- Метаболизм ДНК

Содержание
- 2. Метаболизм ДНК Репликация – процесс точного копирования молекулы ДНК. Репарация – поддержание целостности биологического материала в
- 3. Репликация Процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетического материала и передачу его от поколения
- 4. Репликация ДНК происходит в определенной фазе клеточного цикла. Основные свойства процесса репликации ДНК и каталитические механизмы
- 5. Основные принципы репликации ДНК Полуконсервативный механизм - каждая из 2 цепей ДНК служит матрицей для образования
- 7. Субстратами для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты – dАТФ, dГТФ, dТТФ, dЦТФ, выполняющие роль строительного материала и
- 8. Точка начала репликации – origin (ориджин). У бактерий в кольцевом геноме имеется только одна точка «origin»,
- 10. Репликон – это участок ДНК между двумя «ориджинами» репликации.
- 11. Родительская ДНК Репликационный глазок Дочерние молекулы ДНК Репликативные вилки
- 12. Репликация идет в двух направлениях
- 13. Синтез новых цепей ДНК может протекать только в направлении 5’ → 3’, так как достраивается 3’-конец
- 14. 5’ 3’ 3’ 5’
- 15. Синтез ДНК
- 16. Вся сложность ферментативного аппарата репликации связана с требованиями высокой точности синтеза. Весь этот комплекс называется ДНК-репликативной
- 17. Ферменты репликации Синтез новой цепи ДНК осуществляется при помощи ДНК-полимеразы. Фермент катализирует синтез дочерних цепей на
- 18. Синтез ДНК У прокариот найдено 3 типа этих ферментов: ДНК-полимераза I ДНК-полимераза II ДНК-полимераза III
- 19. ДНК-полимераза I выполняет функции проверки поддержания порядка во время репликации, репарации и рекомбинации. Обладает 5’ →
- 20. ДНК-полимераза II – участвует в процессах репарации ДНК при повреждении ДНК ультрафиолетовым облучением.
- 21. ДНК-полимераза III – главный фермент репликации у E.coli. Состоит из субъединиц 10 разных типов. Обладает полимеразной
- 22. Для создания Матрицы – одноцепочечной ДНК, необходимы хеликазы – ферменты, разделяющие цепи двухцепочечной ДНК на одинарные
- 23. Хеликазы – это ферменты, способные расплетать две комплементарные нити в ДНК с использованием энергии, полученной при
- 24. Топоизомеразы – ферменты, изменяющие степень сверхспирализации ДНК, путем внесения одноцепочечных или двухцепочечных разрывов в ДНК.
- 25. Топоизомеразы находятся перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее расплетания и раскручивания молекулы ДНК,
- 26. Антибиотики – ингибиторы топоизомеразы: Хинолоны, фторхинолоны Норфлоксацин
- 27. белки, стабилизирующие разделенные нити ДНК – SSB (single strand binding)
- 28. Роль SSB -белков заключается в том, что они связываются с однонитчатой ДНК, выпрямляют ее и блокируют
- 29. ДНК-лигаза – фермент катализирующий сшивание одноцепочечных фрагментов ДНК. ДНК-праймаза — это фермент РНК-полимераза, синтезирующий короткий фрагмент
- 30. Синтез ДНК Синтез ДНК включает в себя три этапа: инициация элонгация терминация
- 31. Инициация инициация синтеза ДНК у E.coli происходит в oriC (245п.н.) В точках начала репликации отмечено большое
- 32. Процесс инициации начинается с присоединения к хромосоме белка DnaA (в origin области). 8 молекул DnaA, каждая
- 33. DnaА белки Dna A белки садятся на oriC – участок начала репликации Индуцируется расплетение ДНК Расплетенный
- 34. Это приводит к разделению цепей и способствует работе основного расплетающего белка - хеликазы (DnaB). С образовавшейся
- 35. Хеликазы Хеликазы разделяют ДНК в двух направлениях в районе двух вилок вилка вилка
- 37. Синтез ДНК В результате действия хеликазы, топоизомеразы, SSB- белков, ионов Mg+2 образуется репликативная вилка - участок
- 38. Синтез ДНК ДНК-полимераза III не способна начинать синтез новой цепи с ее первого нуклеотида. Поэтому репликация
- 39. Синтез ДНК Праймер - короткий сегмент РНК, комплементарный матричной цепи ДНК. Праймер синтезируется при участии ДНК-
- 40. Синтез ДНК от 3’ –конца праймера начинается синтез новой цепи ДНК при помощи ДНК- полимераза III.
- 41. Синтез ДНК ДНК-полимераза III удлиняет РНК-затравку, присоединяя к ней один за другим нуклеотиды, комплементарные матричной цепи
- 42. Элонгация Синтез лидирующей цепи. Синтез отстающей цепи.
- 43. Синтез лидирующей цепи начинается с синтеза праймера, дезоксирибонуклеотиды добавляются к этому праймеру ДНК-полимеразой III, связанной с
- 44. Синтез отстающей цепи Синтез осуществляется в виде коротких фрагментов ОКАЗАКИ. Каждый фрагмент Оказаки состоит примерно из
- 45. ДНК-полимераза III связывается с праймером и присоединяет дезоксирибонуклеотиды. Синтез фрагментов Оказаки происходит с участием ферментативного аппарата
- 46. Реплисома присоединяет по 1000 нуклеотидов в секунду к каждой цепи (лидирующей и отстающей). После завершения сборки
- 47. Оставшийся разрыв «сшивает» ДНК-лигаза. ДНК-лигаза катализирует образование фосфодиэфирной связи между 3’-гидроксильной группой на конце одной цепи
- 49. Терминация У прокариот есть специальные терминаторы (ter) – специальные последовательности нуклеотидов, прекращающие синтез цепи ДНК Ter-последовательности
- 50. Комплекс Ter-Tus может задержать репликативную вилку, движущуюся только в одном направлении, т.е. останавливает одну из вилок
- 51. Для терминации репликации линейных эукариотических хромосом на концах каждой хромосомы синтезируются специальные структуры, называемые теломерами. Иначе
- 52. Это многократно повторяющиеся последовательности нуклеотидов ( у одноклеточных эукариот от 20 до100, у млекопитающих превышает 1500).
- 53. Теломеры присоединяются к концам эукариотических хромосом с помощью фермента теломеразы.
- 54. Одна из причин старения — закон делимости клеток, открытый американским биологом Леонардом Хейфликом (Leonard Hayflick) в
- 55. есть клетки, которые могут делиться бесконечно, например кроветворные или раковые. В этих случаях механизм ограничения количества
- 56. Источники повреждения ДНК УФ излучение Радиация Химические вещества Ошибки репликации ДНК Апуринизация - отщепление азотистых оснований
- 57. Репарация ДНК Репарация- процесс восстановления повреждений ДНК. Распознавание дефекта сопровождается непосредственно при репликации, все ДНК-полимеразы обладают
- 58. Репарация осуществляется с помощью: специфического набора ферментов, постоянно присутствующих в нормально функционирующих клетках (фотореактивационная, эксцизионная) активации
- 59. У бактерий имеются 2 ферментные системы, ведущие репарацию: прямая эксцизионная
- 60. Прямая репарация ДНК Фотореактивация. Расщепление пиримидиновых димеров (приУФ облучении) осуществляется ферментом ДНК - фотолиазой. Реакция расщепления
- 61. фермент ДНК – фотолиаза - мономерный флавин-зависимый фермент и 2 кофактора (хромофоры). 5,10-метенилтетрагидрофолат (5,10-MTГФ) –поглощает фотоны
- 62. Репарация ДНК
- 63. Репарация ДНК Темновая эксцизионная репарация Не нуждается в энергии видимого света
- 64. Каждая из систем репарации включает следующие компоненты: фермент, «узнающий» химически изменённые участки в цепи ДНК и
- 65. Эксцизионная репарация Base excision repair – BER ДНК гликозилазы, распознают аномальные основания ДНК и катализируют гидролитическое
- 66. Фосфодиэстераза отщепляет от ДНК сахарофосфатную группу, к которой не присоединено основание. Брешь размеров в 1 н.
- 67. Nucleotide excision repair - NER Узнавание повреждений. Связывание мультисубъединичного комплекса с поврежденным сайтом. Двойное надрезание поврежденной
- 68. Освобождение олигонуклеотида, содержащего повреждение между двумя надрезами. Заполнение образовавшейся бреши ДНК полимеразой. Лигирование.
- 69. При некоторых типах повреждений ДНК (двухнитевые разрывы,поперечные сшивки). Репликативная вилка наталкивается на нерепарированное повреждение ДНК. Результат
- 70. SOS-репарация SOS – белки всегда присутствуют в клетке, но при запуске SOS-ответа их уровень значительно повышается.
- 71. В геноме типичной клетки млекопитающих за 24 ч аккумулируется много тысяч повреждений. Благодаря репарации менее одного
- 72. Все дефекты генов белков, участвующих в эксцизионной репарации связаны с онкологическими заболеваниями, генетическими заболеваниями, например, пигментная
- 73. У человека нет ДНК-фотолиазы и эксцизионная репарация оснований – единственный способ репарации пиримидиновых димеров. Инактивация этой
- 74. Молекулярные мутации Анемия Фанкони – генетические отклонения, возникающие при репарации ДНК. Болезнь названа в честь швейцарского
- 75. У 60—75 % больных также встречаются врожденные дефекты, такие как низкорослость, ненормальная пигментация, маленькая голова, аномалии
- 77. Скачать презентацию

Метаболизм ДНК
Репликация – процесс точного копирования молекулы ДНК.
Репарация – поддержание целостности
Метаболизм ДНК
Репликация – процесс точного копирования молекулы ДНК.
Репарация – поддержание целостности

Репликация
Процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетического материала и
Репликация
Процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетического материала и

Репликация ДНК происходит в определенной фазе клеточного цикла.
Основные свойства процесса репликации
Репликация ДНК происходит в определенной фазе клеточного цикла.
Основные свойства процесса репликации

Основные принципы репликации ДНК
Полуконсервативный механизм - каждая из 2 цепей ДНК
Основные принципы репликации ДНК
Полуконсервативный механизм - каждая из 2 цепей ДНК


Субстратами для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты – dАТФ, dГТФ, dТТФ, dЦТФ,
Субстратами для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты – dАТФ, dГТФ, dТТФ, dЦТФ,

Точка начала репликации – origin (ориджин).
У бактерий в кольцевом геноме имеется
Точка начала репликации – origin (ориджин).
У бактерий в кольцевом геноме имеется


Репликон – это участок ДНК между двумя «ориджинами» репликации.
Репликон – это участок ДНК между двумя «ориджинами» репликации.

Родительская ДНК
Репликационный глазок
Дочерние молекулы ДНК
Репликативные вилки
Родительская ДНК
Репликационный глазок
Дочерние молекулы ДНК
Репликативные вилки

Репликация идет в двух направлениях
Репликация идет в двух направлениях

Синтез новых цепей ДНК может протекать только в направлении
5’ →
Синтез новых цепей ДНК может протекать только в направлении
5’ →

5’
3’
3’
5’
5’
3’
3’
5’
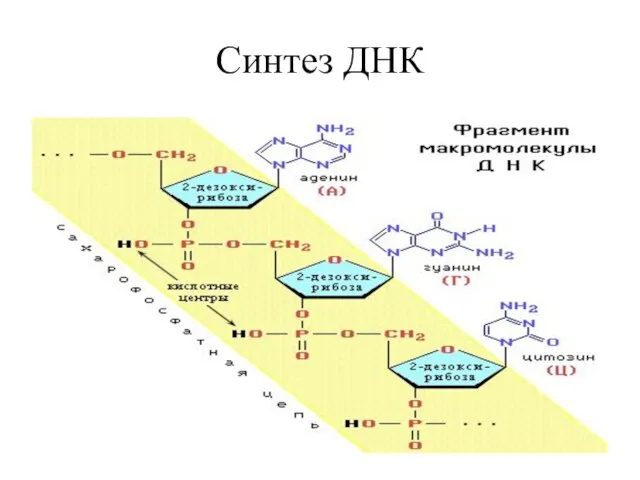
Синтез ДНК
Синтез ДНК

Вся сложность ферментативного аппарата репликации связана с требованиями высокой точности синтеза.
Вся сложность ферментативного аппарата репликации связана с требованиями высокой точности синтеза.

Ферменты репликации
Синтез новой цепи ДНК осуществляется при помощи ДНК-полимеразы.
Фермент катализирует
Ферменты репликации
Синтез новой цепи ДНК осуществляется при помощи ДНК-полимеразы.
Фермент катализирует

Синтез ДНК
У прокариот найдено 3 типа этих ферментов:
ДНК-полимераза I
Синтез ДНК
У прокариот найдено 3 типа этих ферментов:
ДНК-полимераза I

ДНК-полимераза I выполняет функции проверки поддержания порядка во время репликации, репарации
ДНК-полимераза I выполняет функции проверки поддержания порядка во время репликации, репарации

ДНК-полимераза II – участвует в процессах репарации ДНК при повреждении ДНК
ДНК-полимераза II – участвует в процессах репарации ДНК при повреждении ДНК

ДНК-полимераза III – главный фермент репликации у E.coli.
Состоит из субъединиц 10
ДНК-полимераза III – главный фермент репликации у E.coli.
Состоит из субъединиц 10

Для создания Матрицы – одноцепочечной ДНК, необходимы хеликазы – ферменты, разделяющие
Для создания Матрицы – одноцепочечной ДНК, необходимы хеликазы – ферменты, разделяющие

Хеликазы – это ферменты, способные расплетать две комплементарные нити в ДНК
Хеликазы – это ферменты, способные расплетать две комплементарные нити в ДНК

Топоизомеразы – ферменты, изменяющие степень сверхспирализации ДНК, путем внесения одноцепочечных
Топоизомеразы – ферменты, изменяющие степень сверхспирализации ДНК, путем внесения одноцепочечных

Топоизомеразы находятся перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее
Топоизомеразы находятся перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее

Антибиотики – ингибиторы топоизомеразы:
Хинолоны, фторхинолоны
Норфлоксацин
Антибиотики – ингибиторы топоизомеразы:
Хинолоны, фторхинолоны
Норфлоксацин

белки, стабилизирующие разделенные нити ДНК – SSB (single strand binding)
белки, стабилизирующие разделенные нити ДНК – SSB (single strand binding)

Роль SSB -белков заключается в том, что они связываются с однонитчатой
Роль SSB -белков заключается в том, что они связываются с однонитчатой

ДНК-лигаза – фермент катализирующий сшивание одноцепочечных фрагментов ДНК.
ДНК-праймаза — это
ДНК-лигаза – фермент катализирующий сшивание одноцепочечных фрагментов ДНК.
ДНК-праймаза — это

Синтез ДНК
Синтез ДНК включает в себя три этапа:
инициация
элонгация
терминация
Синтез ДНК
Синтез ДНК включает в себя три этапа:
инициация
элонгация
терминация

Инициация
инициация синтеза ДНК у E.coli происходит в oriC (245п.н.)
В точках начала
Инициация
инициация синтеза ДНК у E.coli происходит в oriC (245п.н.)
В точках начала

Процесс инициации начинается с присоединения к хромосоме белка DnaA (в origin
Процесс инициации начинается с присоединения к хромосоме белка DnaA (в origin
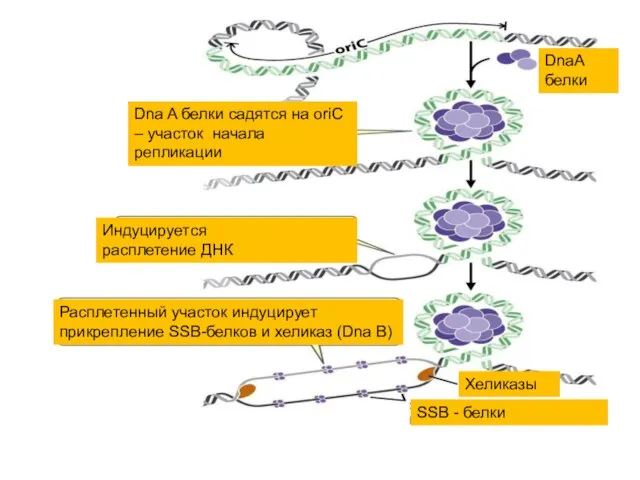
DnaА белки
Dna A белки садятся на oriC – участок начала репликации
Индуцируется
DnaА белки
Dna A белки садятся на oriC – участок начала репликации
Индуцируется

Это приводит к разделению цепей и способствует работе основного расплетающего белка
Это приводит к разделению цепей и способствует работе основного расплетающего белка
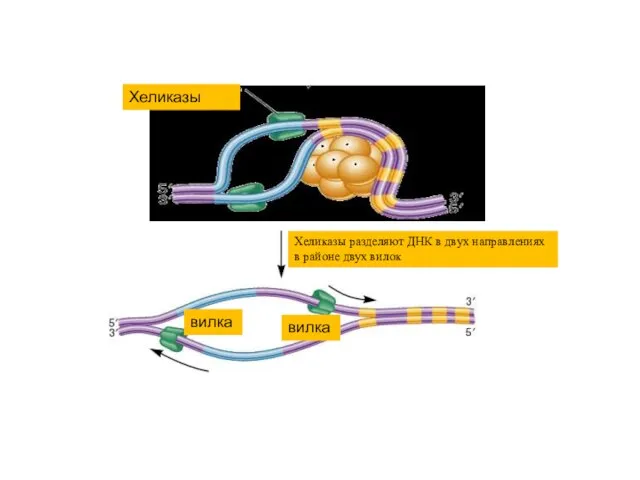
Хеликазы
Хеликазы разделяют ДНК в двух направлениях в районе двух вилок
вилка
вилка
Хеликазы
Хеликазы разделяют ДНК в двух направлениях в районе двух вилок
вилка
вилка


Синтез ДНК
В результате действия хеликазы, топоизомеразы, SSB- белков, ионов Mg+2
Синтез ДНК
В результате действия хеликазы, топоизомеразы, SSB- белков, ионов Mg+2

Синтез ДНК
ДНК-полимераза III не способна начинать синтез новой цепи с ее
Синтез ДНК
ДНК-полимераза III не способна начинать синтез новой цепи с ее

Синтез ДНК
Праймер - короткий сегмент РНК, комплементарный матричной цепи ДНК.
Праймер
Синтез ДНК
Праймер - короткий сегмент РНК, комплементарный матричной цепи ДНК.
Праймер

Синтез ДНК
от 3’ –конца праймера начинается синтез новой цепи ДНК при
Синтез ДНК
от 3’ –конца праймера начинается синтез новой цепи ДНК при

Синтез ДНК
ДНК-полимераза III удлиняет РНК-затравку, присоединяя к ней один за другим
Синтез ДНК
ДНК-полимераза III удлиняет РНК-затравку, присоединяя к ней один за другим

Элонгация
Синтез лидирующей цепи.
Синтез отстающей цепи.
Элонгация
Синтез лидирующей цепи.
Синтез отстающей цепи.

Синтез лидирующей цепи начинается с синтеза праймера, дезоксирибонуклеотиды добавляются к этому
Синтез лидирующей цепи начинается с синтеза праймера, дезоксирибонуклеотиды добавляются к этому

Синтез отстающей цепи
Синтез осуществляется в виде коротких фрагментов ОКАЗАКИ.
Каждый фрагмент Оказаки
Синтез отстающей цепи
Синтез осуществляется в виде коротких фрагментов ОКАЗАКИ.
Каждый фрагмент Оказаки

ДНК-полимераза III связывается с праймером и присоединяет дезоксирибонуклеотиды.
Синтез фрагментов Оказаки происходит
ДНК-полимераза III связывается с праймером и присоединяет дезоксирибонуклеотиды.
Синтез фрагментов Оказаки происходит

Реплисома присоединяет по 1000 нуклеотидов в секунду к каждой цепи (лидирующей
Реплисома присоединяет по 1000 нуклеотидов в секунду к каждой цепи (лидирующей

Оставшийся разрыв «сшивает» ДНК-лигаза.
ДНК-лигаза катализирует образование фосфодиэфирной связи между 3’-гидроксильной
Оставшийся разрыв «сшивает» ДНК-лигаза.
ДНК-лигаза катализирует образование фосфодиэфирной связи между 3’-гидроксильной


Терминация
У прокариот есть специальные терминаторы (ter) – специальные последовательности нуклеотидов, прекращающие
Терминация
У прокариот есть специальные терминаторы (ter) – специальные последовательности нуклеотидов, прекращающие

Комплекс Ter-Tus может задержать репликативную вилку, движущуюся только в одном направлении,
Комплекс Ter-Tus может задержать репликативную вилку, движущуюся только в одном направлении,

Для терминации репликации линейных эукариотических хромосом на концах каждой хромосомы синтезируются
Для терминации репликации линейных эукариотических хромосом на концах каждой хромосомы синтезируются

Это многократно повторяющиеся последовательности нуклеотидов ( у одноклеточных эукариот от 20
Это многократно повторяющиеся последовательности нуклеотидов ( у одноклеточных эукариот от 20

Теломеры присоединяются к концам эукариотических хромосом с помощью фермента теломеразы.
Теломеры присоединяются к концам эукариотических хромосом с помощью фермента теломеразы.

Одна из причин старения — закон делимости клеток, открытый американским биологом
Одна из причин старения — закон делимости клеток, открытый американским биологом

есть клетки, которые могут делиться бесконечно, например кроветворные или раковые. В
есть клетки, которые могут делиться бесконечно, например кроветворные или раковые. В

Источники повреждения ДНК
УФ излучение
Радиация
Химические вещества
Ошибки репликации ДНК
Апуринизация -
Источники повреждения ДНК
УФ излучение
Радиация
Химические вещества
Ошибки репликации ДНК
Апуринизация -

Репарация ДНК
Репарация- процесс восстановления повреждений ДНК.
Распознавание дефекта сопровождается непосредственно при репликации,
Репарация ДНК
Репарация- процесс восстановления повреждений ДНК.
Распознавание дефекта сопровождается непосредственно при репликации,

Репарация осуществляется с помощью: специфического набора ферментов, постоянно присутствующих в нормально
Репарация осуществляется с помощью: специфического набора ферментов, постоянно присутствующих в нормально

У бактерий имеются 2 ферментные системы, ведущие репарацию:
прямая
У бактерий имеются 2 ферментные системы, ведущие репарацию:
прямая

Прямая репарация ДНК
Фотореактивация. Расщепление пиримидиновых димеров (приУФ облучении) осуществляется ферментом
Прямая репарация ДНК
Фотореактивация. Расщепление пиримидиновых димеров (приУФ облучении) осуществляется ферментом

фермент ДНК – фотолиаза - мономерный флавин-зависимый фермент и 2
фермент ДНК – фотолиаза - мономерный флавин-зависимый фермент и 2
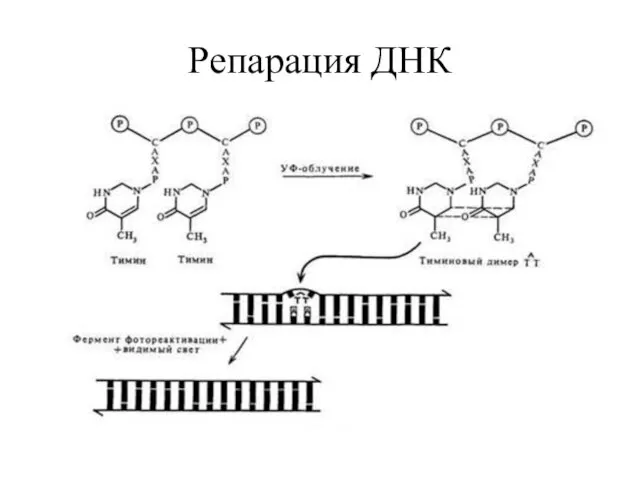
Репарация ДНК
Репарация ДНК

Репарация ДНК
Темновая эксцизионная репарация
Не нуждается в энергии видимого света
Репарация ДНК
Темновая эксцизионная репарация
Не нуждается в энергии видимого света

Каждая из систем репарации включает следующие компоненты:
фермент, «узнающий» химически изменённые участки
Каждая из систем репарации включает следующие компоненты:
фермент, «узнающий» химически изменённые участки

Эксцизионная репарация
Base excision repair – BER
ДНК гликозилазы, распознают аномальные основания ДНК
Эксцизионная репарация
Base excision repair – BER
ДНК гликозилазы, распознают аномальные основания ДНК

Фосфодиэстераза отщепляет от ДНК
сахарофосфатную группу, к которой не
присоединено основание.
Фосфодиэстераза отщепляет от ДНК
сахарофосфатную группу, к которой не
присоединено основание.

Nucleotide excision repair - NER
Узнавание повреждений.
Связывание мультисубъединичного комплекса с
Nucleotide excision repair - NER
Узнавание повреждений.
Связывание мультисубъединичного комплекса с

Освобождение олигонуклеотида, содержащего повреждение между двумя надрезами.
Заполнение образовавшейся бреши
Освобождение олигонуклеотида, содержащего повреждение между двумя надрезами.
Заполнение образовавшейся бреши

При некоторых типах повреждений ДНК (двухнитевые разрывы,поперечные сшивки).
Репликативная вилка наталкивается
При некоторых типах повреждений ДНК (двухнитевые разрывы,поперечные сшивки).
Репликативная вилка наталкивается

SOS-репарация
SOS – белки всегда присутствуют в клетке, но при запуске SOS-ответа
SOS-репарация
SOS – белки всегда присутствуют в клетке, но при запуске SOS-ответа

В геноме типичной клетки млекопитающих за 24 ч аккумулируется много тысяч
В геноме типичной клетки млекопитающих за 24 ч аккумулируется много тысяч

Все дефекты генов белков, участвующих в эксцизионной репарации связаны с онкологическими
Все дефекты генов белков, участвующих в эксцизионной репарации связаны с онкологическими

У человека нет ДНК-фотолиазы и эксцизионная репарация оснований – единственный способ
У человека нет ДНК-фотолиазы и эксцизионная репарация оснований – единственный способ

Молекулярные мутации
Анемия Фанкони – генетические отклонения, возникающие при репарации ДНК.
Болезнь
Молекулярные мутации
Анемия Фанкони – генетические отклонения, возникающие при репарации ДНК.
Болезнь

У 60—75 % больных также встречаются врожденные дефекты, такие как низкорослость, ненормальная
У 60—75 % больных также встречаются врожденные дефекты, такие как низкорослость, ненормальная
 Презентация на тему Виды и формы иммунитета
Презентация на тему Виды и формы иммунитета  Сезонные явления в жизни млекопитающих
Сезонные явления в жизни млекопитающих Сухие корма для кошек и собак
Сухие корма для кошек и собак Дыхательная система человека
Дыхательная система человека Презентация на тему "Скелет" - скачать бесплатно презентации по Биологии
Презентация на тему "Скелет" - скачать бесплатно презентации по Биологии Белый медведь
Белый медведь Дикие животные. Белка-летяга
Дикие животные. Белка-летяга НАРУШЕНИЯ ЗРЕНИЯ У ШКОЛЬНИКОВ Выполнила ученица 10 А класса: Чистопрудова Ольга Руководитель: учитель биологии Зенкова Наталья Ва
НАРУШЕНИЯ ЗРЕНИЯ У ШКОЛЬНИКОВ Выполнила ученица 10 А класса: Чистопрудова Ольга Руководитель: учитель биологии Зенкова Наталья Ва Гипофиз
Гипофиз Особенности высшей нервной деятельности человека. Речь. Мышление.труд. Познавательные процессы
Особенности высшей нервной деятельности человека. Речь. Мышление.труд. Познавательные процессы Органы чувств. Анализаторы
Органы чувств. Анализаторы День апельсиновых сказок
День апельсиновых сказок Презентация на тему "Индивидуальное развитие организма" - скачать презентации по Биологии
Презентация на тему "Индивидуальное развитие организма" - скачать презентации по Биологии Мхи. Сфагнум
Мхи. Сфагнум Размножение и развитие земноводных
Размножение и развитие земноводных Презентация на тему "Бабочка-капустница" - скачать презентации по Биологии
Презентация на тему "Бабочка-капустница" - скачать презентации по Биологии Презентация на тему Клеточный уровень жизни Изучить структуру клеток Механизм устойчивости клетки как биосистемы
Презентация на тему Клеточный уровень жизни Изучить структуру клеток Механизм устойчивости клетки как биосистемы  Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии
Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии Основные систематические группы Рыб
Основные систематические группы Рыб Механизмы инвазии вирусов в клетки
Механизмы инвазии вирусов в клетки Время посева и глубина заделки семян
Время посева и глубина заделки семян Марганец. Роль марганца в организме
Марганец. Роль марганца в организме Динозавры
Динозавры Презентация по биологии Слуховой анализатор. Гигиена слуха. 8 класс
Презентация по биологии Слуховой анализатор. Гигиена слуха. 8 класс Специфичность паразитов. Приуроченность определенных видов паразитов к определенным видам хозяев
Специфичность паразитов. Приуроченность определенных видов паразитов к определенным видам хозяев Белка-летяга
Белка-летяга Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Cигналинг в овулярной фазе. Диалог гаметофитов от А до Я
Cигналинг в овулярной фазе. Диалог гаметофитов от А до Я