- Cигналинг в овулярной фазе. Диалог гаметофитов от А до Я

Содержание
- 2. Сигнальные механизмы в пестике Преовулярные механизмы направляют рост пыльцевой трубки к завязи. Овулярные механизмы контролируют ее
- 3. ROP ГТФазы и рецепторные киназы В клетках растений основную роль играют мономерные (малые) G белки (20-30
- 4. Начальные стадии прорастания Активация пыльцы как in vitro, так и in vivo сопровождается активацией ROP ГТФаз
- 5. ROP1 В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована преимущественно в апикальной плазмалемме. Экспериментальный
- 6. Регуляция по принципу обратной связи Рецепторная киназуа RLK активирует фактор обмена нуклеотидов ROP GEF, ROP переводится
- 7. Эффекторные белки Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и контролирует накопление везикул, несущих
- 8. А что же АФК? Как мы помним, АФК контролируют 3 этапа прогамной фазы: собственно прорастание, рост
- 9. Ещё немного о Са2+ и АФК Фосфорилирование НАДФН-оксидазы Rboh придает ей способность связывать Са2+ Присоединение Са2+
- 10. Убегание от NO? YES Оксид азота содержится в пероксисомах ПТ и выделяется пыльцой, снижая концентрацию АФК
- 11. NO как сигнал от семяпочки Нацеливание трубки на микропиле происходит с помощью NO, который образуется в
- 12. NO и кальций При «убегании» трубок лилии от источника NO сначала концентрация Ca2+ возрастала, при этом
- 13. NО, актин и пектины
- 14. Teun Munnik, and Christa Testerink J. Lipid Res. 2009;50:S260-S265
- 15. Распределение фосфолипидов Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения актинового кольца. PI(4,5)P2 локализован
- 16. Фосфолипиды и актин PI(4,5)P2 может различными способами влиять на динамику актина и его связь с мембраной:
- 17. Фосфолипиды и везикулярный транспорт Экзогенная PA и ингибитор деградации PA стимулировали образование пучков актиновых филаментов, везикулярный
- 18. Итак, приближаемся к семяпочке Овулярный этап разделяют на две фазы регуляции роста пыльцевой трубки: фуникулярная и
- 19. Фуникулярные сигналы Природа фуникулярных сигналов, исходящих из семяпочки, неясна. В числе кандидатов рассматривают NO и D-серин,
- 20. GABA Ингибирование глутаматдекарбоксилазы – фермента, лимитирующего скорость биосинтеза GABA, – вызывало в пыльцевой трубке дезорганизацию актинового
- 21. Как действует GABA? Активирует вход кальция Регулирует экспрессию ряда генов, включая ГТФазы и ферменты липидного сигналинга
- 22. GABA в пестике В пестике табака обнаружен градиент концентрации GABA. Предполагается, что секретируемая пестиком GABA активирует
- 23. Микропилярные сигналы Эти сигналы имеют короткий радиус действия (100-200 мкм от микропиле), значит, здесь могут работать
- 24. Механика врастания В конце своего пути пыльцевая трубка замедляет рост и проникает в одну из синергид
- 25. Зона взаимодействия гамет Цитоплазмы ВК и С смешиваются Плазмалемма С разрушается, и спермии в составе MGU
- 26. Оплодотворение После разрыва трубки, скорость движения спермиев возрастает примерно в 100 раз. Неизвестно, используется ли на
- 27. Экспрессировали в спермиях флуоресцентный белок, способный к фотоконверсии: облучение УФ сдвигало спектр флуоресценции этого белка. Один
- 28. Одинаковые ли синергиды? Две синергиды в зрелом зародышевом мешке многих растений неразличимы, и выбор трубкой рецептивной
- 29. Нитчатый аппарат Нитчатый аппарат представляет собой систему извилистых и разветвленных впячиваний клеточной стенки в цитоплазму. Они
- 30. Сигнальные пептиды 2Д В числе первых аттрактантов, синтезируемых синергидами, были идентифицированы цистеин-богатые полипептиды LURE, обнаруженные у
- 31. А у однодольных? У кукурузы Z. Mays EGG APPARATUS 1 (ZmEA1). Радиус его аттрагирующего действия составляет
- 32. О том же, но красиво
- 33. Богиня плодородия Ключевую роль в разрыве трубки играют рецепторные киназы. Первая РК синергиды, названа в честь
- 34. Причем тут АФК? Итак, FERONIA должна вызвать разрыв пыльцевой трубки, она же должна прекратить аттракцию новых
- 35. Другие участники Два других белка синергиды, контролирующих поведение трубки в зародышевом мешке, – это NORTIA (NTA)
- 36. Красивые имена NORTIA и LORELEI Для NTA, как и для FER, характерна полярная локализация – в
- 37. LRE, как и FER, индуцирует образование АФК в нитчатом аппарате Популярная версия: LRE, наряду с FER,
- 38. Взаимодействие гамет. Оплодотворение Двойное оплодотворение у покрытосеменных растений было открыто С.Г. Навашиным в конце XIX в.
- 39. Гаметы и Са2+ (in vitro) При слиянии спермия и яйцеклетки кукурузы in vitro наблюдали временное увеличение
- 40. Кальциевая подпись in vivo
- 41. In vivo 2 пика Более сложная картина: 2 пика! Первый пик соответствует высвобождению спермиев. Значение этого
- 42. А что в синергидах? В синергиде при непосредственном контакте с ней апекса пыльцевой трубки возникали кальциевые
- 43. Центральная клетка В ЦК наблюдается один короткий пик, соответствующий выходу спермиев из трубки. выход пауза плазмогамия
- 44. Диалог гамет во время «паузы» GENERATIVE CELL SPECIFIC 1 (GCS1) у Lilium longiflorum и его ортолог
- 45. Адгезия и слияние Оказалось, что адгезия мембран и их слияние – разные процессы. Гаметы мутантных растений
- 46. На протяжении роста пыльцевой трубки Arabidopsis белок HAP2/GCS1 локализован в эндомембранной системе спермиев. Он выходит на
- 47. Активация гамет При этом из эндомембранной системы на поверхность спермия выходят белки HAP2/GCS1. Это происходит одновременно
- 48. Яйцеклетка: скромная или активная? В целом, слияние гамет у растений происходит почти так же, как у
- 49. Предотвращение полиспермии После высвобождения спермиев быстро прекращается аттракция новых ПТ. Если слияния гамет почему-либо не произошло,
- 50. Итак, слияние Каким образом ядро спермия транспортируется к ядру женской гаметы? В яйцеклетках животных эту функцию
- 51. Актин в ЦК Актин – основной участник движения ядер спермиев как в ЯК, так и в
- 52. Детали Ядро спермия, войдя в цитоплазму ЦК, окружалось волокнами F-актина с образованием звездообразной структуры. Вся эта
- 53. Кариогамия 1) наружные ядерные мембраны гамет соприкасаются и затем сливаются, 2) внутренние ядерные мембраны сливаются, и
- 54. В системе in vitro у риса путь ядра спермия до ядра яйцеклетки занимал 5-10 мин, причем,
- 55. САМОНЕСОВМЕСТИМОСТЬ Когда «родной» вовсе значит «желанный»
- 56. Два типа СН Гаметофитная СН: фенотип пыльцы определяется гаплоидным геномом (гаметофитным) Спорофитная СН: фенотип пыльцы определяется
- 57. Гаметофитная СН. S-RNAse Рост пыльцы подавляется только тогда, когда она вошла в проводниковый тракт столбика. Что
- 58. S-РНКаза Этот фермент деградирует рРНК в самонесовместимой пыльце При потере РНКазной активности СН не работает Гены
- 59. Две модели Думали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто не заходят Но оказалось,
- 60. Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому Своя SРНКаза узнаётся рецептором SLF (S-locus
- 61. Спорофитная СН у капусты Меняемся ролями: мужской пепитид – лиганд, женский – рецептор У капусты S-локус
- 62. Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein) Полиморфизм мужской детерминанты выше Она экспрессируется
- 65. Скачать презентацию
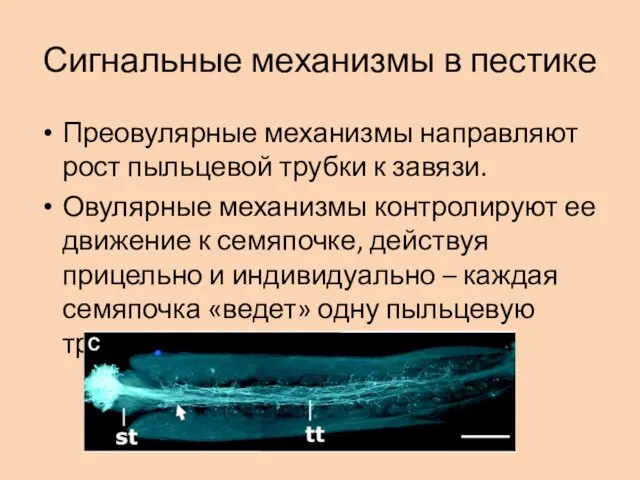
Сигнальные механизмы в пестике
Преовулярные механизмы направляют рост пыльцевой трубки к завязи.
Сигнальные механизмы в пестике
Преовулярные механизмы направляют рост пыльцевой трубки к завязи.

ROP ГТФазы и рецепторные киназы
В клетках растений основную роль играют мономерные
ROP ГТФазы и рецепторные киназы
В клетках растений основную роль играют мономерные

Начальные стадии прорастания
Активация пыльцы как in vitro, так и in vivo
Начальные стадии прорастания
Активация пыльцы как in vitro, так и in vivo

ROP1
В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована преимущественно
ROP1
В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована преимущественно
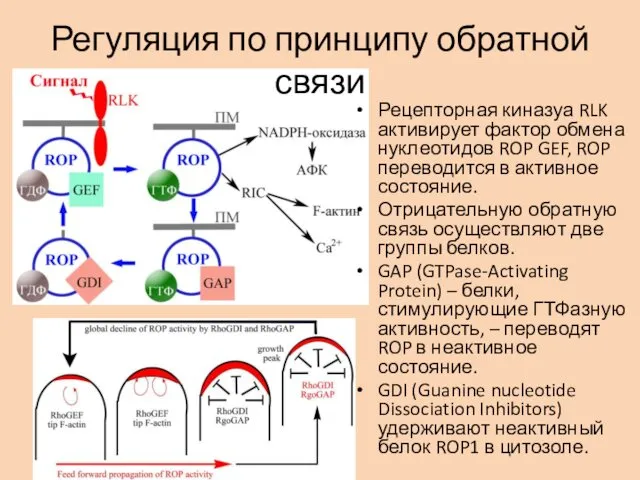
Регуляция по принципу обратной связи
Рецепторная киназуа RLK активирует фактор обмена нуклеотидов
Регуляция по принципу обратной связи
Рецепторная киназуа RLK активирует фактор обмена нуклеотидов
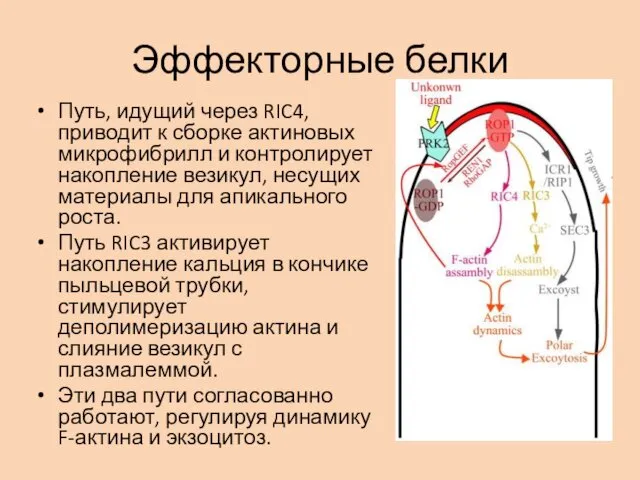
Эффекторные белки
Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и
Эффекторные белки
Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и

А что же АФК?
Как мы помним, АФК контролируют 3 этапа прогамной
А что же АФК?
Как мы помним, АФК контролируют 3 этапа прогамной
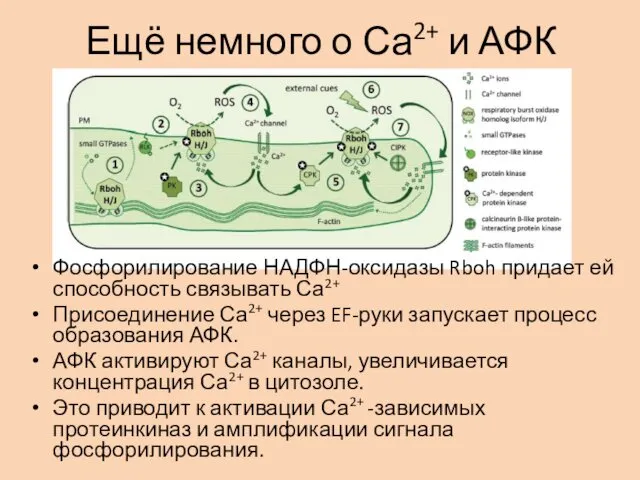
Ещё немного о Са2+ и АФК
Фосфорилирование НАДФН-оксидазы Rboh придает ей способность
Ещё немного о Са2+ и АФК
Фосфорилирование НАДФН-оксидазы Rboh придает ей способность
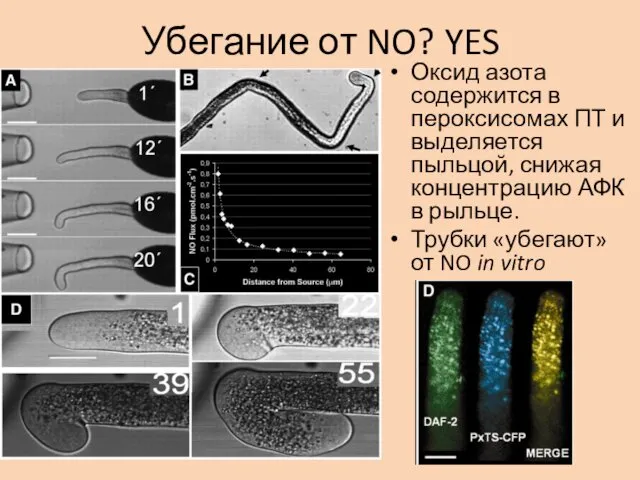
Убегание от NO? YES
Оксид азота содержится в пероксисомах ПТ и выделяется
Убегание от NO? YES
Оксид азота содержится в пероксисомах ПТ и выделяется
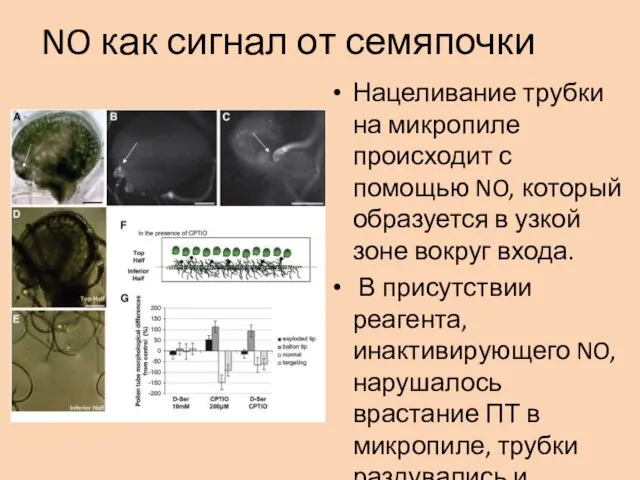
NO как сигнал от семяпочки
Нацеливание трубки на микропиле происходит с помощью
NO как сигнал от семяпочки
Нацеливание трубки на микропиле происходит с помощью
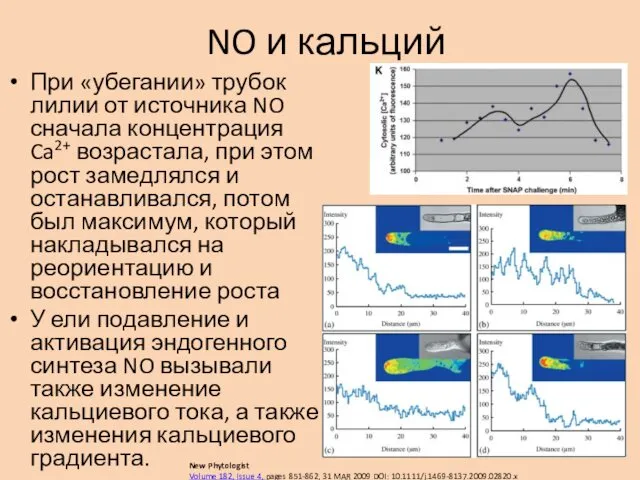
NO и кальций
При «убегании» трубок лилии от источника NO сначала концентрация
NO и кальций
При «убегании» трубок лилии от источника NO сначала концентрация
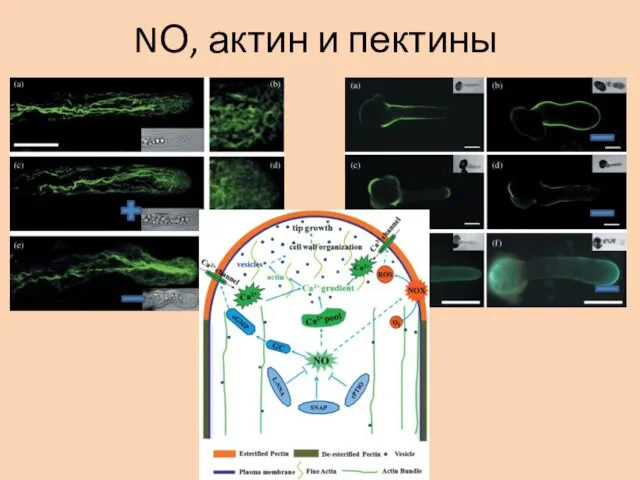
NО, актин и пектины
NО, актин и пектины
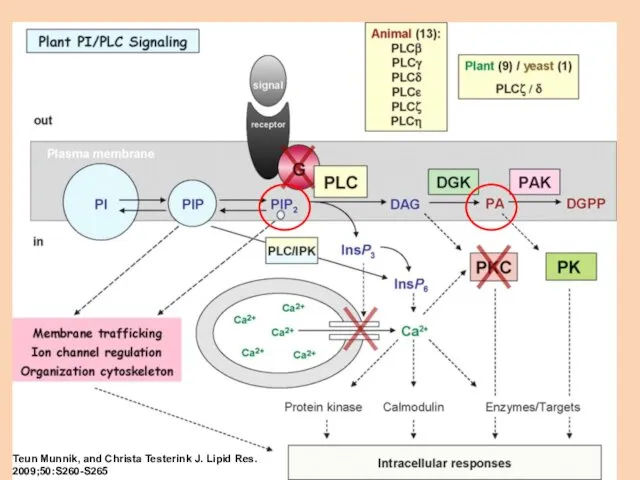
Teun Munnik, and Christa Testerink J. Lipid Res. 2009;50:S260-S265
Teun Munnik, and Christa Testerink J. Lipid Res. 2009;50:S260-S265
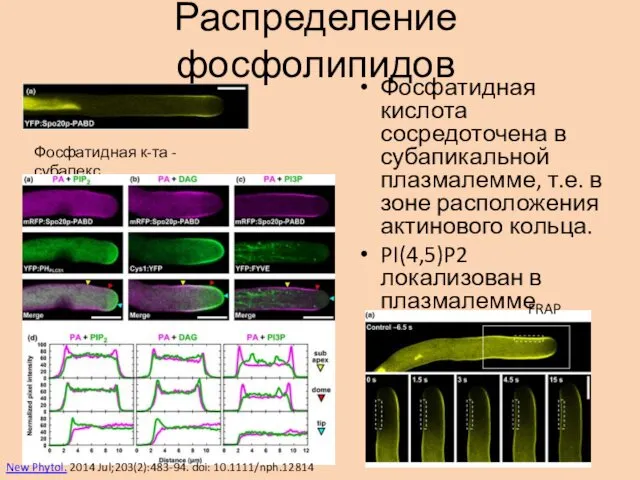
Распределение фосфолипидов
Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения
Распределение фосфолипидов
Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения
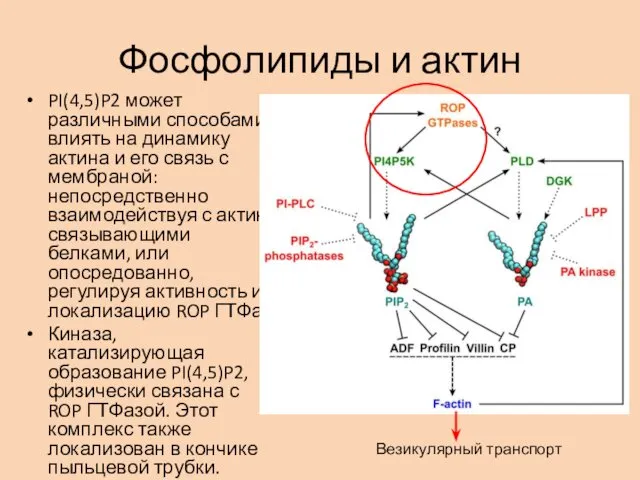
Фосфолипиды и актин
PI(4,5)P2 может различными способами влиять на динамику актина и
Фосфолипиды и актин
PI(4,5)P2 может различными способами влиять на динамику актина и
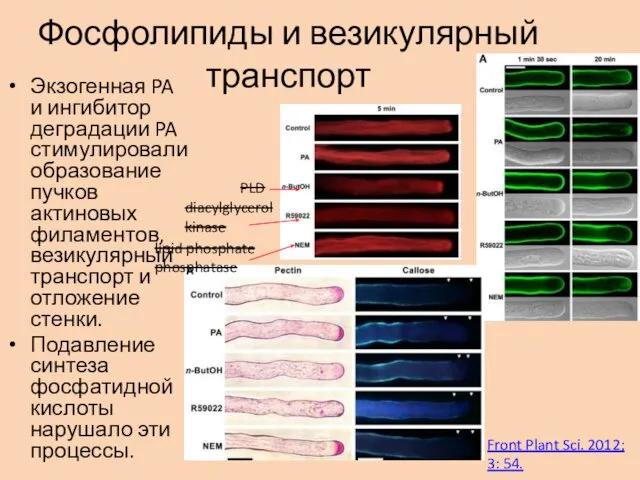
Фосфолипиды и везикулярный транспорт
Экзогенная PA и ингибитор деградации PA стимулировали образование
Фосфолипиды и везикулярный транспорт
Экзогенная PA и ингибитор деградации PA стимулировали образование

Итак, приближаемся к семяпочке
Овулярный этап разделяют на две фазы регуляции роста
Итак, приближаемся к семяпочке
Овулярный этап разделяют на две фазы регуляции роста

Фуникулярные сигналы
Природа фуникулярных сигналов, исходящих из семяпочки, неясна. В числе кандидатов
Фуникулярные сигналы
Природа фуникулярных сигналов, исходящих из семяпочки, неясна. В числе кандидатов
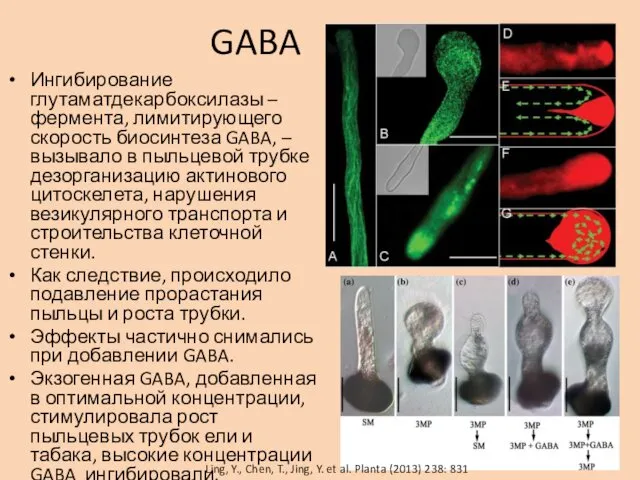
GABA
Ингибирование глутаматдекарбоксилазы – фермента, лимитирующего скорость биосинтеза GABA, – вызывало в
GABA
Ингибирование глутаматдекарбоксилазы – фермента, лимитирующего скорость биосинтеза GABA, – вызывало в
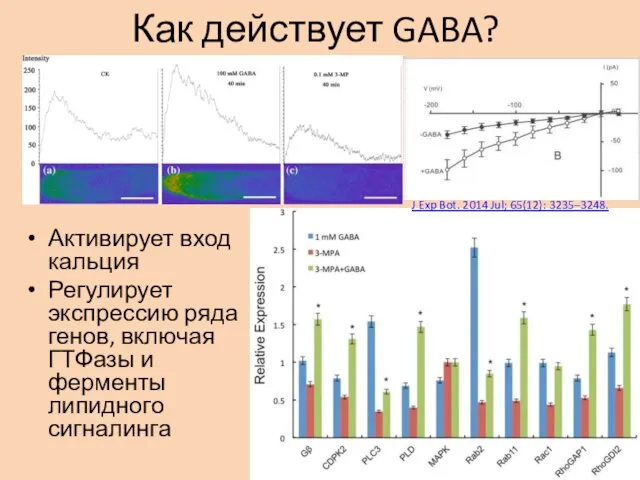
Как действует GABA?
Активирует вход кальция
Регулирует экспрессию ряда генов, включая ГТФазы
Как действует GABA?
Активирует вход кальция
Регулирует экспрессию ряда генов, включая ГТФазы
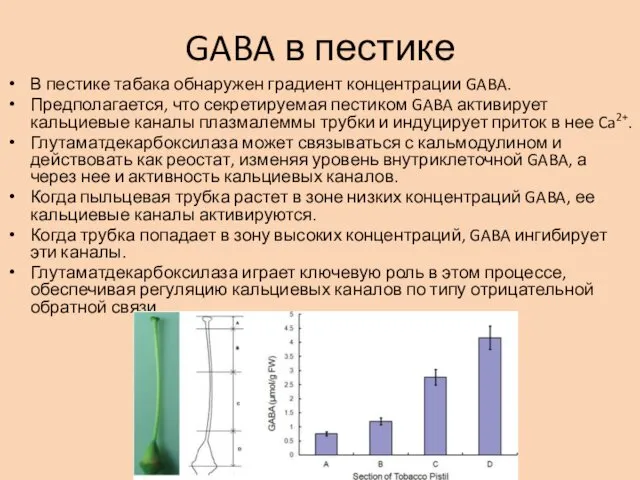
GABA в пестике
В пестике табака обнаружен градиент концентрации GABA.
Предполагается, что
GABA в пестике
В пестике табака обнаружен градиент концентрации GABA.
Предполагается, что

Микропилярные сигналы
Эти сигналы имеют короткий радиус действия (100-200 мкм от микропиле),
Микропилярные сигналы
Эти сигналы имеют короткий радиус действия (100-200 мкм от микропиле),
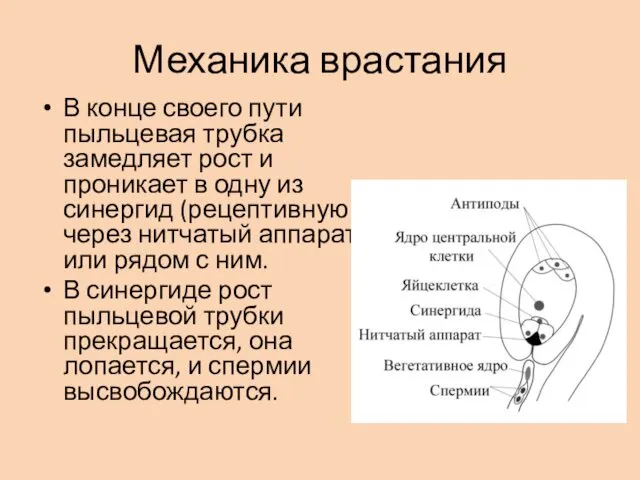
Механика врастания
В конце своего пути пыльцевая трубка замедляет рост и проникает
Механика врастания
В конце своего пути пыльцевая трубка замедляет рост и проникает
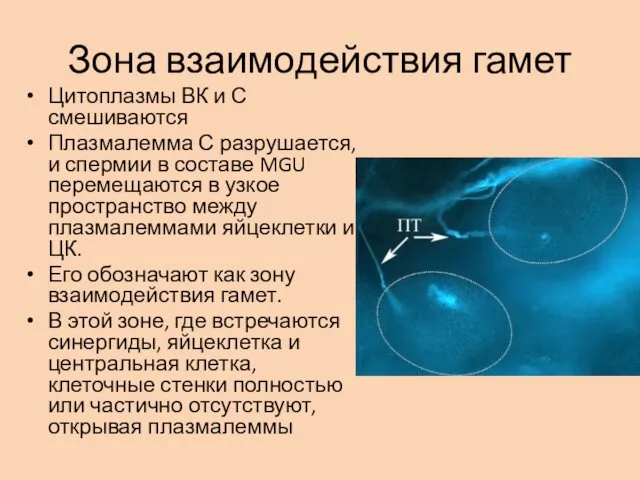
Зона взаимодействия гамет
Цитоплазмы ВК и С смешиваются
Плазмалемма С разрушается, и спермии
Зона взаимодействия гамет
Цитоплазмы ВК и С смешиваются
Плазмалемма С разрушается, и спермии

Оплодотворение
После разрыва трубки, скорость движения спермиев возрастает примерно в 100 раз.
Неизвестно,
Оплодотворение
После разрыва трубки, скорость движения спермиев возрастает примерно в 100 раз.
Неизвестно,
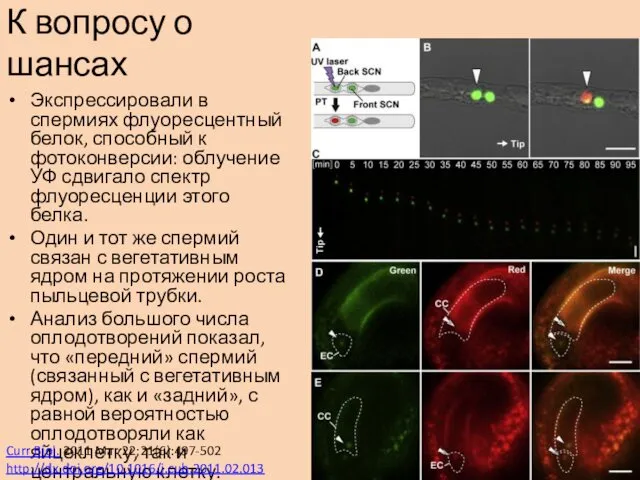
Экспрессировали в спермиях флуоресцентный белок, способный к фотоконверсии: облучение УФ сдвигало
Экспрессировали в спермиях флуоресцентный белок, способный к фотоконверсии: облучение УФ сдвигало

Одинаковые ли синергиды?
Две синергиды в зрелом зародышевом мешке многих растений неразличимы,
Одинаковые ли синергиды?
Две синергиды в зрелом зародышевом мешке многих растений неразличимы,
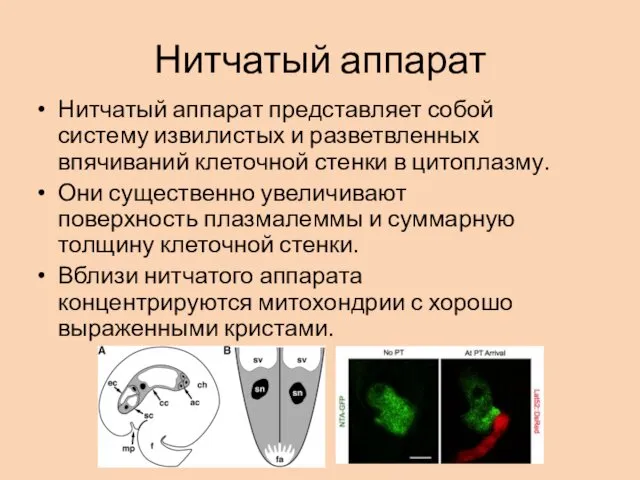
Нитчатый аппарат
Нитчатый аппарат представляет собой систему извилистых и разветвленных впячиваний клеточной
Нитчатый аппарат
Нитчатый аппарат представляет собой систему извилистых и разветвленных впячиваний клеточной
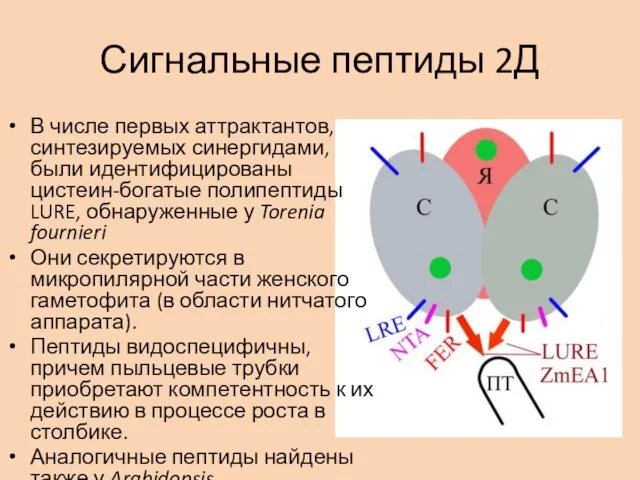
Сигнальные пептиды 2Д
В числе первых аттрактантов, синтезируемых синергидами, были идентифицированы цистеин-богатые
Сигнальные пептиды 2Д
В числе первых аттрактантов, синтезируемых синергидами, были идентифицированы цистеин-богатые
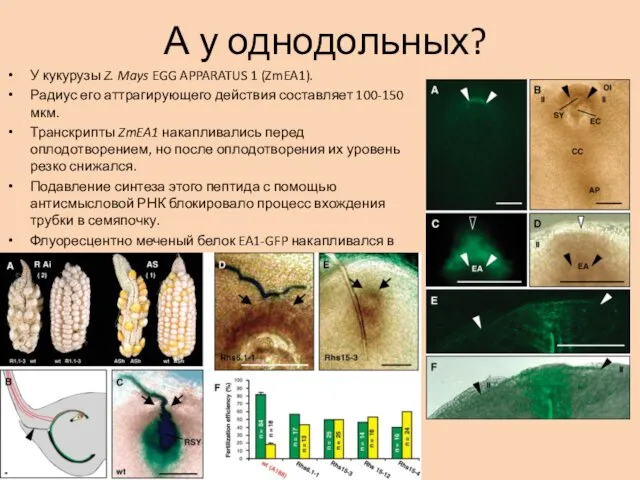
А у однодольных?
У кукурузы Z. Mays EGG APPARATUS 1 (ZmEA1).
Радиус
А у однодольных?
У кукурузы Z. Mays EGG APPARATUS 1 (ZmEA1).
Радиус
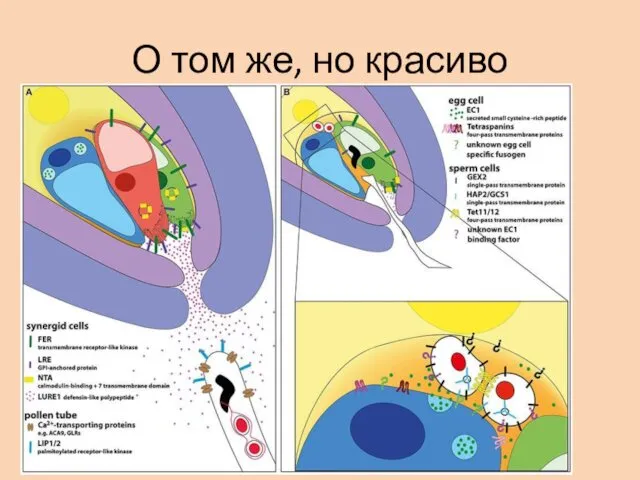
О том же, но красиво
О том же, но красиво
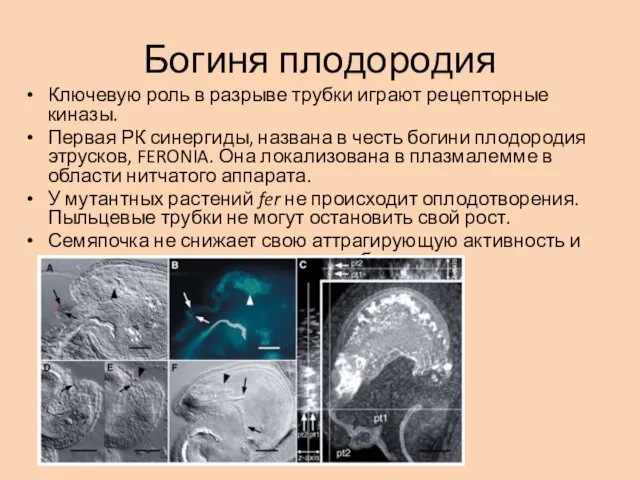
Богиня плодородия
Ключевую роль в разрыве трубки играют рецепторные киназы.
Первая РК
Богиня плодородия
Ключевую роль в разрыве трубки играют рецепторные киназы.
Первая РК
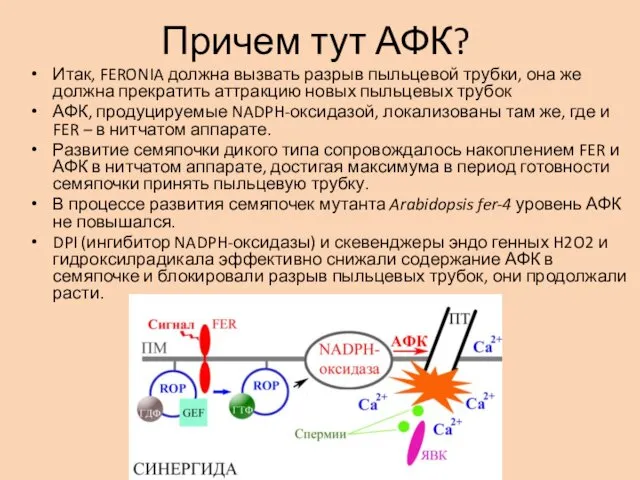
Причем тут АФК?
Итак, FERONIA должна вызвать разрыв пыльцевой трубки, она же
Причем тут АФК?
Итак, FERONIA должна вызвать разрыв пыльцевой трубки, она же
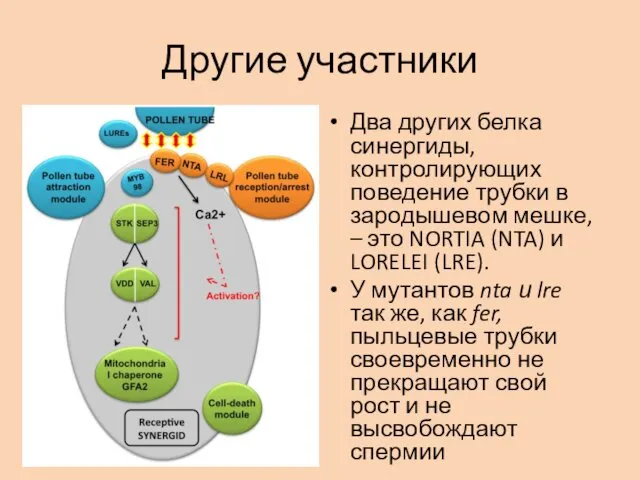
Другие участники
Два других белка синергиды, контролирующих поведение трубки в зародышевом мешке,
Другие участники
Два других белка синергиды, контролирующих поведение трубки в зародышевом мешке,
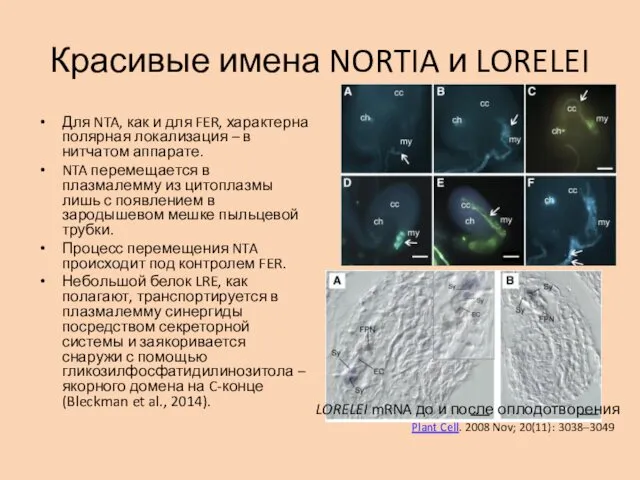
Красивые имена NORTIA и LORELEI
Для NTA, как и для FER, характерна
Красивые имена NORTIA и LORELEI
Для NTA, как и для FER, характерна

LRE, как и FER, индуцирует образование АФК в нитчатом аппарате
Популярная версия:
LRE, как и FER, индуцирует образование АФК в нитчатом аппарате
Популярная версия:

Взаимодействие гамет. Оплодотворение
Двойное оплодотворение у покрытосеменных растений было открыто С.Г. Навашиным
Взаимодействие гамет. Оплодотворение
Двойное оплодотворение у покрытосеменных растений было открыто С.Г. Навашиным

Гаметы и Са2+ (in vitro)
При слиянии спермия и яйцеклетки кукурузы in
Гаметы и Са2+ (in vitro)
При слиянии спермия и яйцеклетки кукурузы in
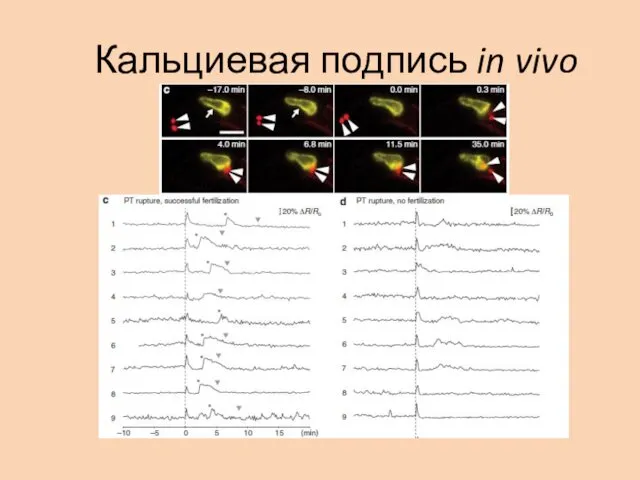
Кальциевая подпись in vivo
Кальциевая подпись in vivo

In vivo 2 пика
Более сложная картина: 2 пика!
Первый пик соответствует
In vivo 2 пика
Более сложная картина: 2 пика!
Первый пик соответствует
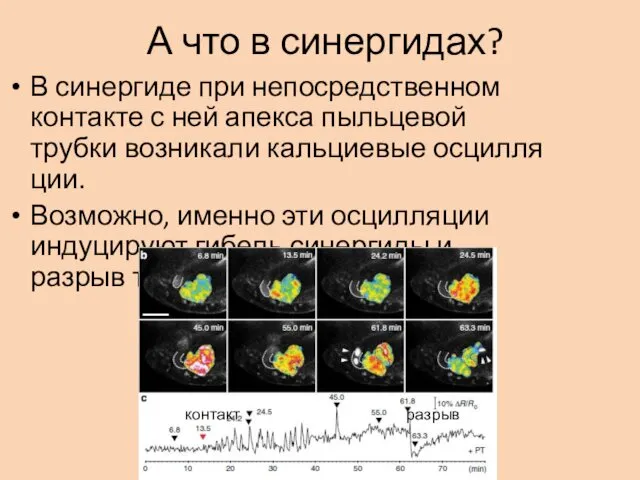
А что в синергидах?
В синергиде при непосредственном контакте с ней апекса
А что в синергидах?
В синергиде при непосредственном контакте с ней апекса
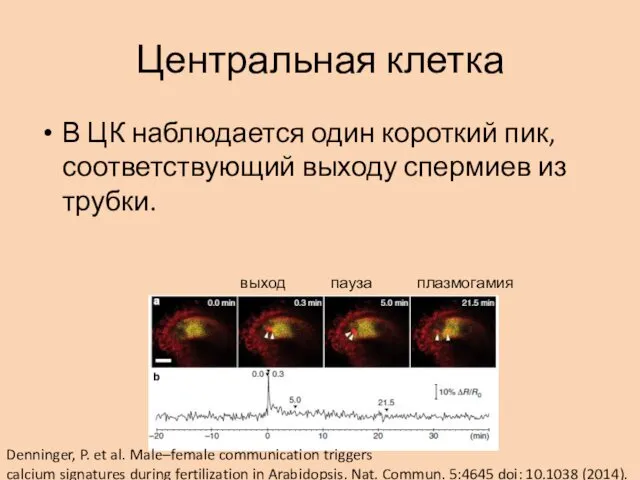
Центральная клетка
В ЦК наблюдается один короткий пик, соответствующий выходу спермиев из
Центральная клетка
В ЦК наблюдается один короткий пик, соответствующий выходу спермиев из

Диалог гамет во время «паузы»
GENERATIVE CELL SPECIFIC 1 (GCS1) у Lilium
Диалог гамет во время «паузы»
GENERATIVE CELL SPECIFIC 1 (GCS1) у Lilium

Адгезия и слияние
Оказалось, что адгезия мембран и их слияние – разные
Адгезия и слияние
Оказалось, что адгезия мембран и их слияние – разные
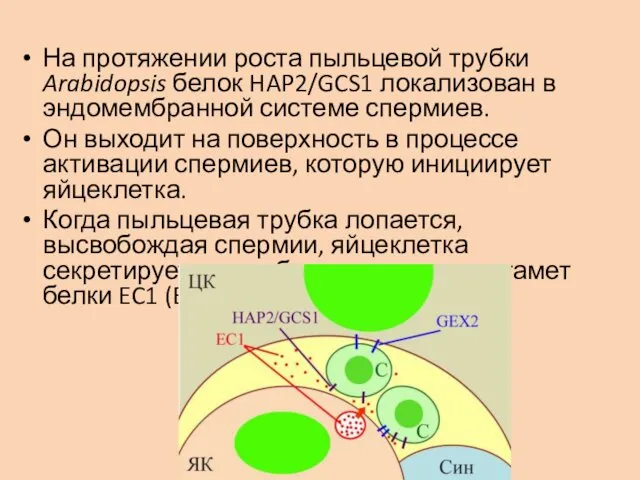
На протяжении роста пыльцевой трубки Arabidopsis белок HAP2/GCS1 локализован в эндомембранной
На протяжении роста пыльцевой трубки Arabidopsis белок HAP2/GCS1 локализован в эндомембранной

Активация гамет
При этом из эндомембранной системы на поверхность спермия выходят белки
Активация гамет
При этом из эндомембранной системы на поверхность спермия выходят белки

Яйцеклетка: скромная или активная?
В целом, слияние гамет у растений происходит почти
Яйцеклетка: скромная или активная?
В целом, слияние гамет у растений происходит почти
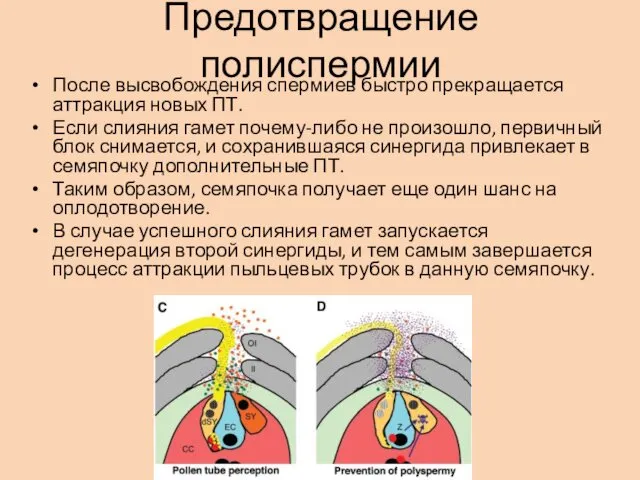
Предотвращение полиспермии
После высвобождения спермиев быстро прекращается аттракция новых ПТ.
Если слияния
Предотвращение полиспермии
После высвобождения спермиев быстро прекращается аттракция новых ПТ.
Если слияния
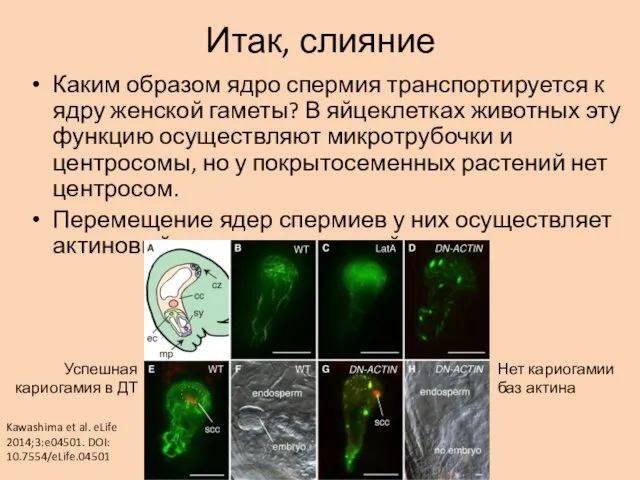
Итак, слияние
Каким образом ядро спермия транспортируется к ядру женской гаметы? В
Итак, слияние
Каким образом ядро спермия транспортируется к ядру женской гаметы? В
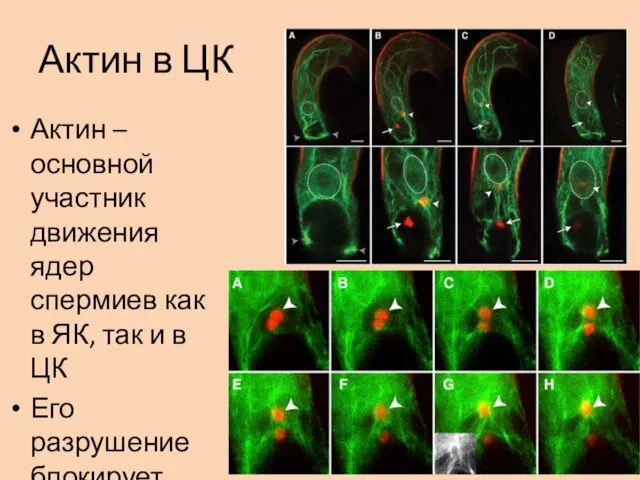
Актин в ЦК
Актин – основной участник движения ядер спермиев как в
Актин в ЦК
Актин – основной участник движения ядер спермиев как в

Детали
Ядро спермия, войдя в цитоплазму ЦК, окружалось волокнами F-актина с образованием
Детали
Ядро спермия, войдя в цитоплазму ЦК, окружалось волокнами F-актина с образованием
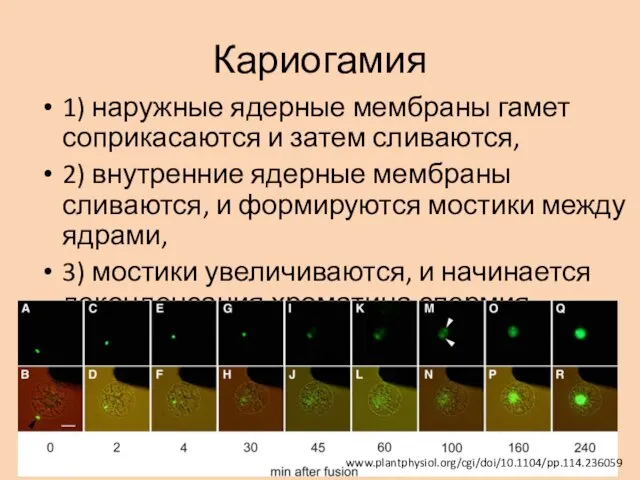
Кариогамия
1) наружные ядерные мембраны гамет соприкасаются и затем сливаются,
2) внутренние
Кариогамия
1) наружные ядерные мембраны гамет соприкасаются и затем сливаются,
2) внутренние

В системе in vitro у риса путь ядра спермия до ядра
В системе in vitro у риса путь ядра спермия до ядра

САМОНЕСОВМЕСТИМОСТЬ
Когда «родной» вовсе значит «желанный»
САМОНЕСОВМЕСТИМОСТЬ
Когда «родной» вовсе значит «желанный»

Два типа СН
Гаметофитная СН: фенотип пыльцы определяется гаплоидным геномом (гаметофитным)
Спорофитная СН:
Два типа СН
Гаметофитная СН: фенотип пыльцы определяется гаплоидным геномом (гаметофитным)
Спорофитная СН:

Гаметофитная СН. S-RNAse
Рост пыльцы подавляется только тогда, когда она вошла в
Гаметофитная СН. S-RNAse
Рост пыльцы подавляется только тогда, когда она вошла в
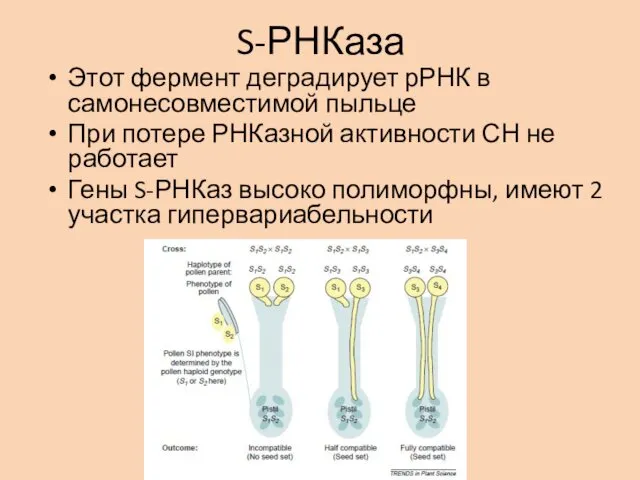
S-РНКаза
Этот фермент деградирует рРНК в самонесовместимой пыльце
При потере РНКазной активности СН
S-РНКаза
Этот фермент деградирует рРНК в самонесовместимой пыльце
При потере РНКазной активности СН
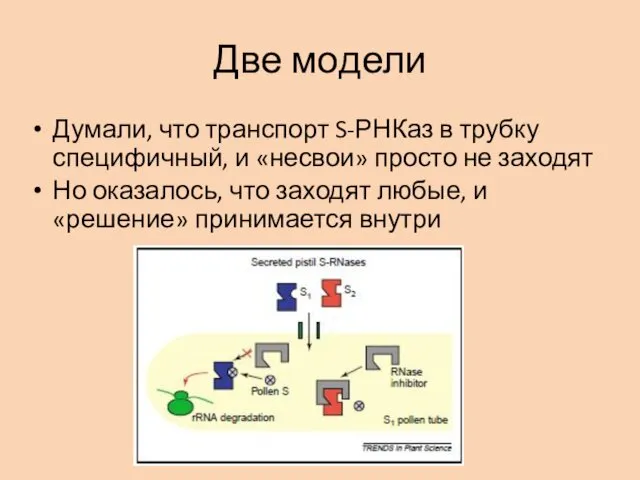
Две модели
Думали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто
Две модели
Думали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто
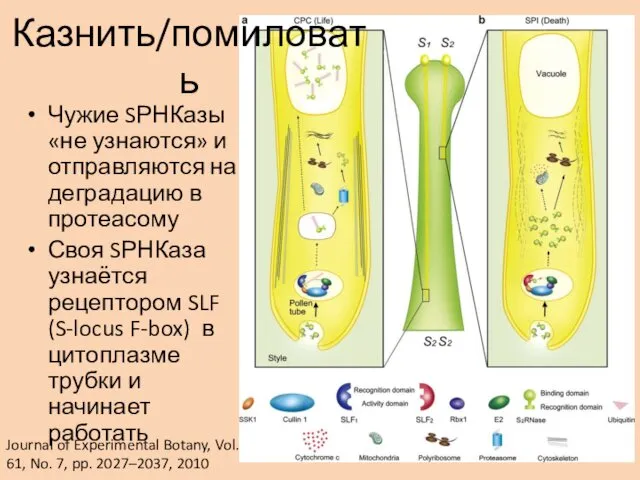
Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому
Своя SРНКаза
Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому
Своя SРНКаза
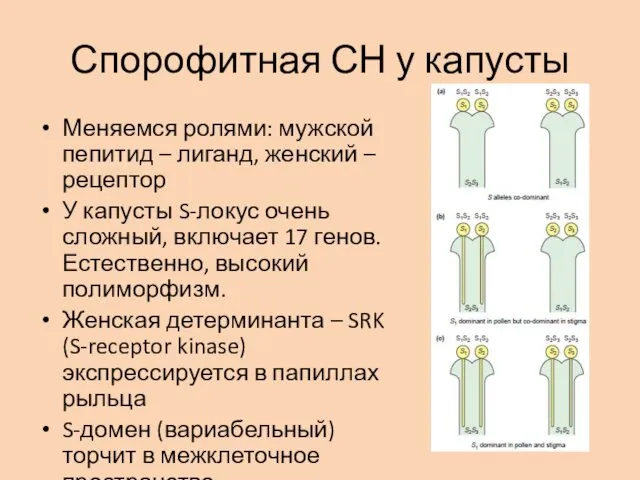
Спорофитная СН у капусты
Меняемся ролями: мужской пепитид – лиганд, женский –
Спорофитная СН у капусты
Меняемся ролями: мужской пепитид – лиганд, женский –

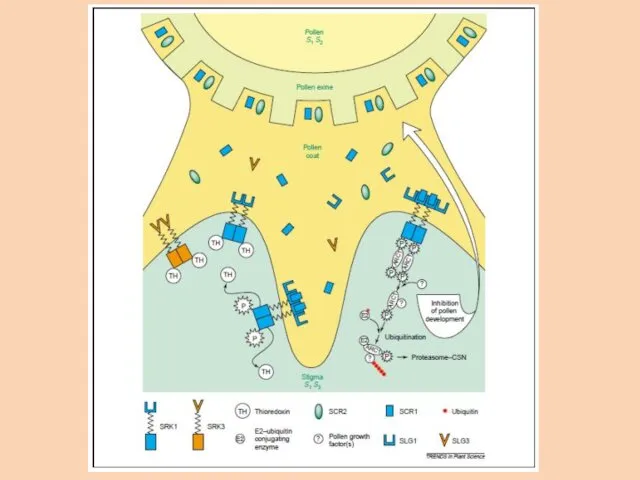
Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein)
Полиморфизм мужской
Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein)
Полиморфизм мужской
 Гормоны. Эндокринная система
Гормоны. Эндокринная система Професія дощового черв'яка
Професія дощового черв'яка Значение птиц в жизни человека
Значение птиц в жизни человека Презентация на тему "Грибы-растения или животные" - скачать презентации по Биологии
Презентация на тему "Грибы-растения или животные" - скачать презентации по Биологии Презентация урока «Работа сердца»
Презентация урока «Работа сердца» Кошки
Кошки Презентация на тему "Общая Характеристика водорослей" - скачать презентации по Биологии
Презентация на тему "Общая Характеристика водорослей" - скачать презентации по Биологии Современные методы ДНК-диагностики наследственных болезней
Современные методы ДНК-диагностики наследственных болезней Учебная презентация по теме: Растительный и животный мир Омской области
Учебная презентация по теме: Растительный и животный мир Омской области Дидактическая игра для детей Природный мир Сибири
Дидактическая игра для детей Природный мир Сибири Презентация на тему Пищеварение
Презентация на тему Пищеварение  Презентация на тему "Многообразие живых организмов" - скачать презентации по Биологии
Презентация на тему "Многообразие живых организмов" - скачать презентации по Биологии Клеточный цикл
Клеточный цикл Группа Пчёлки
Группа Пчёлки Мхи. Сфагнум
Мхи. Сфагнум Презентация на тему Органы дыхания человека
Презентация на тему Органы дыхания человека Памятка по выполнению контрольной работы по дисциплине Биология
Памятка по выполнению контрольной работы по дисциплине Биология Борьба за существование
Борьба за существование Дерево Граб
Дерево Граб Пути заражения паразитарными заболеваниями
Пути заражения паразитарными заболеваниями Климент Аркадьевич Тимирязев 1843–1920 гг. Фотосинтез
Климент Аркадьевич Тимирязев 1843–1920 гг. Фотосинтез Green House
Green House Презентация на тему экологических факторах. Основные среды жизни.
Презентация на тему экологических факторах. Основные среды жизни.  Урок- игра по теме «Описание внешности человека» 7 класс Доронина В.В., учитель русского языка и литературы ГБОУ СОШ с.Кошки С
Урок- игра по теме «Описание внешности человека» 7 класс Доронина В.В., учитель русского языка и литературы ГБОУ СОШ с.Кошки С Физиология пищеварения
Физиология пищеварения Окружающий мир Воспитываем положительные личностные качества. Узнаем как из зерна получилась булка.
Окружающий мир Воспитываем положительные личностные качества. Узнаем как из зерна получилась булка. About plants
About plants Разнообразие растений.
Разнообразие растений.