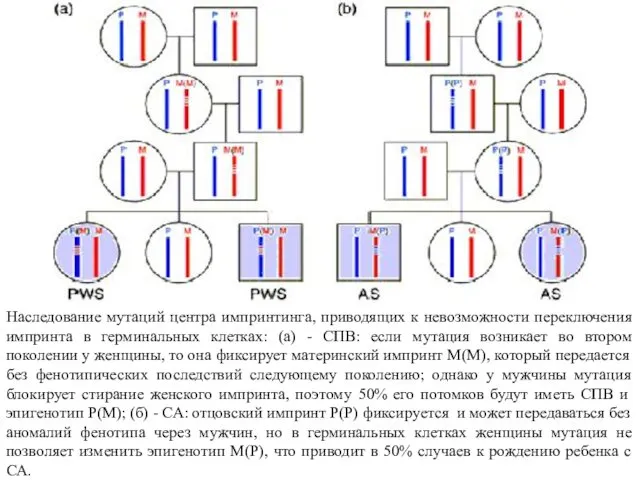
Наследование мутаций центра импринтинга, приводящих к невозможности переключения импринта в герминальных
Наследование мутаций центра импринтинга, приводящих к невозможности переключения импринта в герминальных
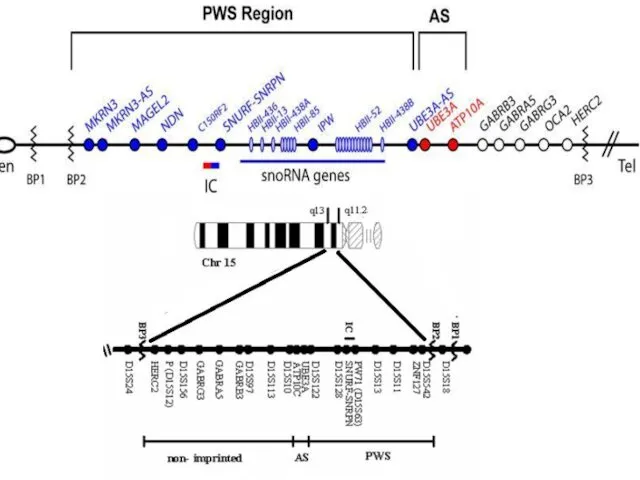
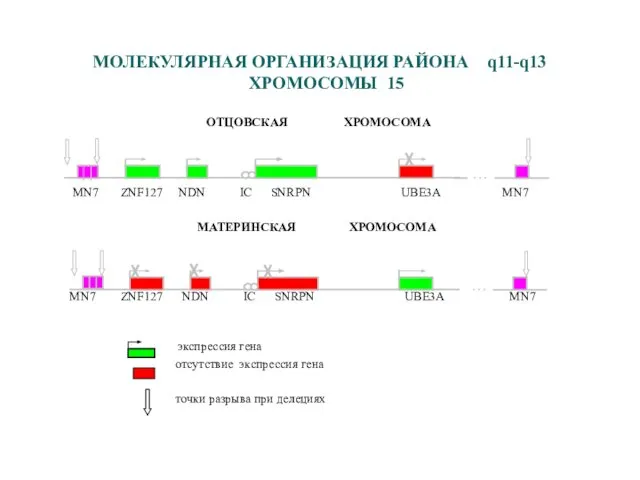
МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ РАЙОНА q11-q13 ХРОМОСОМЫ 15
ОТЦОВСКАЯ ХРОМОСОМА
MN7 ZNF127 NDN IC
МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ РАЙОНА q11-q13 ХРОМОСОМЫ 15
ОТЦОВСКАЯ ХРОМОСОМА
MN7 ZNF127 NDN IC
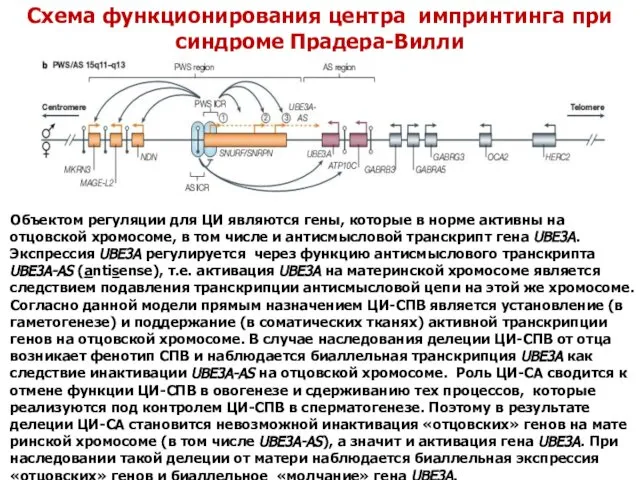
Схема функционирования центра импринтинга при синдроме Прадера-Вилли
Объектом регуляции для ЦИ являются
Схема функционирования центра импринтинга при синдроме Прадера-Вилли
Объектом регуляции для ЦИ являются
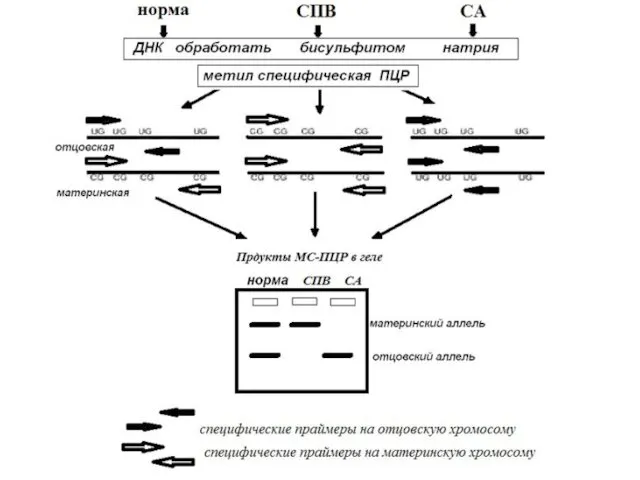
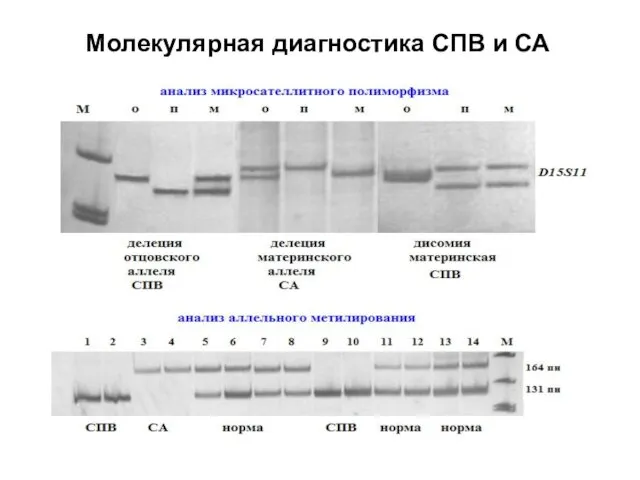
Молекулярная диагностика СПВ и СА
Молекулярная диагностика СПВ и СА
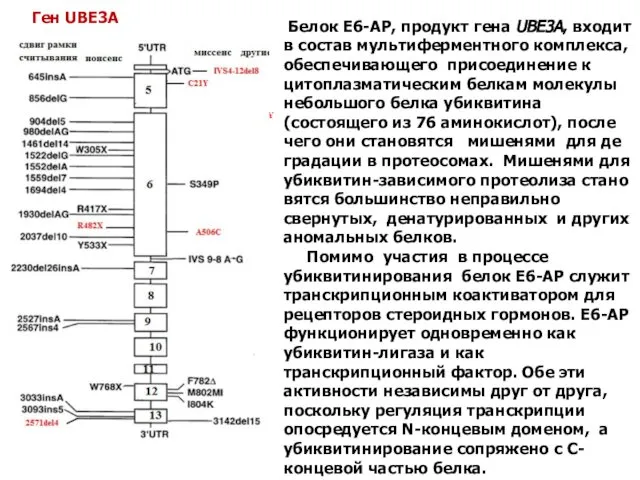
Белок E6-AP, продукт гена UBE3A, входит в состав мультиферментного комплекса,
Белок E6-AP, продукт гена UBE3A, входит в состав мультиферментного комплекса,
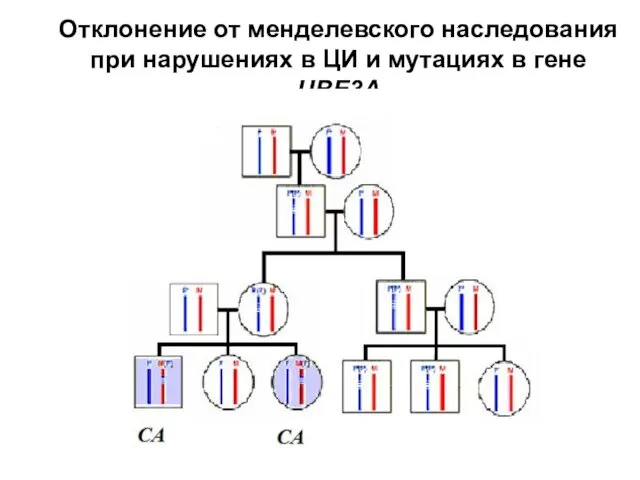
Отклонение от менделевского наследования при нарушениях в ЦИ и мутациях в
Отклонение от менделевского наследования при нарушениях в ЦИ и мутациях в
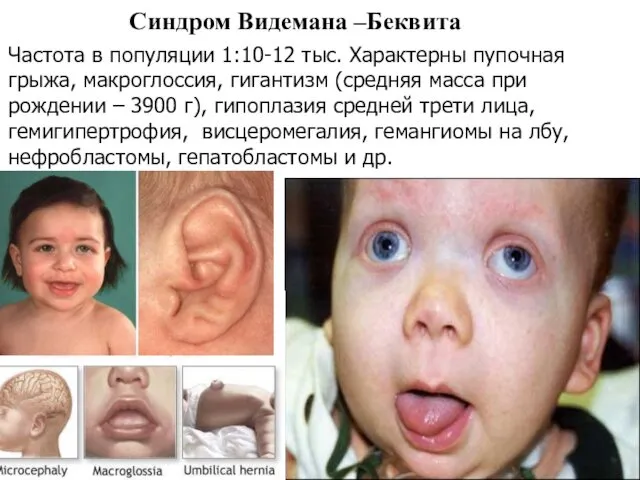
Синдром Видемана –Беквита
Частота в популяции 1:10-12 тыс. Характерны пупочная грыжа, макроглоссия,
Синдром Видемана –Беквита
Частота в популяции 1:10-12 тыс. Характерны пупочная грыжа, макроглоссия,
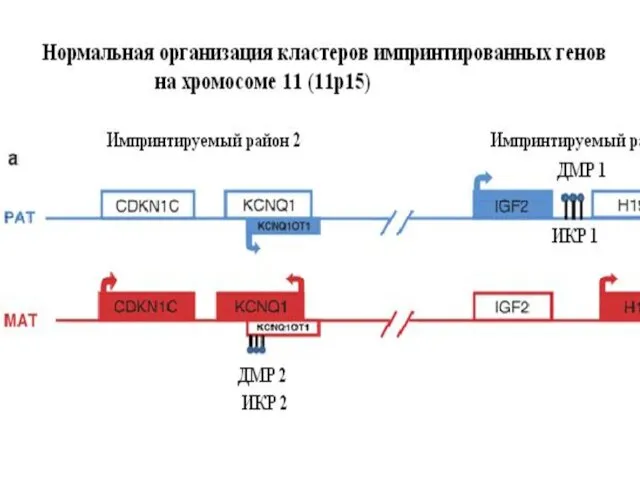
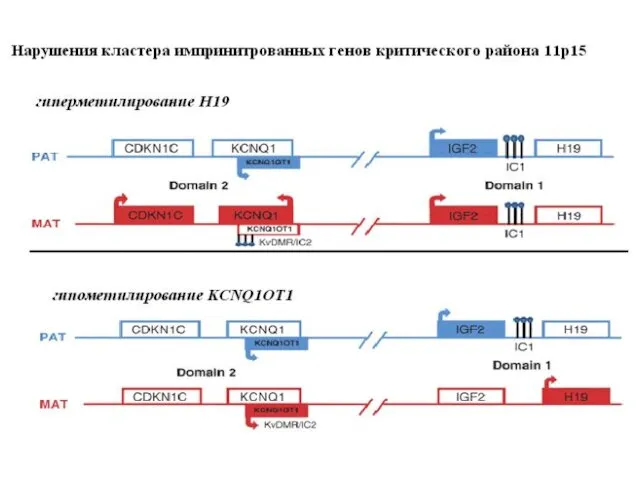
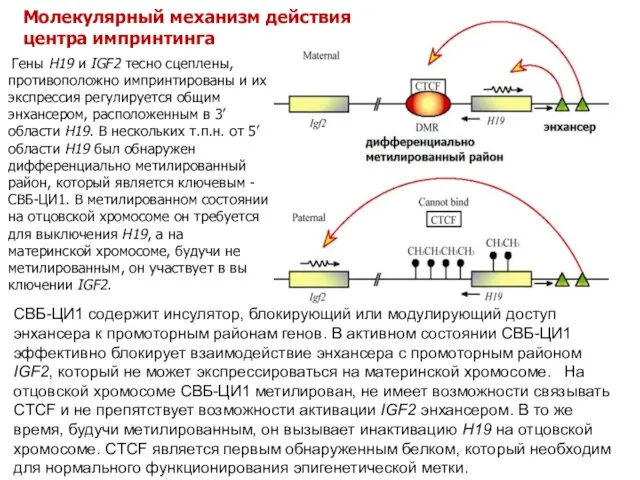
Гены H19 и IGF2 тесно сцеплены, противоположно импринтированы и их
Гены H19 и IGF2 тесно сцеплены, противоположно импринтированы и их

Гено-фенотипические корреляции при СВБ
о
о
о
о
о
о
м
м
м
м
м
о
Гено-фенотипические корреляции при СВБ
о
о
о
о
о
о
м
м
м
м
м
о
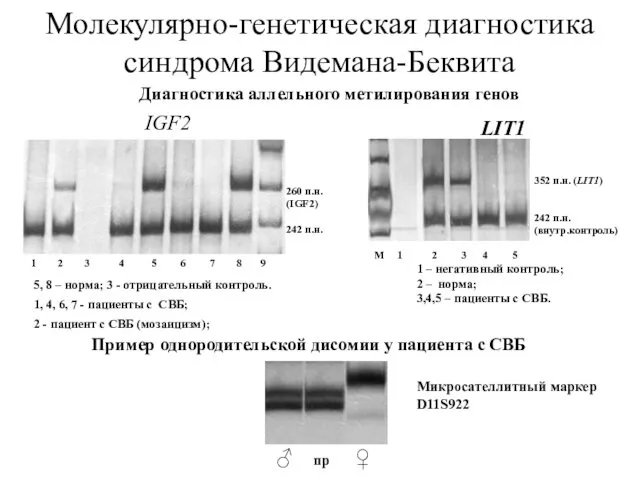
Молекулярно-генетическая диагностика синдрома Видемана-Беквита
1 – негативный контроль;
2 – норма;
3,4,5
Молекулярно-генетическая диагностика синдрома Видемана-Беквита
1 – негативный контроль;
2 – норма;
3,4,5

Пренатальная и постнатальная задержка роста;
Треугольное лицо с выступающим лбом;
Клинодактилия или брахидактилия;
Макроцефалия;
Скелетная
Пренатальная и постнатальная задержка роста;
Треугольное лицо с выступающим лбом;
Клинодактилия или брахидактилия;
Макроцефалия;
Скелетная
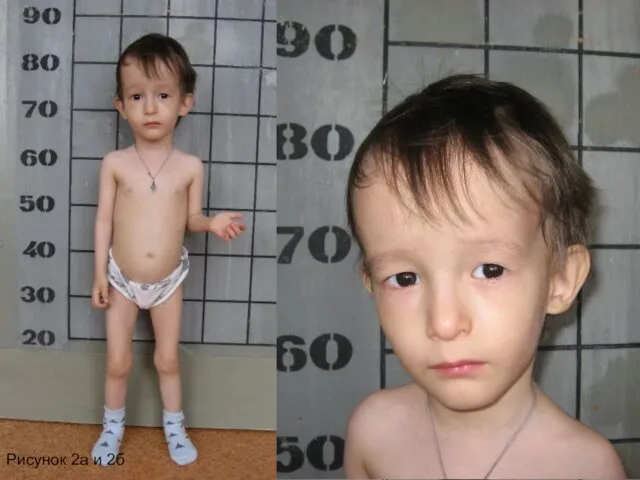
Рисунок 2а и 2б
Рисунок 2а и 2б
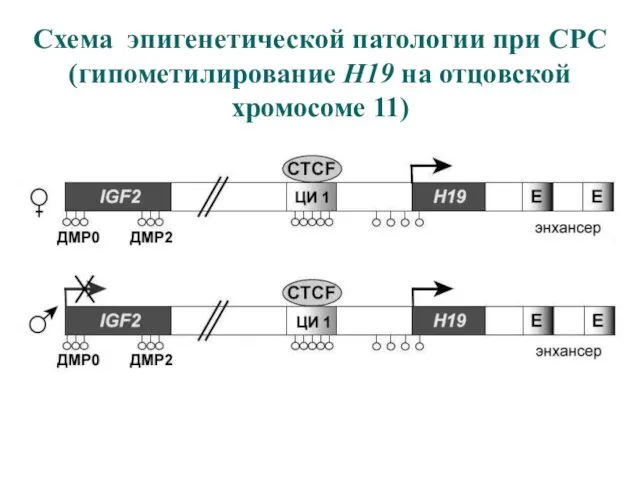
Схема эпигенетической патологии при СРС
(гипометилирование H19 на отцовской хромосоме 11)
Схема эпигенетической патологии при СРС
(гипометилирование H19 на отцовской хромосоме 11)
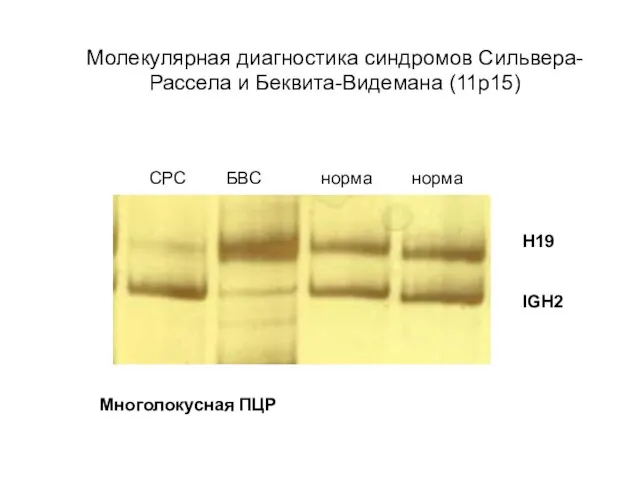
Молекулярная диагностика синдромов Сильвера-Рассела и Беквита-Видемана (11р15)
СРС БВС норма норма
Молекулярная диагностика синдромов Сильвера-Рассела и Беквита-Видемана (11р15)
СРС БВС норма норма

7p11.2-p13. У человека отцовская экспрессия GRB10 установлена в головном и
7p11.2-p13. У человека отцовская экспрессия GRB10 установлена в головном и
Молекулярно-генетические причины синдромов Беквита-Видемана и Сильвера-Рассела
Молекулярно-генетические причины синдромов Беквита-Видемана и Сильвера-Рассела
ХАРАКТЕРИСТИКА ЦЕНТРА ИМПРИНТИНГА
1. Регулирует импринтированные гены в кластере in cis;
2. Имеет
ХАРАКТЕРИСТИКА ЦЕНТРА ИМПРИНТИНГА
1. Регулирует импринтированные гены в кластере in cis;
2. Имеет
 Жири Склад жирів, їх утворення. Жири у природі. Біологічна роль жирів
Жири Склад жирів, їх утворення. Жири у природі. Біологічна роль жирів  Презентация Анти-спид
Презентация Анти-спид The Integumentary System
The Integumentary System Презентация на тему "Мхи, папоротники, хвощи и плауны" - скачать презентации по Биологии
Презентация на тему "Мхи, папоротники, хвощи и плауны" - скачать презентации по Биологии Генератор первичных культур эндотелиальных клеток для тканеинженерных технологий
Генератор первичных культур эндотелиальных клеток для тканеинженерных технологий Экзменационные препараты по гистологии
Экзменационные препараты по гистологии Синичкин день. 12 ноября
Синичкин день. 12 ноября Сбор урожая
Сбор урожая Презентация на тему Экология и здоровье человека
Презентация на тему Экология и здоровье человека  Микроорганизмдер-ақуыздар мен майлардың өнеркәсіптік өнімдері
Микроорганизмдер-ақуыздар мен майлардың өнеркәсіптік өнімдері Керунг
Керунг Биохимия углеводов
Биохимия углеводов В гости к весне. Апрель
В гости к весне. Апрель Характеристики реакций ферментативного катализа Выполнила Кузнецова Екатерина
Характеристики реакций ферментативного катализа Выполнила Кузнецова Екатерина Презентация на тему Устойчивость и динамика экосистем
Презентация на тему Устойчивость и динамика экосистем Центральная нервная система
Центральная нервная система Эндокринные железы
Эндокринные железы Презентация по биологии Лишайники
Презентация по биологии Лишайники Понятие наследственности и изменчивости
Понятие наследственности и изменчивости Презентация на тему ПИНГВИНЫ
Презентация на тему ПИНГВИНЫ  Конструктивный метаболизм микроорганизмов
Конструктивный метаболизм микроорганизмов Эволюционное учение Ч. Дарвина
Эволюционное учение Ч. Дарвина Психогенетика
Психогенетика Весеннее пробуждение растений Обобщение работы над проектом. Презентация выполнена на основе исследований учащихся 2-х класс
Весеннее пробуждение растений Обобщение работы над проектом. Презентация выполнена на основе исследований учащихся 2-х класс Сердечно-сосудистая система
Сердечно-сосудистая система  Озоновые Дыры
Озоновые Дыры  Окружающий мир 3 класс
Окружающий мир 3 класс  К.Линней. Его работы по систематике растений и животных.
К.Линней. Его работы по систематике растений и животных.