- Клеточный механизм действия медиаторов сердечных нервов

Содержание
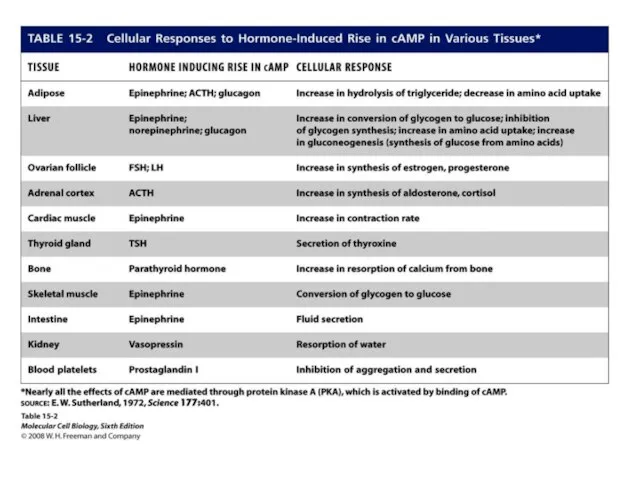
- 2. Норадреналин в кардиомиоцитах: повышение проницаемости для кальция через аденилатциклазный механизм
- 3. Изменение ПД водителя ритма
- 4. Эффекты норадреналина Положительные Инотропный Хронотропный Батмотропный Дромотропный
- 5. К чему приведет избыток норадреналина? ↗темп метаболизма → ↗гликолиз и окисление ЖК → ↗ кислородный запрос
- 6. Ацетилхолин Снижение возбудимости за счет : увеличения проницаемости для калия снижения активности аденилатциклазы.
- 7. Изменение ПД водителя ритма
- 8. Ацетилхолин Отрицательные Хронотропный Инотропный Батмотропный Дромотропный
- 9. К чему приведет избыток ацетилхолина? Снижение возбудимости до невозможности возникновения ПД. Пример – в опыте Гольца
- 10. Физиологические свойства гладких мышц Обладают автоматией. Способны к длительным тоническим сокращениям Сокращаются в ответ на растяжение
- 11. Механизм мышечного сокращения Комплекс Са++ с кальмодулином Активация киназы легких цепей миозина Фосфорилирование головки миозина Образование
- 12. Механизм действия БАВ
- 13. Сосуды иннервируются симпатическими нервами Постганглионарные волокна выделяют НОРАДРЕНАЛИН
- 16. Эндотелий сосудов Саморегуляция клеточного роста и восстановления Местная регуляция сосудистого гладкомышечного тонуса: синтез простагландинов, эндотелинов, оксида
- 18. Клеточные рецепторы делятся на следующие классы мембранные рецепторы, сопряжённые с G-белками рецепторные тирозинкиназы сопряженные с ионными
- 19. Мембранные рецепторы
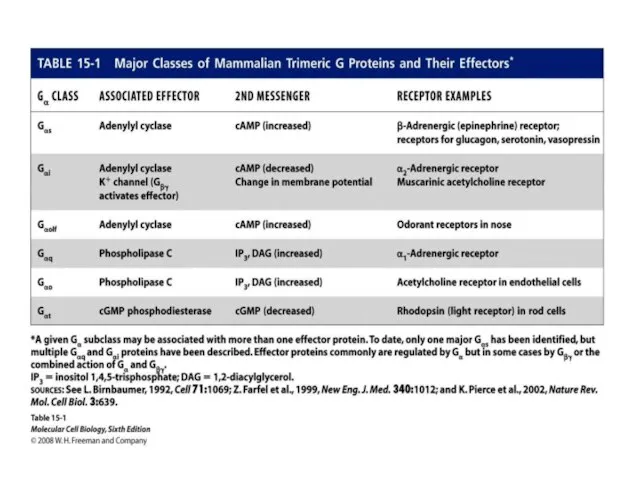
- 20. G-белки - ГТФ-связывающие белки, гуанин-нуклеотидсвязывающие белки Виды G-белков: Gs-белок – стимулирует аденилатциклазу; Gi-белок – ингибирует аденилатциклазу;
- 21. Гетеротримерные G-белки состоят из трёх субъединиц (СЕ): α,β,γ. Функция и специфичность G-белка обычно определяется α-СЕ. Как
- 23. Регуляторный цикл G-белка: 1) Рецептор, активированный лигандом, катализирует освобождение ГДФ, связанного с α-СЕ G-белка и связывание
- 24. 7-доменный мембранный рецептор
- 25. GPCR
- 28. Основные пары «рецептор-G-белок»
- 29. Gs-белок – стимулирует аденилатциклазу; Gi-белок – ингибирует аденилатциклазу;
- 30. Эффект норадреналина в клетках миокарда обусловлен взаимодействием с β1-адренорецепторами
- 32. Ацетилхолин Влияние АХ сказывается преимущественно в уменьшении ЧСС. Однако уменьшение ЧСС вместе с некоторым ослаблением силы
- 34. Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путем фосфорилирования остатков аминокислот, имеющих гидроксильные
- 35. Протеинкиназы Протеинкиназа А, или цАМФ-зависимая протеинкиназа, Протеинкиназы С это семейство протеинкиназ, содержащее порядка десяти изоферментов, которые
- 37. цАМФ-зависимая протеинкиназа (А) Состоит из 2-х регуляторных и 2-х каталитических СЕ. Регуляторные СЕ подавляют ферментативную активность
- 38. Протеинкиназа А
- 39. В сердце: 1) L-тип Ca-каналов, 2) Рианодиновый рецептор (RyR), 3) Тропонин I, 4) миозин-связывающий белок C,
- 40. Ключевым регулятором расслабления миокарда является белок мембраны саркоплазматического ретикулума фосфоламбан Фосфорилирование фосфоламбана при участии вторичного посредника
- 41. Протеинкиназа А находится вблизи своих мишеней благодаря заякоривающим белкам (AKAP – A-kinase anchored protein)
- 42. ПКА в мышечных клетках и клетках печени
- 43. Протеинкиназа С
- 44. Протеинкиназа G цГМФ зависимые протеинкиназы ( ПК-G ) не способны к аутофосфорилированию.
- 48. Скачать презентацию
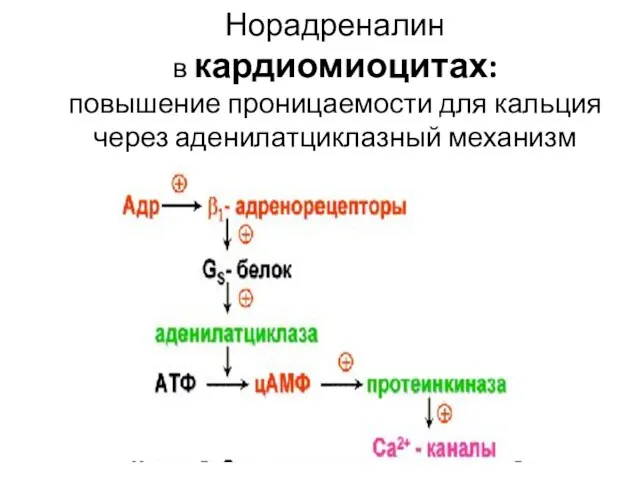
Норадреналин
в кардиомиоцитах:
повышение проницаемости для кальция через аденилатциклазный механизм
Норадреналин
в кардиомиоцитах:
повышение проницаемости для кальция через аденилатциклазный механизм
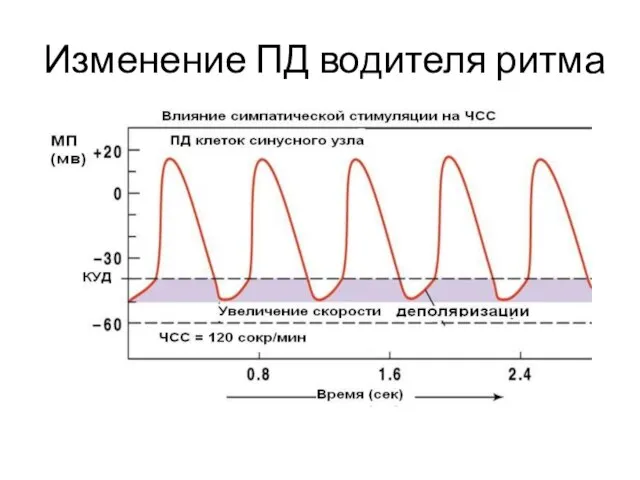
Изменение ПД водителя ритма
Изменение ПД водителя ритма

Эффекты норадреналина
Положительные
Инотропный
Хронотропный
Батмотропный
Дромотропный
Эффекты норадреналина
Положительные
Инотропный
Хронотропный
Батмотропный
Дромотропный

К чему приведет избыток норадреналина?
↗темп метаболизма → ↗гликолиз и окисление ЖК
К чему приведет избыток норадреналина?
↗темп метаболизма → ↗гликолиз и окисление ЖК

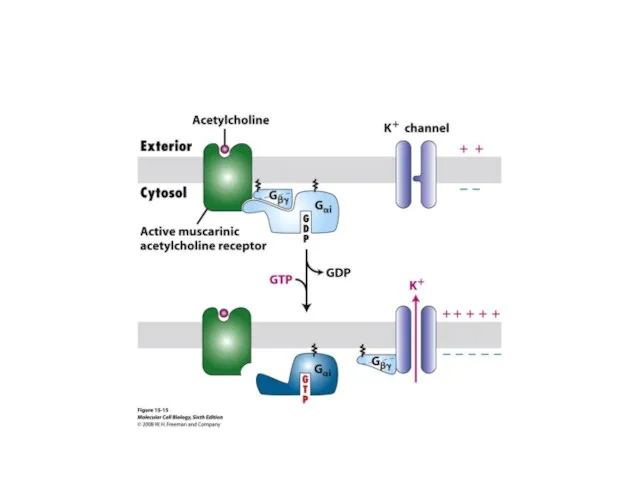
Ацетилхолин
Снижение возбудимости за счет :
увеличения проницаемости для калия
снижения активности аденилатциклазы.
Ацетилхолин
Снижение возбудимости за счет :
увеличения проницаемости для калия
снижения активности аденилатциклазы.
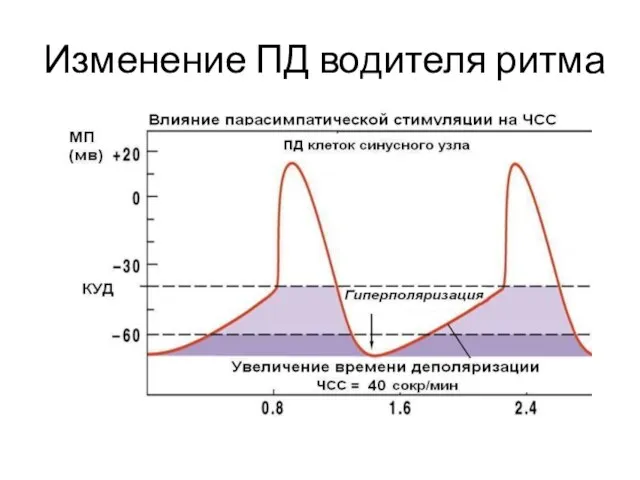
Изменение ПД водителя ритма
Изменение ПД водителя ритма

Ацетилхолин
Отрицательные
Хронотропный
Инотропный
Батмотропный
Дромотропный
Ацетилхолин
Отрицательные
Хронотропный
Инотропный
Батмотропный
Дромотропный

К чему приведет избыток ацетилхолина?
Снижение возбудимости до невозможности возникновения ПД.
Пример
К чему приведет избыток ацетилхолина?
Снижение возбудимости до невозможности возникновения ПД.
Пример

Физиологические свойства
гладких мышц
Обладают автоматией.
Способны к длительным тоническим сокращениям
Сокращаются в
Физиологические свойства
гладких мышц
Обладают автоматией.
Способны к длительным тоническим сокращениям
Сокращаются в

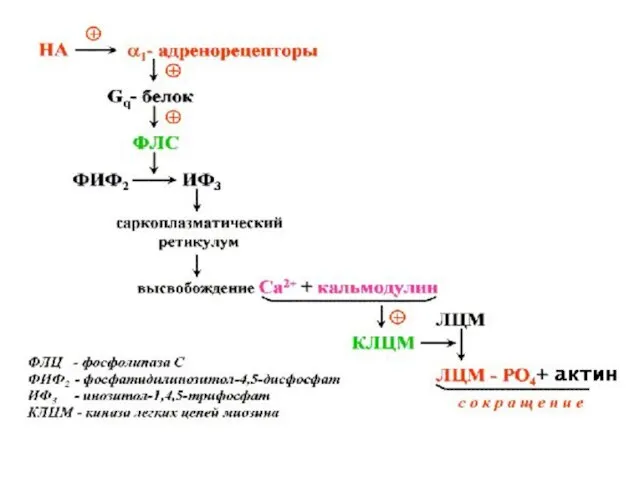
Механизм мышечного сокращения
Комплекс Са++ с кальмодулином
Активация киназы легких цепей миозина
Фосфорилирование головки
Механизм мышечного сокращения
Комплекс Са++ с кальмодулином
Активация киназы легких цепей миозина
Фосфорилирование головки
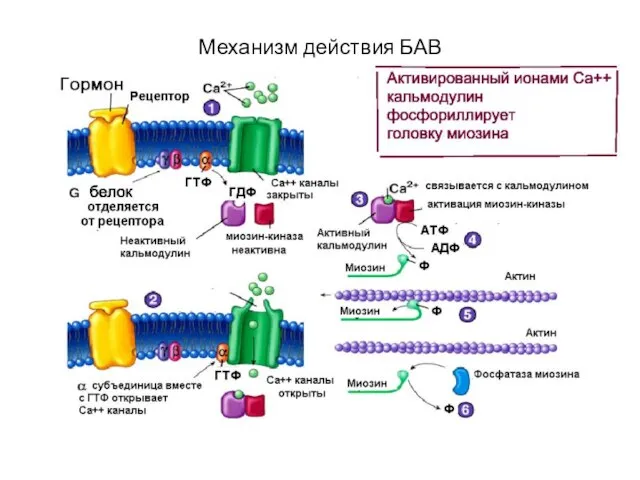
Механизм действия БАВ
Механизм действия БАВ

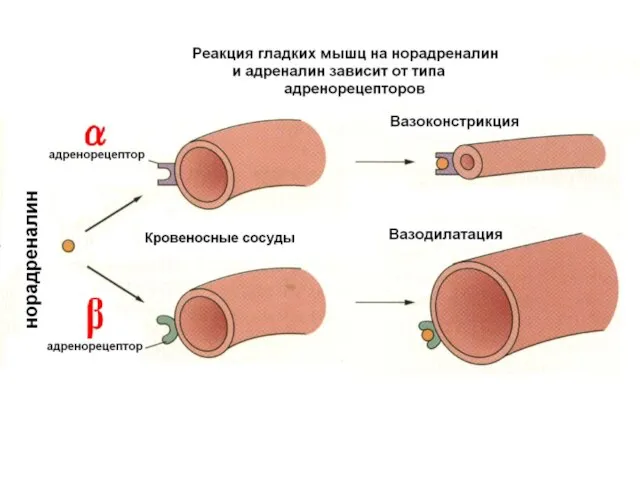
Сосуды иннервируются симпатическими нервами
Постганглионарные волокна выделяют
НОРАДРЕНАЛИН
Сосуды иннервируются симпатическими нервами
Постганглионарные волокна выделяют
НОРАДРЕНАЛИН

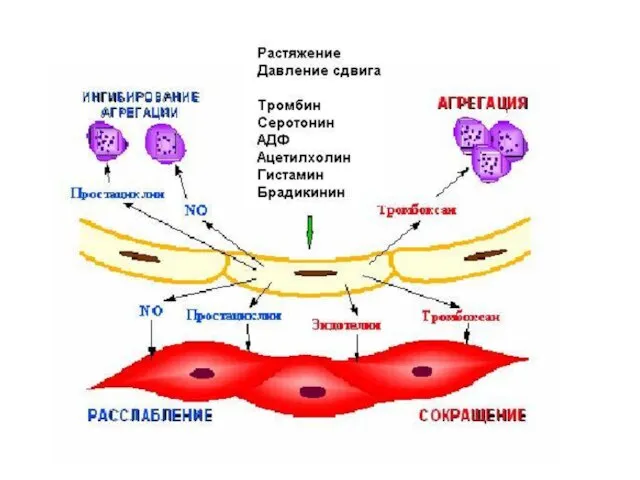
Эндотелий сосудов
Саморегуляция клеточного роста и восстановления
Местная регуляция сосудистого гладкомышечного тонуса: синтез
Эндотелий сосудов
Саморегуляция клеточного роста и восстановления
Местная регуляция сосудистого гладкомышечного тонуса: синтез

Клеточные рецепторы делятся на следующие классы
мембранные
рецепторы, сопряжённые с G-белками
рецепторные
Клеточные рецепторы делятся на следующие классы
мембранные
рецепторы, сопряжённые с G-белками
рецепторные
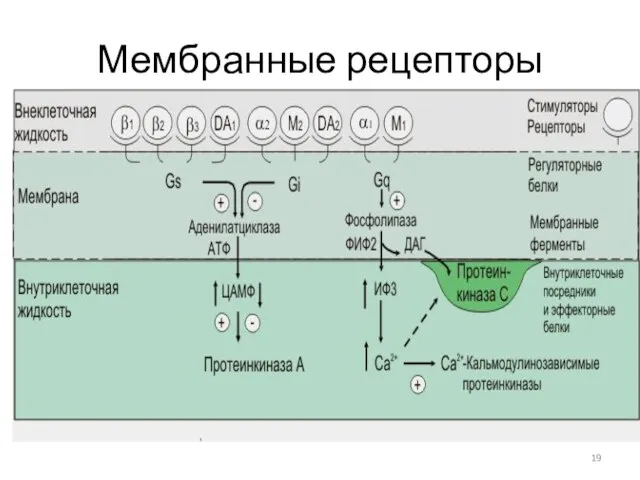
Мембранные рецепторы
Мембранные рецепторы

G-белки - ГТФ-связывающие белки, гуанин-нуклеотидсвязывающие белки
Виды G-белков:
Gs-белок – стимулирует аденилатциклазу;
Gi-белок –
G-белки - ГТФ-связывающие белки, гуанин-нуклеотидсвязывающие белки
Виды G-белков:
Gs-белок – стимулирует аденилатциклазу;
Gi-белок –

Гетеротримерные G-белки состоят из трёх субъединиц (СЕ): α,β,γ.
Функция и специфичность
Гетеротримерные G-белки состоят из трёх субъединиц (СЕ): α,β,γ.
Функция и специфичность
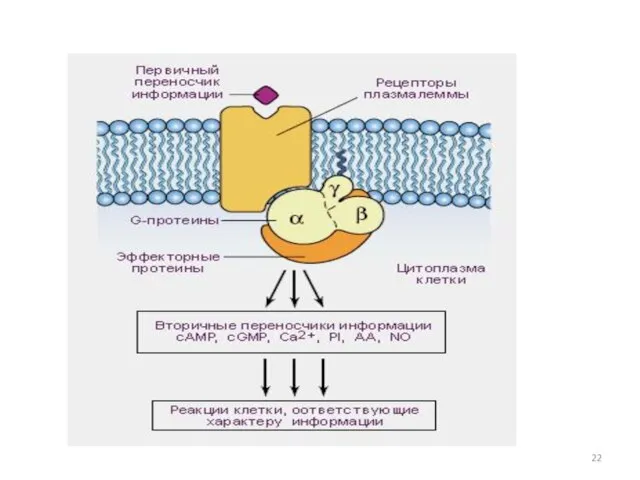

Регуляторный цикл G-белка:
1) Рецептор, активированный лигандом,
катализирует освобождение ГДФ, связанного с
Регуляторный цикл G-белка:
1) Рецептор, активированный лигандом,
катализирует освобождение ГДФ, связанного с
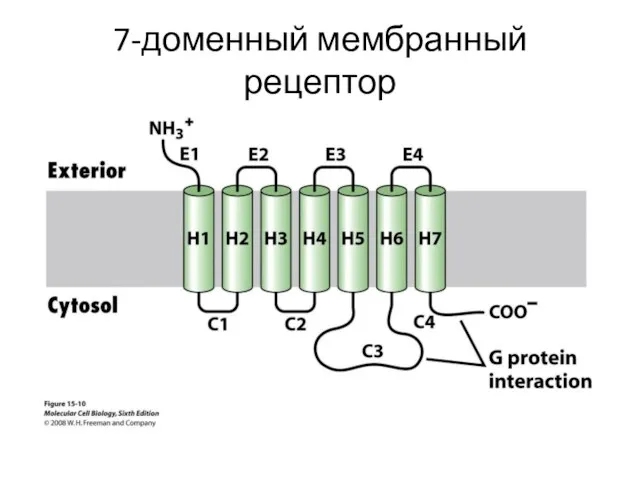
7-доменный мембранный рецептор
7-доменный мембранный рецептор
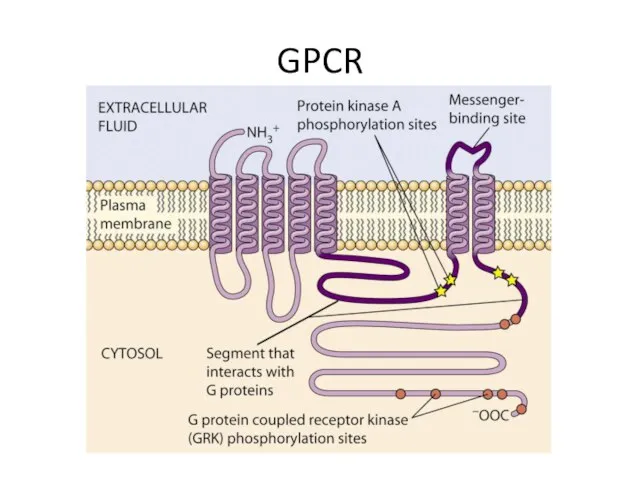
GPCR
GPCR
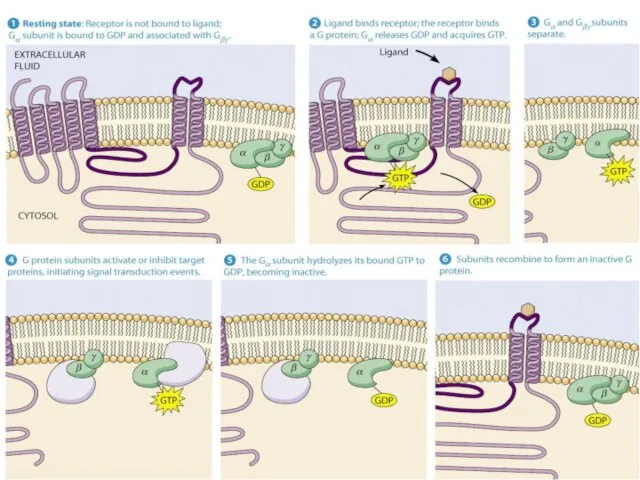
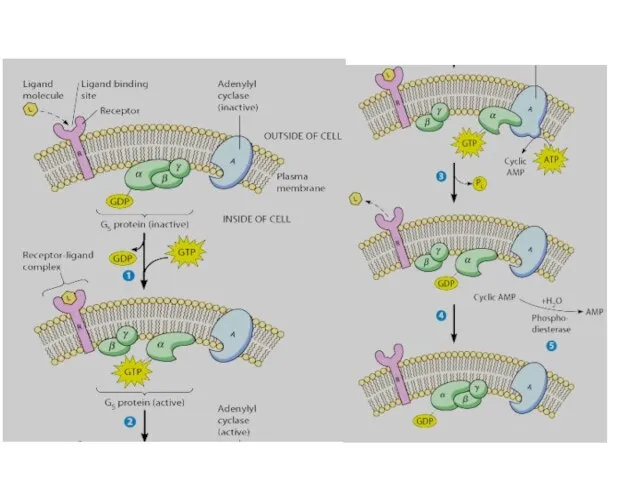

Основные пары «рецептор-G-белок»
Основные пары «рецептор-G-белок»
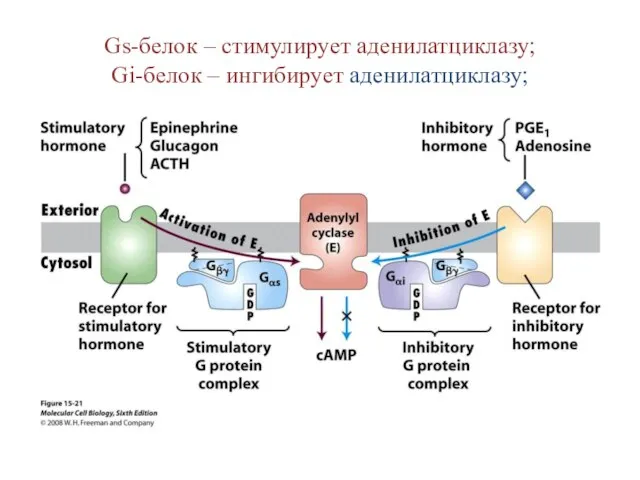
Gs-белок – стимулирует аденилатциклазу;
Gi-белок – ингибирует аденилатциклазу;
Gs-белок – стимулирует аденилатциклазу;
Gi-белок – ингибирует аденилатциклазу;

Эффект норадреналина в клетках миокарда обусловлен взаимодействием с
β1-адренорецепторами
Эффект норадреналина в клетках миокарда обусловлен взаимодействием с
β1-адренорецепторами
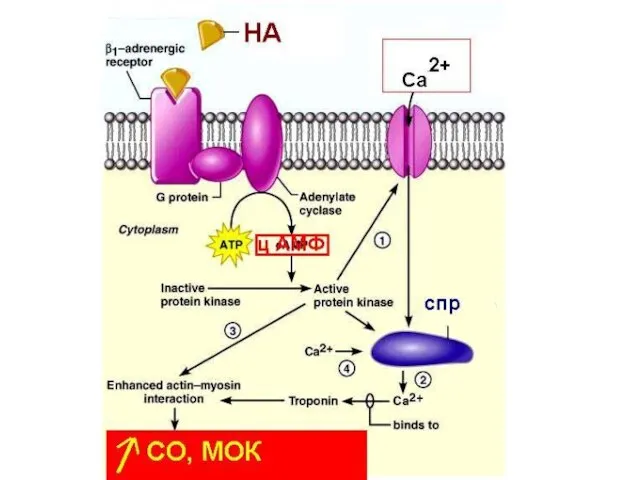

Ацетилхолин
Влияние АХ сказывается преимущественно в уменьшении ЧСС.
Однако уменьшение ЧСС вместе с
Ацетилхолин
Влияние АХ сказывается преимущественно в уменьшении ЧСС.
Однако уменьшение ЧСС вместе с

Протеинкина́зы
— подкласс ферментов киназ (фосфотрансфераз).
Протеинкиназы модифицируют другие белки путем фосфорилирования
Протеинкина́зы
— подкласс ферментов киназ (фосфотрансфераз).
Протеинкиназы модифицируют другие белки путем фосфорилирования

Протеинкиназы
Протеинкиназа А, или цАМФ-зависимая протеинкиназа,
Протеинкиназы С это семейство протеинкиназ, содержащее
Протеинкиназы
Протеинкиназа А, или цАМФ-зависимая протеинкиназа,
Протеинкиназы С это семейство протеинкиназ, содержащее
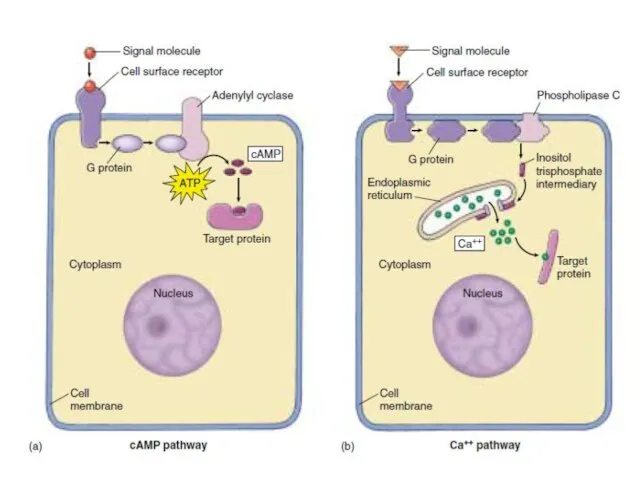

цАМФ-зависимая протеинкиназа (А)
Состоит из 2-х регуляторных и 2-х каталитических СЕ.
Регуляторные
цАМФ-зависимая протеинкиназа (А)
Состоит из 2-х регуляторных и 2-х каталитических СЕ.
Регуляторные
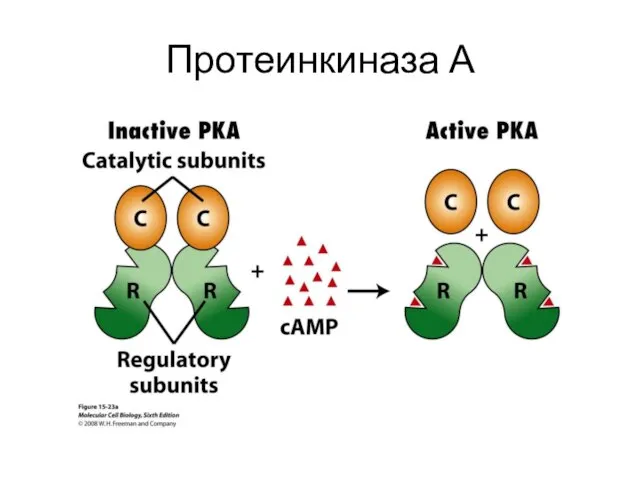
Протеинкиназа А
Протеинкиназа А

В сердце:
1) L-тип Ca-каналов,
2) Рианодиновый рецептор (RyR),
3) Тропонин I,
4) миозин-связывающий
В сердце:
1) L-тип Ca-каналов,
2) Рианодиновый рецептор (RyR),
3) Тропонин I,
4) миозин-связывающий

Ключевым регулятором расслабления миокарда является белок мембраны саркоплазматического ретикулума фосфоламбан
Фосфорилирование
Ключевым регулятором расслабления миокарда является белок мембраны саркоплазматического ретикулума фосфоламбан
Фосфорилирование
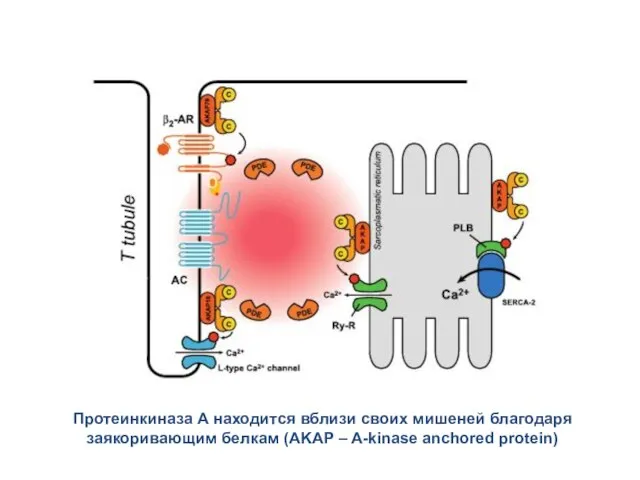
Протеинкиназа А находится вблизи своих мишеней благодаря заякоривающим белкам (AKAP –
Протеинкиназа А находится вблизи своих мишеней благодаря заякоривающим белкам (AKAP –
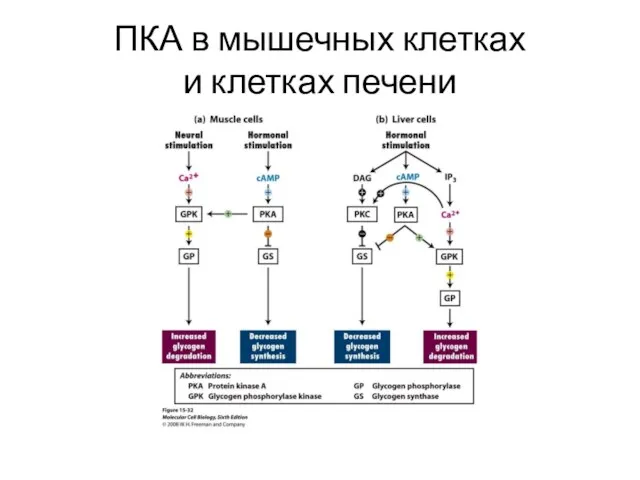
ПКА в мышечных клетках
и клетках печени
ПКА в мышечных клетках
и клетках печени
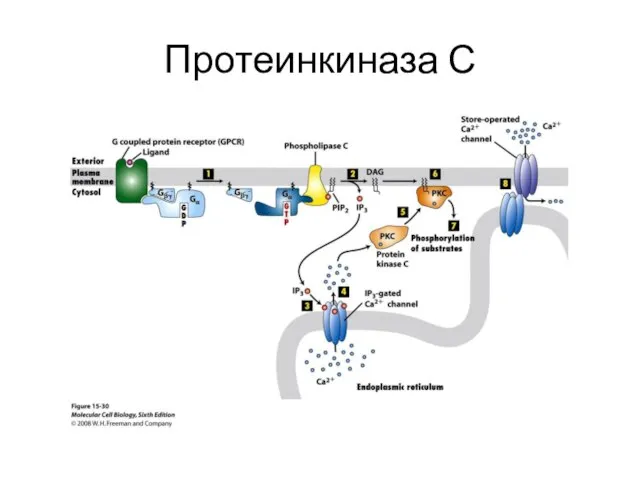
Протеинкиназа С
Протеинкиназа С
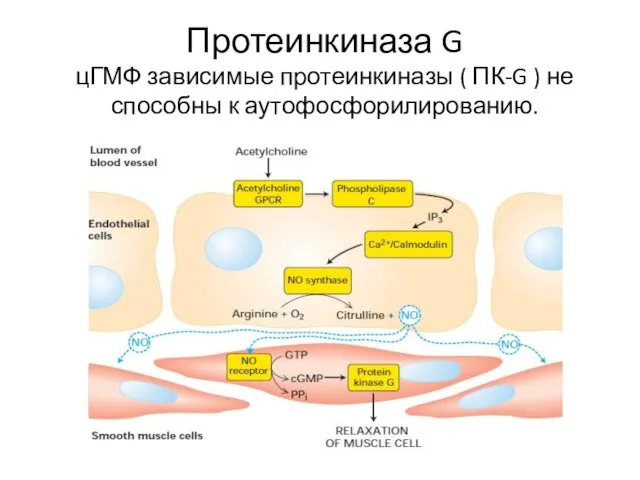
Протеинкиназа G
цГМФ зависимые протеинкиназы ( ПК-G ) не способны к аутофосфорилированию.
Протеинкиназа G
цГМФ зависимые протеинкиназы ( ПК-G ) не способны к аутофосфорилированию.
 Презентация по биологии Оплодотворение и развитие организма
Презентация по биологии Оплодотворение и развитие организма  Прогулка под водой внеклассное мероприятие по биологии, 7 класс Автор: Лаврентьева Снежана Павловна, учитель химии и биологи
Прогулка под водой внеклассное мероприятие по биологии, 7 класс Автор: Лаврентьева Снежана Павловна, учитель химии и биологи По cледам птиц и животных
По cледам птиц и животных Презентация на тему Неорганические вещества, входящие в состав клетки
Презентация на тему Неорганические вещества, входящие в состав клетки  Презентация на тему Хрящевые рыбы
Презентация на тему Хрящевые рыбы  Природа родного края
Природа родного края Физиология бактерий. Метаболизм. (Часть 1)
Физиология бактерий. Метаболизм. (Часть 1) Бионика как одно из направлений биологии и кибернетики. Лекция 36
Бионика как одно из направлений биологии и кибернетики. Лекция 36 Биотехнология металлов
Биотехнология металлов Исследовательская работа по биологии на тему «В гостях у белой кувшинки» Ученицы 6 класса МОУ «СОШ с. Дмитриевка Духовницкого р
Исследовательская работа по биологии на тему «В гостях у белой кувшинки» Ученицы 6 класса МОУ «СОШ с. Дмитриевка Духовницкого р Презентация на тему "Гаметогенез. Оплодотворение" - скачать презентации по Биологии
Презентация на тему "Гаметогенез. Оплодотворение" - скачать презентации по Биологии Группы генов
Группы генов Организм – единое целое
Организм – единое целое Домашние лабораторные работы по биологии. 8 класс
Домашние лабораторные работы по биологии. 8 класс Презентация на тему "ШМО" - скачать презентации по Биологии
Презентация на тему "ШМО" - скачать презентации по Биологии Фрукты. Выращивание фруктов
Фрукты. Выращивание фруктов Царство бактерии
Царство бактерии Задачи на самопроверку перед промежуточной аттестацией
Задачи на самопроверку перед промежуточной аттестацией Мышечная рецепция
Мышечная рецепция Введение в молекулярную биологию и генетику
Введение в молекулярную биологию и генетику Царство Грибы
Царство Грибы Выращивание лимона в домашних условиях
Выращивание лимона в домашних условиях Презентация на тему "ЗЕМНОВОДНЫЕ" - скачать бесплатно презентации по Биологии
Презентация на тему "ЗЕМНОВОДНЫЕ" - скачать бесплатно презентации по Биологии Презентация по биологии Слуховой анализатор. Гигиена слуха. 8 класс
Презентация по биологии Слуховой анализатор. Гигиена слуха. 8 класс Жизнь в морях и океанах_
Жизнь в морях и океанах_ Презентація На тему:”Південноазійський тропічний центр різноманітності і походження культурних рослин” Виконав учень 11-А класу Острожчук Богдан
Презентація На тему:”Південноазійський тропічний центр різноманітності і походження культурних рослин” Виконав учень 11-А класу Острожчук Богдан  Тип Членистоногі
Тип Членистоногі Презентация по биологии Отряд хищные
Презентация по биологии Отряд хищные