- Мембранный потенциал

Содержание
- 2. Потенциал покоя (ПП) нейрона – его постоянный отрицательный заряд, равный в среднем -70 мВ. Измерить ПП
- 3. Механизмы возникновения мембранного потенциала (Ю.Бернштейн, 1902). Мембрана клеток в покое обладает высокой проницаемостью для ионов калия,
- 4. Наличие ПП – результат жизнедеятельности нейрона, совместного функционирования всех биополимеров и органоидов клетки; погибший нейрон быстро
- 5. В результате работы Na+-K+-АТФазы в нейроне оказывается примерно в 10 раз меньше Na+ и в 30
- 6. выход К+ сопровождается накоплением в цитоплазме отрицательного заряда. Этот отрицательный заряд мешает диф-фузии и в конце
- 7. ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в межкле-точную среду. Вальтер Нернст (Ноб.пр.
- 8. ПП = -91 мВ («равновесный потенциал» для К+) В реальной клетке ПП находится ближе к нулю
- 9. внутриклеточная среда Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда
- 10. Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название –
- 11. Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название –
- 12. В целом ПП зависит от 3-х главных факторов: - диффузии K+ из клетки; - диффузии Na+
- 13. В 1785 году Визеринг сообщил об использовании листьев наперстянки для лечения сердечной недостаточности. Действующим началом являлся
- 14. Наперстянка (дигиталис) Симптомы отравления: Сердечный приступ Тошнота, рвота Диарея Головная боль Медленный пульс Делирий Первая помощь
- 15. Интересные факты о наперстянке Один из побочных эффектов лекарств с наперстянкой – изменённое восприятие цветов Воздействие
- 16. Уравнение Гольдмана-Ходжкина-Катца
- 17. Биоэлектрические явления в возбудимых тканях Физиологию возбудимых тканей изучает электрофизиология. Электрофизиология –исследует электрические проявления жизнедеятельности клеток,
- 18. Основные задачи электрофизиологии: 1. Изучение физических и химических механизмов электрических процессов в живых тканях, т.е. природы
- 19. В основе реакций организма лежит раздражимость - способность реагировать на действие различных факторов изменением функции. Возбудимость
- 20. Явления связанные с природным электричеством (молнии, статическое электричество, электрические рыбы и др.) были известны очень давно
- 21. Бенджамин Франклин (1706—1790) Разработал (1750) теорию электрических явлений, согласно которой электричество представляет особую “электрическую жидкость», пронизывающую
- 23. Впервые искусственную электрическую искру получили от электрической машины, изобретённой Герике - шар из серы, насаженный на
- 24. Первые электрофизиологические опыты. Луиджи Гальвани (1737-1798) Он обратил внимание на сокращение препарата задних лапок лягушки при
- 25. Итальянский физиолог Луиджи Гальвани из Болоньи сделал важное наблюдение. Он заметил, что тело мертвой лягушки вздрагивало
- 26. Лаборатория Гальвани
- 27. Алессандро Вольта (1745-1827) Вольта повторил опыт Гальвани, но объяснил, его результаты тем, что в цепи из
- 28. Второй опыт Л. Гальвани Для того, чтобы доказать, что ткани животных тоже способны генерировать электрический ток
- 29. Первый «балконный» опыт Л.Гальвани (1786 г). Гальвани повторил этот опыт в условиях лаборатории, прикасаясь к препаратам
- 30. Триумф исследовательской программы Дж. Альдини произошел в Лондоне 17 января 1803 года. В этот день, имевший
- 31. Дальнейшие исследования В 1838 г. Матеуччи обнаружил, что можно зарегистрировать электрический ток (ток покоя), который течет
- 32. Что такое возбудимые ткани Это ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию
- 33. Все электрические процессы разворачиваются на цитоплазматической мембране. Клеточные мембраны состоят из жидкой фазы липидов и встроенных
- 34. Белковые молекулы частично погружены в слой липидов, либо с внеклеточной стороны, либо с внешней стороны. Некоторые
- 35. Функции клеточных мембран: Барьерная функция – участие в механизмах электрогенеза, формирования ПП и ПД. Регуляторная –
- 36. сома (тело) нейрона: размер 5-100 мкм, разнообразие форм (пирамидная, звездчатая, грушевидная и др.); функция – обработка
- 37. Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов – потенциалов действия (ПД). Источником является
- 38. ПД – универсальный ответ нервной клетки на стимуляцию 20 мВ: пороговый стимул при ПП= -70 мВ
- 39. В основе этих процессов – открывание и закрывание электрочувствительных Na+- и К+-каналов. Эти каналы имеют створки,
- 40. В основе этих процессов – открывание и закрывание электрочувствительных Na+- и К+-каналов. Эти каналы имеют створки,
- 41. Именно разная скорость открытия Na+-каналов и К+-каналов позволяет возникнуть сначала восходящей, а затем – нисходящей фазе
- 42. Именно разная скорость открытия Na+-каналов и К+-каналов позволяет возникнуть сначала восходящей, а затем – нисходящей фазе
- 43. Поскольку К+-каналы начинают закрываться довольно поздно (вслед за проходом уровня -50 мВ), заряд нейрона после ПД
- 44. тетродотоксин – яд рыбы фугу (аминогруппа работает как «пробка» для Na+-канала) В результате действия токсина прекра
- 45. ТЭА – тетраэтиламмоний: работает как «пробка» по отношению к К+-каналу. В результате восходящая фаза ПД изменяется
- 46. Проведение возбуждения в нервных волокнах При распространении возбуждения по безмиелиновому нервному волокну местные токи, возникающие между
- 47. РАСПРОСТРАНЕНИЕ ПД. Если ПД возник хотя бы в одной точке мембраны ней- рона – он распространяется
- 48. РАСПРОСТРАНЕНИЕ ПД. Если ПД возник хотя бы в одной точке мембраны ней- рона – он распространяется
- 49. Скорость такого распространения низка и не пре-вышает у человека 1-2 м/с (диаметр аксона 1-2 мкм). Но:
- 50. Батрахотоксин: токсин кожи некоторых лягушек-листолазов; модифициро- ванный стероидный гормон насекомых (?). Токсин проникает внутрь клетки и
- 51. Мозг человека содержит ≈ 100 миллиардов нейронов. Каждый нейрон образует контакты в среднем с 1000 других
- 52. Структурные требования к электрическому механизму синаптической передачи 1.Тесное прилегание пре- и постсинаптических мембран 2. Наличие системы
- 53. Физиологическое значение – синхронность ответа множества клеток на приходящий сигнал, несмотря на ослабление сигнала В электрическом
- 54. Менее распространены, чем химические (≈ 1%). Прилегающие мембраны соединены щелевым контактом. Ток течет из одной клетки
- 55. Структурные требования к химическому механизму синаптической передачи Взаимодействие трех структур: (а) пресинаптической –электрически управляемая секреция медиатора;
- 56. Медиаторы Требования к молекулярным свойствам медиаторов Высокая скорость диффузии, а значит низкий молекулярный вес Относительная простота
- 57. Электронная микроскопия химических синапсов Пре- Пре- Пост- Пост- Видны: пресинаптическая нервная терминаль, содержащая синаптические пузырьки, заполненные
- 58. Значение колокализации Са2+ каналов и участков экзоцитоза Короткая дистанция – открытие одного канала достаточно для экзоцитоза
- 59. Экзоцитоз содержимого синаптических пузырьков Непременными условиями экзоцитоза являются:(i) контакт синаптического пузырька с определенным участком пресинаптической мембраны;
- 60. Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 1. Основные белки синаптических пузырьков: Synaptobrevin Synaptotagmin
- 61. Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 2. Пузырек прикреплен к мембране (docking) и
- 62. Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 3 Са2+ зависимый этап. Деполяризация открывает кальциевые
- 63. Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 4 Финал. Произошло слияние (fusion) пузырька с
- 64. Разнообразие морфологии нейронов
- 66. Скачать презентацию
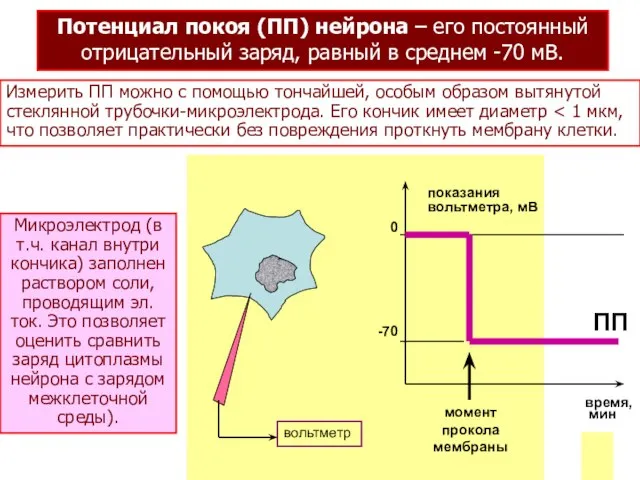
Потенциал покоя (ПП) нейрона – его постоянный
отрицательный заряд, равный в
Потенциал покоя (ПП) нейрона – его постоянный
отрицательный заряд, равный в
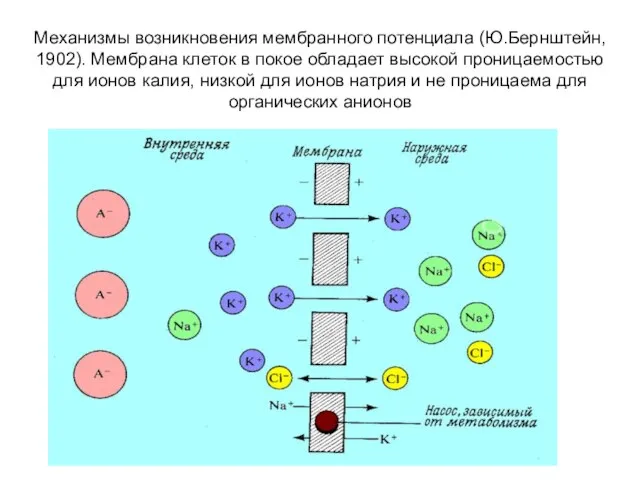
Механизмы возникновения мембранного потенциала (Ю.Бернштейн, 1902). Мембрана клеток в покое обладает
Механизмы возникновения мембранного потенциала (Ю.Бернштейн, 1902). Мембрана клеток в покое обладает
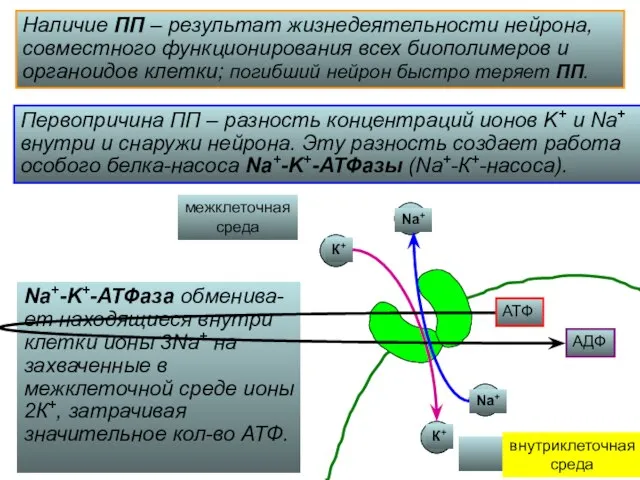
Наличие ПП – результат жизнедеятельности нейрона,
совместного функционирования всех биополимеров и
органоидов
Наличие ПП – результат жизнедеятельности нейрона,
совместного функционирования всех биополимеров и
органоидов

В результате работы Na+-K+-АТФазы в нейроне оказывается
примерно в 10 раз
В результате работы Na+-K+-АТФазы в нейроне оказывается
примерно в 10 раз
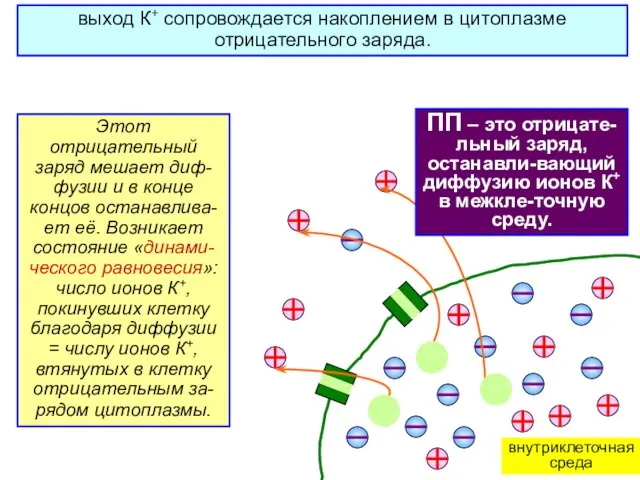
выход К+ сопровождается накоплением в цитоплазме отрицательного заряда.
Этот отрицательный заряд мешает
выход К+ сопровождается накоплением в цитоплазме отрицательного заряда.
Этот отрицательный заряд мешает
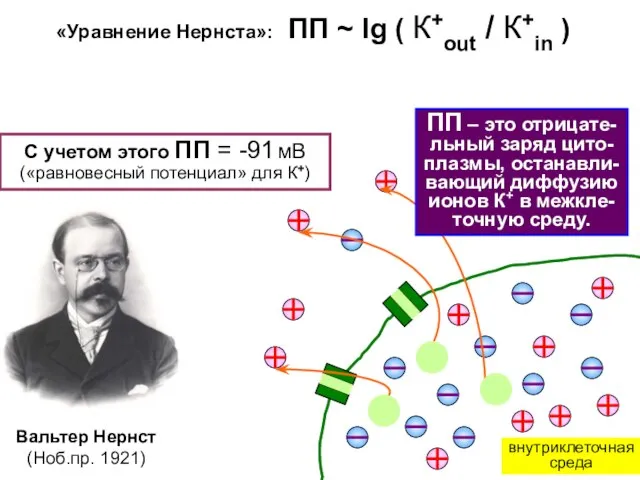
ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в
ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в
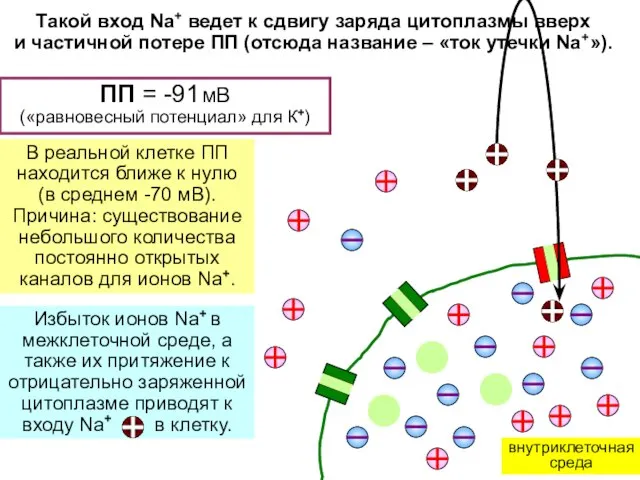
ПП = -91 мВ
(«равновесный потенциал» для К+)
В реальной клетке ПП
находится
ПП = -91 мВ
(«равновесный потенциал» для К+)
В реальной клетке ПП
находится
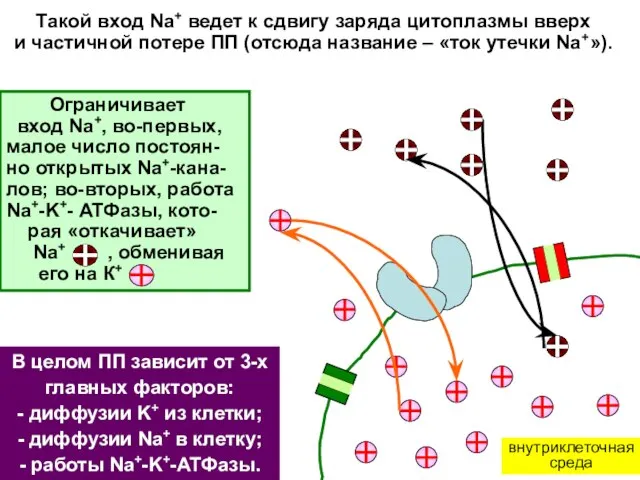
внутриклеточная
среда
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
внутриклеточная
среда
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
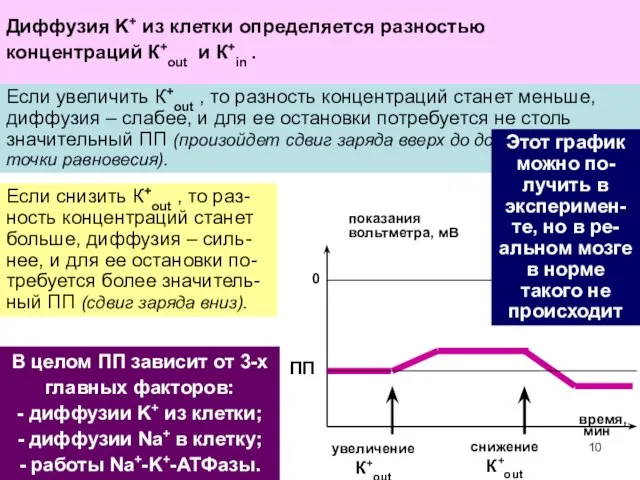
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере

Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере

В целом ПП зависит от 3-х главных факторов:
- диффузии K+ из
В целом ПП зависит от 3-х главных факторов:
- диффузии K+ из

В 1785 году Визеринг сообщил об использовании листьев наперстянки для лечения
В 1785 году Визеринг сообщил об использовании листьев наперстянки для лечения

Наперстянка (дигиталис)
Симптомы отравления:
Сердечный приступ
Тошнота, рвота
Диарея
Головная боль
Медленный пульс
Делирий
Первая помощь при отравлении: очистить
Наперстянка (дигиталис)
Симптомы отравления:
Сердечный приступ
Тошнота, рвота
Диарея
Головная боль
Медленный пульс
Делирий
Первая помощь при отравлении: очистить

Интересные факты о наперстянке
Один из побочных эффектов лекарств с наперстянкой –
Интересные факты о наперстянке
Один из побочных эффектов лекарств с наперстянкой –

Уравнение Гольдмана-Ходжкина-Катца
Уравнение Гольдмана-Ходжкина-Катца

Биоэлектрические явления в возбудимых тканях
Физиологию возбудимых тканей изучает электрофизиология.
Электрофизиология –исследует электрические
Биоэлектрические явления в возбудимых тканях
Физиологию возбудимых тканей изучает электрофизиология. Электрофизиология –исследует электрические

Основные задачи электрофизиологии:
1. Изучение физических и химических механизмов электрических процессов в
Основные задачи электрофизиологии:
1. Изучение физических и химических механизмов электрических процессов в

В основе реакций организма лежит раздражимость - способность реагировать на действие
В основе реакций организма лежит раздражимость - способность реагировать на действие

Явления связанные с природным электричеством (молнии, статическое электричество, электрические рыбы и
Явления связанные с природным электричеством (молнии, статическое электричество, электрические рыбы и

Бенджамин Франклин (1706—1790)
Разработал (1750) теорию электрических явлений, согласно которой электричество
Бенджамин Франклин (1706—1790)
Разработал (1750) теорию электрических явлений, согласно которой электричество


Впервые искусственную электрическую искру получили от электрической машины, изобретённой Герике -
Впервые искусственную электрическую искру получили от электрической машины, изобретённой Герике -

Первые электрофизиологические опыты.
Луиджи Гальвани (1737-1798)
Он обратил внимание на сокращение препарата
Первые электрофизиологические опыты.
Луиджи Гальвани (1737-1798)
Он обратил внимание на сокращение препарата

Итальянский физиолог Луиджи Гальвани из Болоньи сделал важное наблюдение. Он заметил,
Итальянский физиолог Луиджи Гальвани из Болоньи сделал важное наблюдение. Он заметил,

Лаборатория Гальвани
Лаборатория Гальвани

Алессандро Вольта (1745-1827)
Вольта повторил опыт Гальвани, но объяснил, его результаты
Алессандро Вольта (1745-1827)
Вольта повторил опыт Гальвани, но объяснил, его результаты
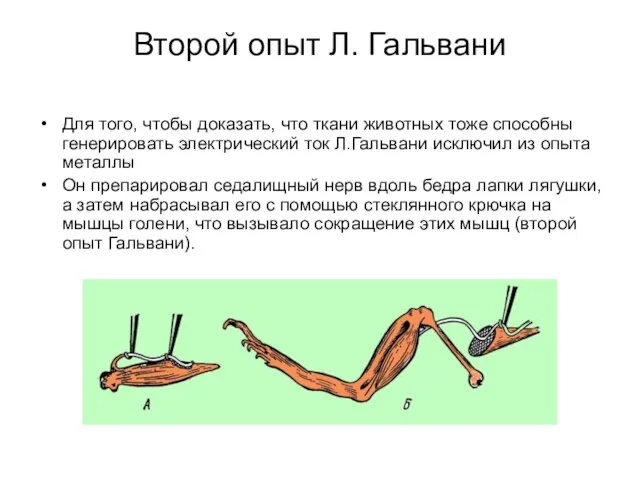
Второй опыт Л. Гальвани
Для того, чтобы доказать, что ткани животных тоже
Второй опыт Л. Гальвани
Для того, чтобы доказать, что ткани животных тоже

Первый «балконный» опыт Л.Гальвани (1786 г).
Гальвани повторил этот опыт в условиях
Первый «балконный» опыт Л.Гальвани (1786 г).
Гальвани повторил этот опыт в условиях

Триумф исследовательской программы Дж. Альдини произошел в Лондоне 17 января 1803
Триумф исследовательской программы Дж. Альдини произошел в Лондоне 17 января 1803

Дальнейшие исследования
В 1838 г. Матеуччи обнаружил, что можно зарегистрировать электрический ток
Дальнейшие исследования
В 1838 г. Матеуччи обнаружил, что можно зарегистрировать электрический ток

Что такое возбудимые ткани
Это ткани, клетки которых способны в ответ на
Что такое возбудимые ткани
Это ткани, клетки которых способны в ответ на

Все электрические процессы разворачиваются на цитоплазматической мембране. Клеточные мембраны состоят из
Все электрические процессы разворачиваются на цитоплазматической мембране. Клеточные мембраны состоят из

Белковые молекулы частично погружены в слой липидов, либо с внеклеточной стороны,
Белковые молекулы частично погружены в слой липидов, либо с внеклеточной стороны,

Функции клеточных мембран:
Барьерная функция – участие в механизмах электрогенеза, формирования ПП
Функции клеточных мембран:
Барьерная функция – участие в механизмах электрогенеза, формирования ПП

сома (тело) нейрона:
размер 5-100 мкм, разнообразие форм (пирамидная, звездчатая, грушевидная и
сома (тело) нейрона:
размер 5-100 мкм, разнообразие форм (пирамидная, звездчатая, грушевидная и

Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов –
Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов –

ПД – универсальный
ответ нервной клетки
на стимуляцию
20 мВ:
пороговый
стимул при
ПП= -70
ПД – универсальный
ответ нервной клетки
на стимуляцию
20 мВ:
пороговый
стимул при
ПП= -70

В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы

В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
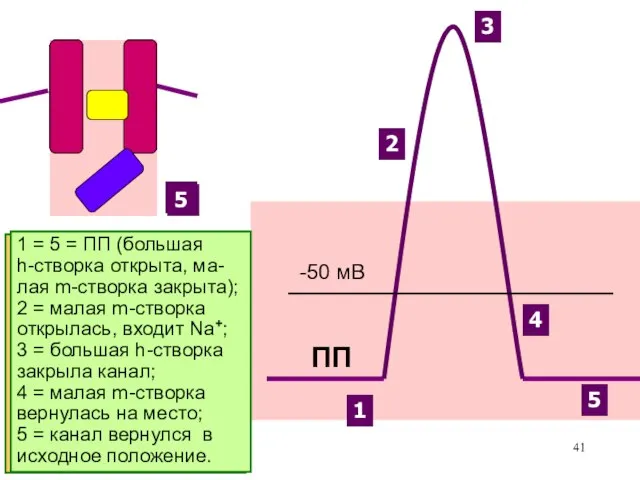
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем

Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем

Поскольку К+-каналы начинают закрываться довольно поздно (вслед за проходом уровня -50
Поскольку К+-каналы начинают закрываться довольно поздно (вслед за проходом уровня -50

тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекра
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекра

ТЭА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД
ТЭА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД

Проведение возбуждения в нервных волокнах
При распространении возбуждения по безмиелиновому нервному волокну
Проведение возбуждения в нервных волокнах
При распространении возбуждения по безмиелиновому нервному волокну

РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона

РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона

Скорость такого распространения низка и не пре-вышает у человека 1-2 м/с
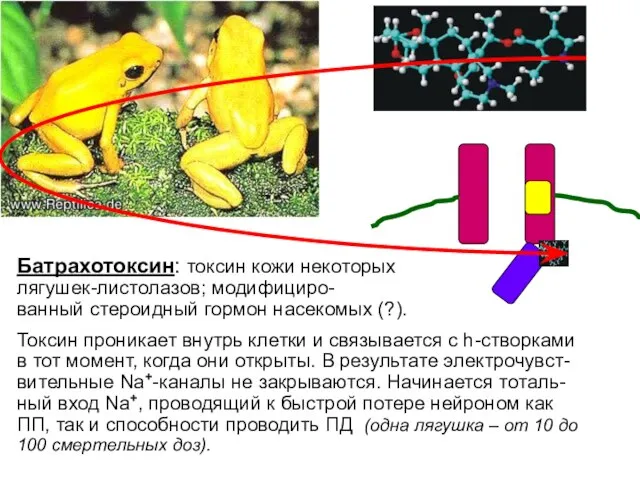
Батрахотоксин: токсин кожи некоторых
лягушек-листолазов; модифициро-
ванный стероидный гормон насекомых (?).
Токсин проникает
Батрахотоксин: токсин кожи некоторых
лягушек-листолазов; модифициро-
ванный стероидный гормон насекомых (?).
Токсин проникает
Мозг человека содержит ≈ 100 миллиардов нейронов.
Каждый нейрон образует контакты
Мозг человека содержит ≈ 100 миллиардов нейронов.
Каждый нейрон образует контакты

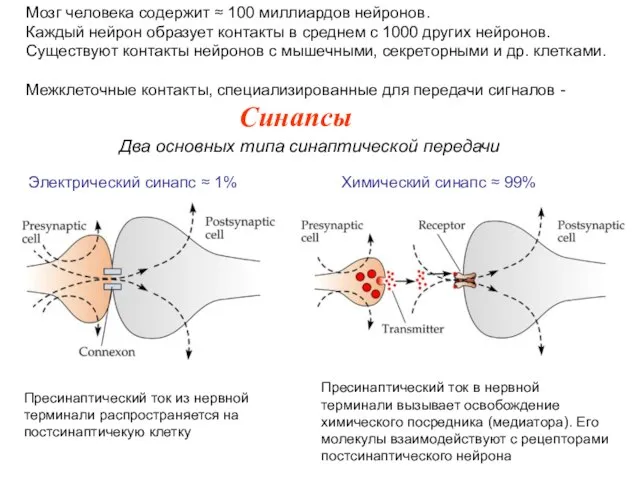
Структурные требования к электрическому механизму синаптической передачи
1.Тесное прилегание пре- и постсинаптических
Структурные требования к электрическому механизму синаптической передачи
1.Тесное прилегание пре- и постсинаптических
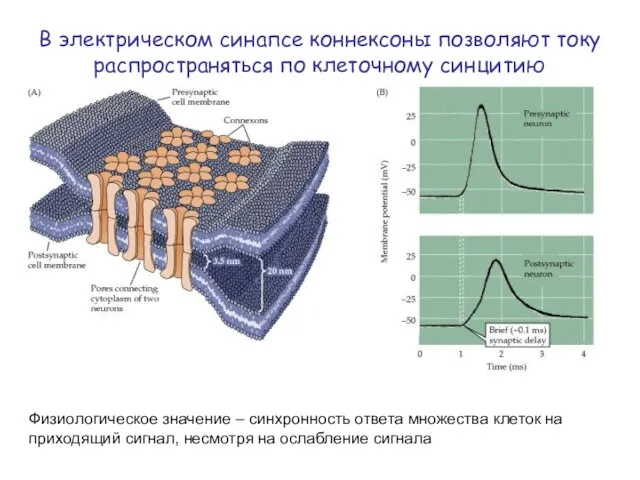
Физиологическое значение – синхронность ответа множества клеток на приходящий сигнал, несмотря
Физиологическое значение – синхронность ответа множества клеток на приходящий сигнал, несмотря

Менее распространены, чем химические (≈ 1%).
Прилегающие мембраны соединены щелевым контактом.
Ток течет
Менее распространены, чем химические (≈ 1%).
Прилегающие мембраны соединены щелевым контактом.
Ток течет

Структурные требования к химическому механизму синаптической передачи
Взаимодействие трех структур: (а) пресинаптической
Структурные требования к химическому механизму синаптической передачи
Взаимодействие трех структур: (а) пресинаптической

Медиаторы
Требования к молекулярным свойствам медиаторов
Высокая скорость диффузии, а значит низкий
Медиаторы
Требования к молекулярным свойствам медиаторов
Высокая скорость диффузии, а значит низкий

Электронная микроскопия химических синапсов
Пре-
Пре-
Пост-
Пост-
Видны:
пресинаптическая нервная терминаль, содержащая синаптические пузырьки, заполненные
Электронная микроскопия химических синапсов
Пре-
Пре-
Пост-
Пост-
Видны:
пресинаптическая нервная терминаль, содержащая синаптические пузырьки, заполненные

Значение колокализации Са2+ каналов и участков экзоцитоза
Короткая дистанция – открытие одного
Значение колокализации Са2+ каналов и участков экзоцитоза
Короткая дистанция – открытие одного

Экзоцитоз содержимого синаптических пузырьков
Непременными условиями экзоцитоза являются:(i) контакт синаптического пузырька с
Экзоцитоз содержимого синаптических пузырьков
Непременными условиями экзоцитоза являются:(i) контакт синаптического пузырька с

Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 1.
Основные белки
Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 1.
Основные белки
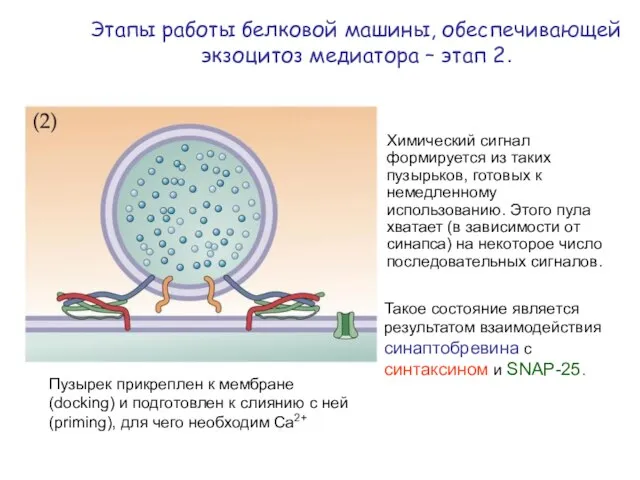
Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 2.
Пузырек прикреплен
Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 2.
Пузырек прикреплен

Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 3
Са2+ зависимый
Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 3
Са2+ зависимый

Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 4
Финал. Произошло
Этапы работы белковой машины, обеспечивающей экзоцитоз медиатора – этап 4
Финал. Произошло

Разнообразие морфологии нейронов
Разнообразие морфологии нейронов
 Механизмы вдоха и выдоха. Регуляция дыхания
Механизмы вдоха и выдоха. Регуляция дыхания Семеноводство зерновых культур
Семеноводство зерновых культур Класс Головоногие моллюски
Класс Головоногие моллюски Видоизменение побегов. Лабораторная работа Учитель биологии ГБОУ СОШ № 1022 Криулина И.В.
Видоизменение побегов. Лабораторная работа Учитель биологии ГБОУ СОШ № 1022 Криулина И.В.  Подкласс Ламииды Презентацию подготовил студент IV курса Ходыка Алексей
Подкласс Ламииды Презентацию подготовил студент IV курса Ходыка Алексей Класс круглоротые (cyclostomata). Систематика. Общая характеристика
Класс круглоротые (cyclostomata). Систематика. Общая характеристика Киндиойская восковая пальма
Киндиойская восковая пальма Деревья. Лес наше богатство. (Начальные классы)
Деревья. Лес наше богатство. (Начальные классы)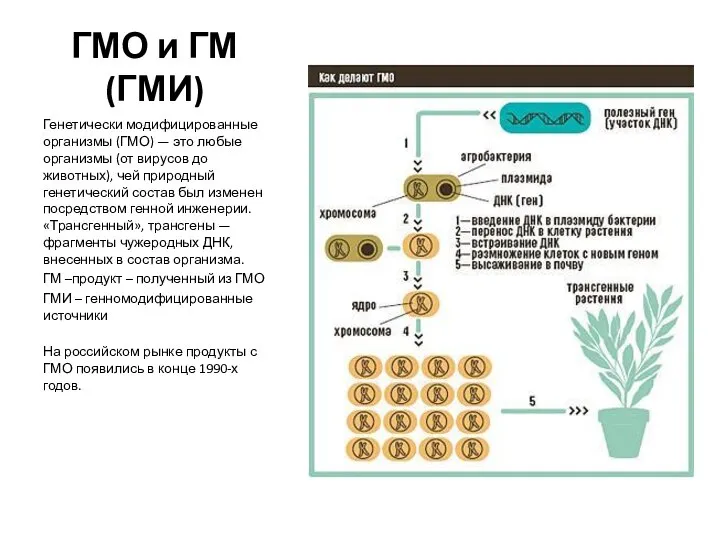 ГМО и ГМ (ГМИ) Генетически модифицированные организмы (ГМО) — это любые организмы (от вирусов до животных), чей природный генети
ГМО и ГМ (ГМИ) Генетически модифицированные организмы (ГМО) — это любые организмы (от вирусов до животных), чей природный генети Физиология анализаторов
Физиология анализаторов Насекомые
Насекомые Тварини - рекордсмени
Тварини - рекордсмени Мейоз. Изучение стадий мейоза на готовых микропрепаратах
Мейоз. Изучение стадий мейоза на готовых микропрепаратах Возникновение и развитие жизни на Земле
Возникновение и развитие жизни на Земле Клеточное строение корня
Клеточное строение корня Самые красивые породы короткошерстных кошек Короткошерстные кошки существовали задолго до длинношерстных их собратьев. Они б
Самые красивые породы короткошерстных кошек Короткошерстные кошки существовали задолго до длинношерстных их собратьев. Они б Презентация по биологии Мочекаменная болезнь
Презентация по биологии Мочекаменная болезнь  Причуды паразитизма
Причуды паразитизма Декоративные растения
Декоративные растения Насекомые
Насекомые Презентация на тему "История развития экологических связей человечества. Древние гоминиды" - скачать презентации по Биологи
Презентация на тему "История развития экологических связей человечества. Древние гоминиды" - скачать презентации по Биологи Общая характеристика периферического отдела скелета. Онто и филогенез
Общая характеристика периферического отдела скелета. Онто и филогенез Преданный друг
Преданный друг Своя игра по биологии
Своя игра по биологии Класс Млекопитающие. Внешнее строение, образ жизни и среда обитания млекопитающих животных.
Класс Млекопитающие. Внешнее строение, образ жизни и среда обитания млекопитающих животных. Химический состав клеток
Химический состав клеток organ_zrenia
organ_zrenia Организм и его свойства МОУ «МИРНЕНСКАЯ СОШ» Уйского района Челябинской области. Учитель биологии: Секерина Н.А.
Организм и его свойства МОУ «МИРНЕНСКАЯ СОШ» Уйского района Челябинской области. Учитель биологии: Секерина Н.А.