- Метаболизм липидов. Характеристика липидов. Значение. Представители. Эйкозаноиды. (Лекция 1-2)

Содержание
- 2. Метаболизм липидов. №1 1. Характеристика липидов. Значение. Представители. Эйкозаноиды. 2. Пищевые липиды их превращения в ЖКТ.
- 3. Общие свойства липидов Липиды - низкомолекулярные органические соединения биологического происхождения . К липидам относятся представители разных
- 4. Биологические функции липидов. 1. Структурная. В комплексе с белками – липиды структурный компонент всех биологических мембран
- 5. Классы, строение, функции липидов
- 6. Основные классы липидов Жирные кислоты; триацилглицериды; глицерофосфолипиды; стероиды - холестерин; сфингофосфолипиды, сфингомиелины; цереброзиды Суточная потребность пищевых
- 7. Основные высшие жирные кислоты организма ВЖК - структурный компонент практически всех липидов (кроме свободного холестерола) СН3
- 8. Эйкозаноиды Эйкозаноиды – БАВ, синтезируемые большинством клеток из полиеновых жирных кислот, содержащих 20 углерод- ных атомов
- 9. Образование эйкозаноидов. Условие образования: активация фосфолипазы А2 отщепление арахидоновой кислоты (при воспалениях!!); 2 пути синтеза эйкозаноидов:
- 10. Этапы превращений липидов в желудочно-кишечном тракте 1. Переваривание ( 12-перстная кишка.). - эмульгирование; - ферментативный гидролиз.
- 11. Эмульгирование. Основные эмульгаторы- желчные кислоты. Эмульгирование пищевых липидов – этап, предшествующий ферментативному гидролизу панкреатическими липазами, необходим
- 12. Эмульгирование Гидрофильная часть молекулы эмульгатора Гидрофобная часть молекулы эмульгатора, погружается в жировую каплю Эмульгаторы-поверхностно-активные в-ва Снижают
- 13. Переваривание триацилглицеролов в кишечнике сн2-о-со-R1 сн2-он сн2-он сн2-он I I I I cн-о-со-R2 cн-о-со-R2 сн-о-со-R2 сн-оH
- 14. Переваривание фосфоглицеролипидов в кишечнике сн2-о-соR1 I I сн-о-соR2 сн-он I I сн2-о-р-о-сн2-сн2-N(cн3)3 сн2-о-р-о-сн2-сн2-N(сн3)3 фосфатидилхолин Фосфолипаза А2
- 15. На заметку стоматологу и педиатору На слизистой оболочке корня языка – в железах фон Эбнера синтезируется
- 16. На заметку врачу Слюнной секрет ядовитых змей и скорпионов содержит очень активную фосфолипазу А2. При укусе
- 17. Всасывание продуктов переваривания в слизистой тонкого кишечника Растворимые продукты гидролиза в к-ки кишечника (энтероциты) всасываются самостоятельно.
- 18. Ресинтез жиров в слизистой кишечника ( синтез из продуктов переваривания специфичных липидов в слизистой кишечника 2
- 19. Упаковка ресинтезированных жиров в транспортные формы Для транспортировки кровотоком гидрофобных липидов необходимы специфические транспортные формы. В
- 20. Транспортные формы липидов. Типы Хиломикроны – эпителий тонкого кишечника Липопротеины очень низкой плотности – клетки печени
- 21. Общая схема строения липопротеина ТАГ Эфиры холестерола Гидрофобные липиды холестерол ФЛ Интегральные апопротеины: Апо В-100; АпоВ-48
- 22. Транспортные формы липидов в крови Значение апопротеинов. Формируют структуру липопротеинов – апоВ-48; апо В-100 Взаимодействуют с
- 23. Транспорт хиломикронов в крови Вопрос: Почему в течении 4-5 часов после приема жирной пищи плазма мутная
- 24. Транспорт экзогенных липидов от кишечника к тканям. Хиломикроны В клетках слизистой кишечника образуются «незрелые» хиломикроны Лимфа,кровь
- 25. Метаболизм липидов №2 Метаболизм триацилглицеролов. Депонирование и мобилизация нейтрального жира. Гормональная регуляция липолиза Бета - окисление
- 26. Триацилглицерол ( ТАГ или нейтральный жир) сн2-о-со-R1 I cн-о-со-R2 I cн2-о-со-R3 I
- 27. Локализация синтеза ТАГ В печени и жировой ткани (преимущественно) В жировой ткани – жиры синтезируются и
- 28. Фосфатидная кислота СН2 – О- СО- R1 I СН – О-СО- R2 I СН2 – О
- 29. Пути образования глицерол -3- фосфата в печени и жировой ткани СН2 - ОН I СН -
- 30. Пути образования глицерол -3-фосфата в печени и жировой ткани СН2 - ОН I СН - ОН
- 31. Источники жирных кислот для синтеза ТАГ Жировая ткань. а) Преимущественно ЖК, освободившиеся при гидролизе жиров ХМ(
- 32. Синтез ТАГ 1. Активация жирной кислоты: R 1 СООН + АТФ + НSКоА → R 1СО-
- 33. Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как источник энергии в период голодания
- 34. Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция Тканевой липолиз ( мобилизация тканевого жира в клетках тканей
- 35. Регуляция тканевого липолиза
- 36. Источники и значение жирных кислот Источники: - Продукты гидролиза (переваривания) экзогенных жиров; Продукты тканевого липолиза; Синтез
- 37. Бета-окисление жирных кислот Жирные кислоты активируются в цитозоле АТФ и НSКоА Окисление - в матриксе митохондрий
- 38. Бета-окисление жирных кислот
- 39. Окисление глицерина СН2 - ОН СН2 – ОН С ОН I 1 I 2 I СН
- 40. Синтез ВЖК В абсорбтивный период (избыток углеводов). В цитозоле. Активируется инсулином Субстрат: Ацетил-КоА- метаболит окисления углеводов
- 41. Особенности синтеза ВЖК Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК. Катализируются реакции полифункциональным ферментом
- 42. Синтез кетоновых тел Кетоновые тела - продукты конденсации двух молекул ацетил-КоА (СН3 - СО- КоА). Кетоновые
- 43. Синтез кетоновых тел
- 45. Скачать презентацию

Метаболизм липидов. №1
1. Характеристика липидов. Значение.
Представители. Эйкозаноиды.
2. Пищевые
Метаболизм липидов. №1
1. Характеристика липидов. Значение.
Представители. Эйкозаноиды.
2. Пищевые

Общие свойства липидов
Липиды - низкомолекулярные органические соединения биологического происхождения .
К липидам
Общие свойства липидов
Липиды - низкомолекулярные органические соединения биологического происхождения .
К липидам

Биологические функции липидов.
1. Структурная. В комплексе с белками – липиды структурный
Биологические функции липидов.
1. Структурная. В комплексе с белками – липиды структурный
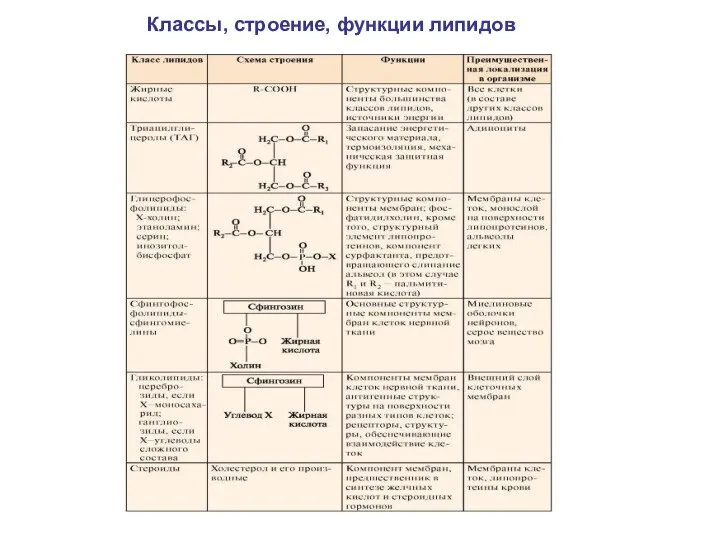
Классы, строение, функции липидов
Классы, строение, функции липидов

Основные классы липидов
Жирные кислоты;
триацилглицериды; глицерофосфолипиды;
стероиды - холестерин;
сфингофосфолипиды, сфингомиелины;
Основные классы липидов
Жирные кислоты;
триацилглицериды; глицерофосфолипиды;
стероиды - холестерин;
сфингофосфолипиды, сфингомиелины;

Основные высшие жирные кислоты организма
ВЖК - структурный компонент практически всех липидов
Основные высшие жирные кислоты организма
ВЖК - структурный компонент практически всех липидов

Эйкозаноиды
Эйкозаноиды – БАВ, синтезируемые большинством клеток из полиеновых жирных кислот, содержащих
Эйкозаноиды
Эйкозаноиды – БАВ, синтезируемые большинством клеток из полиеновых жирных кислот, содержащих

Образование эйкозаноидов.
Условие образования: активация фосфолипазы А2 отщепление арахидоновой кислоты (при воспалениях!!);
2
Образование эйкозаноидов.
Условие образования: активация фосфолипазы А2 отщепление арахидоновой кислоты (при воспалениях!!);
2

Этапы превращений липидов в желудочно-кишечном тракте
1. Переваривание ( 12-перстная кишка.).
Этапы превращений липидов в желудочно-кишечном тракте
1. Переваривание ( 12-перстная кишка.).

Эмульгирование. Основные эмульгаторы- желчные кислоты.
Эмульгирование пищевых липидов – этап, предшествующий ферментативному
Эмульгирование. Основные эмульгаторы- желчные кислоты.
Эмульгирование пищевых липидов – этап, предшествующий ферментативному
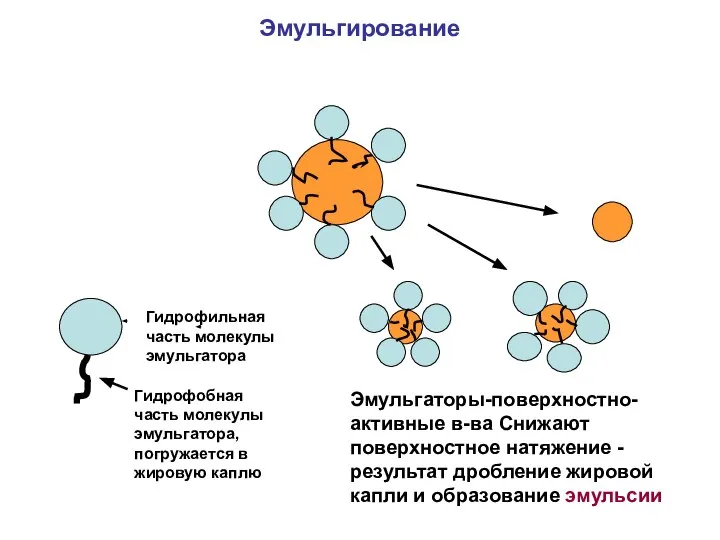
Эмульгирование
Гидрофильная часть молекулы эмульгатора
Гидрофобная часть молекулы эмульгатора, погружается в жировую каплю
Эмульгаторы-поверхностно-активные
Эмульгирование
Гидрофильная часть молекулы эмульгатора
Гидрофобная часть молекулы эмульгатора, погружается в жировую каплю
Эмульгаторы-поверхностно-активные
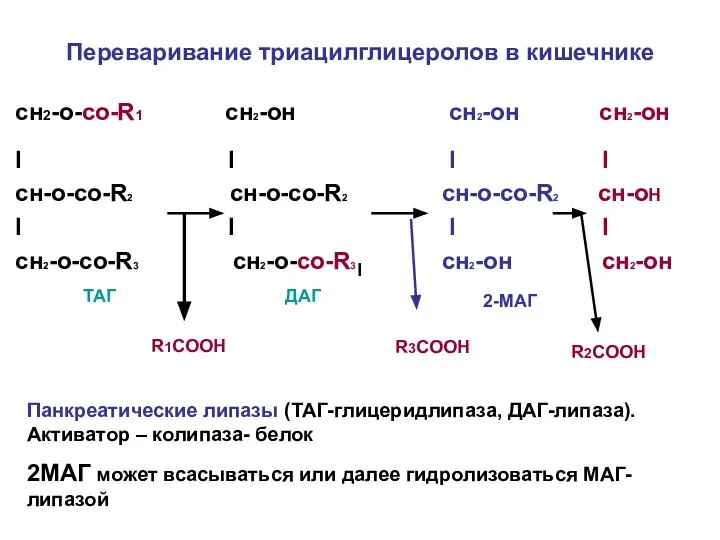
Переваривание триацилглицеролов в кишечнике
сн2-о-со-R1 сн2-он сн2-он сн2-он
I I I I
cн-о-со-R2 cн-о-со-R2
Переваривание триацилглицеролов в кишечнике
сн2-о-со-R1 сн2-он сн2-он сн2-он
I I I I
cн-о-со-R2 cн-о-со-R2
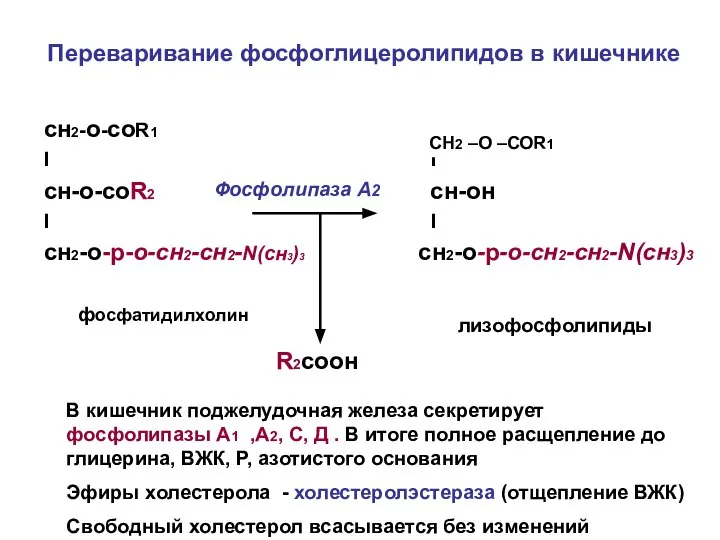
Переваривание фосфоглицеролипидов в кишечнике
сн2-о-соR1
I I
сн-о-соR2 сн-он
I I
сн2-о-р-о-сн2-сн2-N(cн3)3 сн2-о-р-о-сн2-сн2-N(сн3)3
фосфатидилхолин
Фосфолипаза А2
-о-соR1
лизофосфолипиды
CН2 –О
Переваривание фосфоглицеролипидов в кишечнике
сн2-о-соR1
I I
сн-о-соR2 сн-он
I I
сн2-о-р-о-сн2-сн2-N(cн3)3 сн2-о-р-о-сн2-сн2-N(сн3)3
фосфатидилхолин
Фосфолипаза А2
-о-соR1
лизофосфолипиды
CН2 –О

На заметку стоматологу и педиатору
На слизистой оболочке корня языка – в
На заметку стоматологу и педиатору
На слизистой оболочке корня языка – в

На заметку врачу
Слюнной секрет ядовитых змей и скорпионов содержит очень
На заметку врачу
Слюнной секрет ядовитых змей и скорпионов содержит очень

Всасывание продуктов переваривания в слизистой тонкого кишечника
Растворимые продукты гидролиза в к-ки
Всасывание продуктов переваривания в слизистой тонкого кишечника
Растворимые продукты гидролиза в к-ки

Ресинтез жиров в слизистой кишечника
( синтез из продуктов переваривания
Ресинтез жиров в слизистой кишечника
( синтез из продуктов переваривания

Упаковка ресинтезированных жиров в транспортные формы
Для транспортировки кровотоком гидрофобных липидов необходимы
Упаковка ресинтезированных жиров в транспортные формы
Для транспортировки кровотоком гидрофобных липидов необходимы

Транспортные формы липидов. Типы
Хиломикроны – эпителий тонкого кишечника
Липопротеины очень низкой плотности
Транспортные формы липидов. Типы
Хиломикроны – эпителий тонкого кишечника
Липопротеины очень низкой плотности
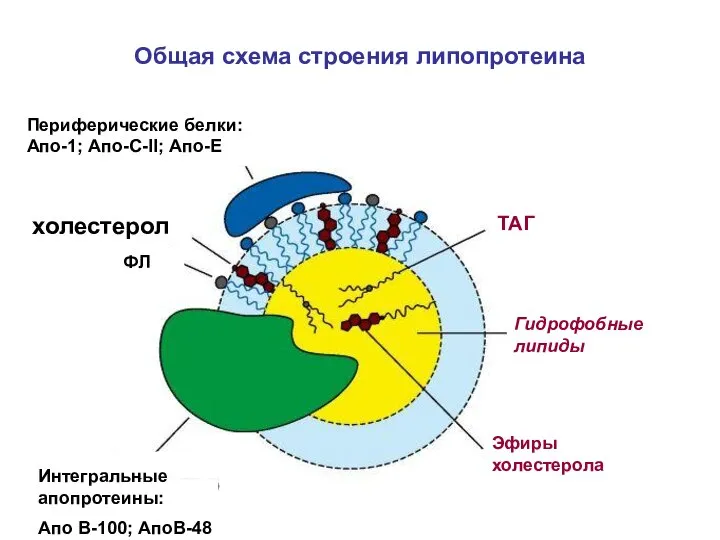
Общая схема строения липопротеина
ТАГ
Эфиры холестерола
Гидрофобные липиды
холестерол
ФЛ
Интегральные апопротеины:
Апо В-100; АпоВ-48
Периферические белки:
Общая схема строения липопротеина
ТАГ
Эфиры холестерола
Гидрофобные липиды
холестерол
ФЛ
Интегральные апопротеины:
Апо В-100; АпоВ-48
Периферические белки:

Транспортные формы липидов в крови
Значение апопротеинов.
Формируют структуру липопротеинов – апоВ-48; апо
Транспортные формы липидов в крови
Значение апопротеинов.
Формируют структуру липопротеинов – апоВ-48; апо

Транспорт хиломикронов в крови
Вопрос:
Почему в течении 4-5 часов после приема жирной
Транспорт хиломикронов в крови
Вопрос:
Почему в течении 4-5 часов после приема жирной

Транспорт экзогенных липидов от кишечника к тканям. Хиломикроны
В клетках слизистой кишечника
Транспорт экзогенных липидов от кишечника к тканям. Хиломикроны
В клетках слизистой кишечника

Метаболизм липидов №2
Метаболизм триацилглицеролов.
Депонирование и мобилизация нейтрального жира.
Гормональная регуляция
Метаболизм липидов №2 Метаболизм триацилглицеролов. Депонирование и мобилизация нейтрального жира. Гормональная регуляция
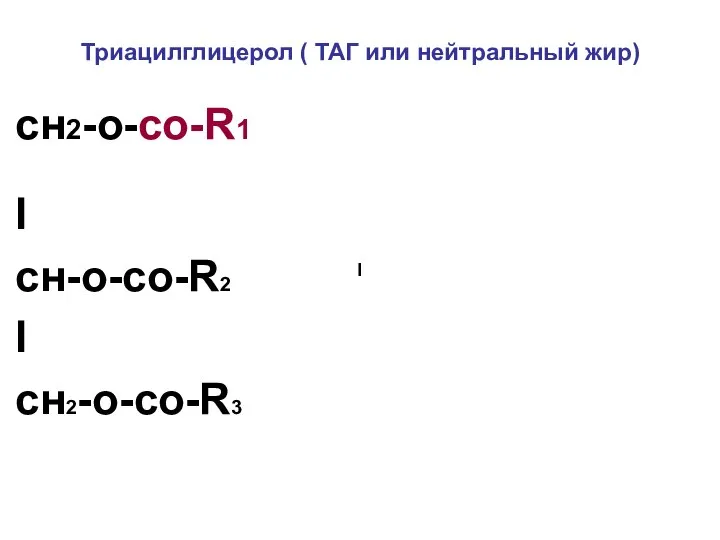
Триацилглицерол ( ТАГ или нейтральный жир)
сн2-о-со-R1
I
cн-о-со-R2
I
cн2-о-со-R3
I
Триацилглицерол ( ТАГ или нейтральный жир)
сн2-о-со-R1
I
cн-о-со-R2
I
cн2-о-со-R3
I

Локализация синтеза ТАГ
В печени и жировой ткани (преимущественно)
В жировой ткани –
Локализация синтеза ТАГ
В печени и жировой ткани (преимущественно)
В жировой ткани –
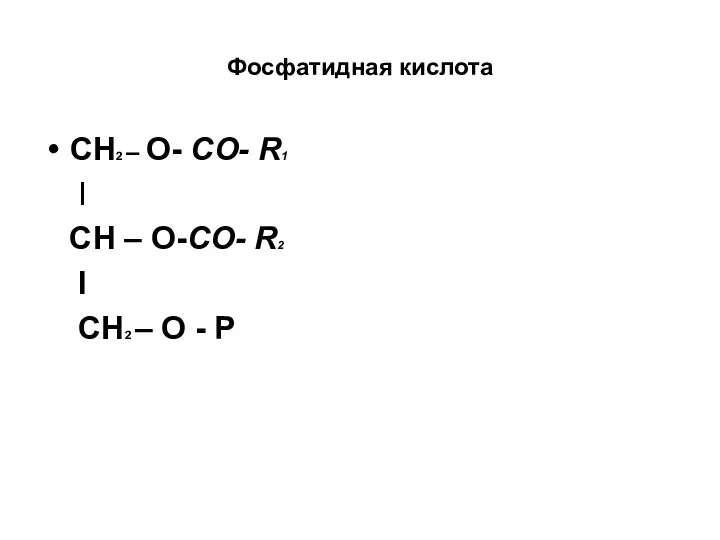
Фосфатидная кислота
СН2 – О- СО- R1
I
СН – О-СО- R2
Фосфатидная кислота
СН2 – О- СО- R1
I
СН – О-СО- R2

Пути образования глицерол -3- фосфата в печени и жировой ткани
СН2
Пути образования глицерол -3- фосфата в печени и жировой ткани
СН2

Пути образования глицерол -3-фосфата в печени и жировой ткани
СН2 -
Пути образования глицерол -3-фосфата в печени и жировой ткани
СН2 -

Источники жирных кислот для синтеза ТАГ
Жировая ткань.
а) Преимущественно ЖК, освободившиеся
Источники жирных кислот для синтеза ТАГ
Жировая ткань.
а) Преимущественно ЖК, освободившиеся

Синтез ТАГ
1. Активация жирной кислоты:
R 1 СООН + АТФ
Синтез ТАГ
1. Активация жирной кислоты:
R 1 СООН + АТФ

Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как
Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как

Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция
Тканевой липолиз ( мобилизация тканевого
Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция
Тканевой липолиз ( мобилизация тканевого
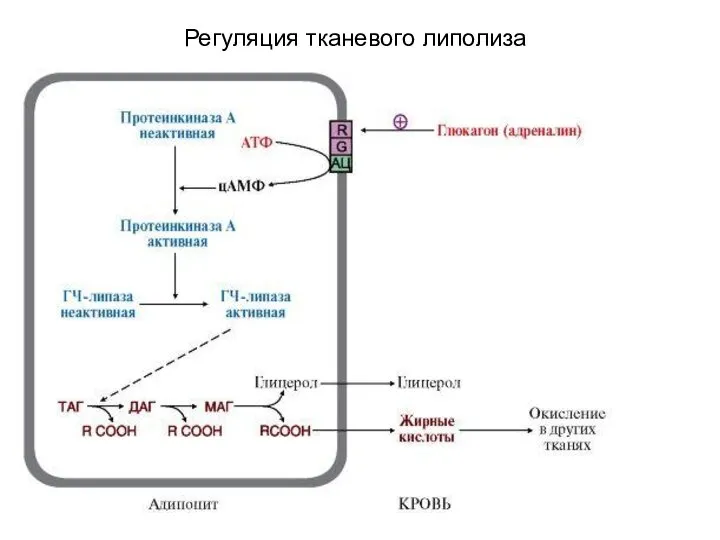
Регуляция тканевого липолиза
Регуляция тканевого липолиза

Источники и значение жирных кислот
Источники:
- Продукты гидролиза (переваривания) экзогенных жиров;
Продукты
Источники и значение жирных кислот
Источники:
- Продукты гидролиза (переваривания) экзогенных жиров;
Продукты

Бета-окисление жирных кислот
Жирные кислоты активируются в цитозоле АТФ и НSКоА
Окисление -
Бета-окисление жирных кислот
Жирные кислоты активируются в цитозоле АТФ и НSКоА
Окисление -
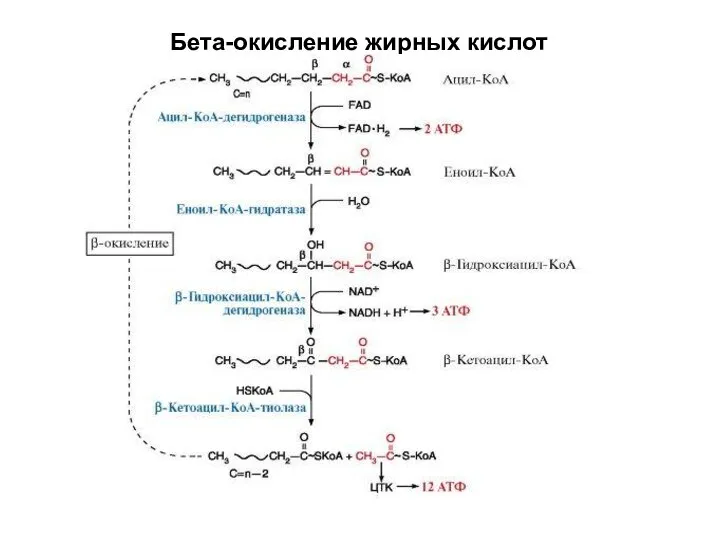
Бета-окисление жирных кислот
Бета-окисление жирных кислот
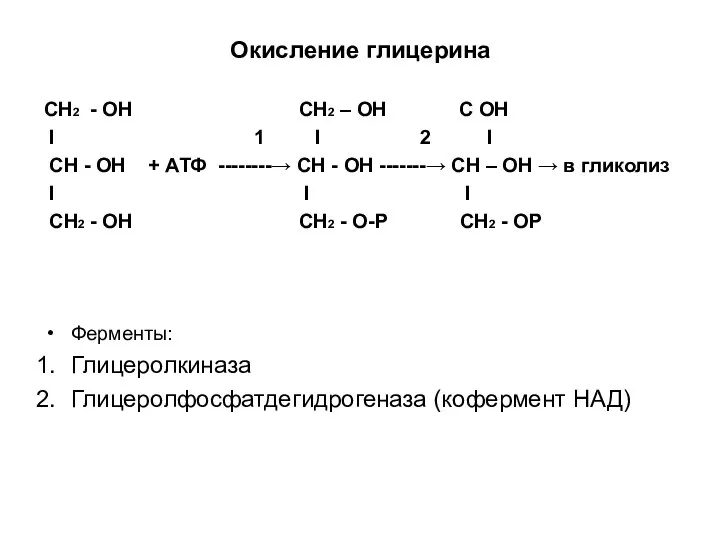
Окисление глицерина
СН2 - ОН СН2 – ОН С ОН
I 1
Окисление глицерина
СН2 - ОН СН2 – ОН С ОН
I 1

Синтез ВЖК
В абсорбтивный период (избыток углеводов). В цитозоле. Активируется инсулином
Субстрат: Ацетил-КоА-
Синтез ВЖК
В абсорбтивный период (избыток углеводов). В цитозоле. Активируется инсулином
Субстрат: Ацетил-КоА-

Особенности синтеза ВЖК
Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК.
Особенности синтеза ВЖК
Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК.

Синтез кетоновых тел
Кетоновые тела - продукты конденсации двух молекул ацетил-КоА (СН3
Синтез кетоновых тел
Кетоновые тела - продукты конденсации двух молекул ацетил-КоА (СН3
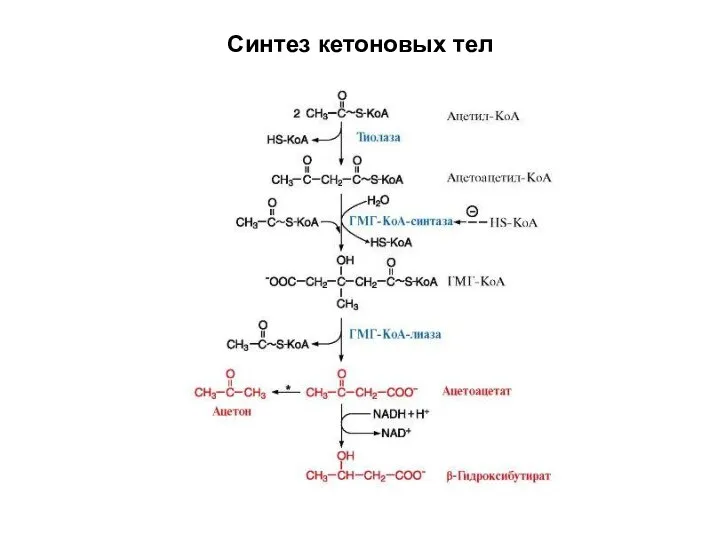
Синтез кетоновых тел
Синтез кетоновых тел
 Влияние минерального питания на рост и развитие растений Авторы: Дюрягина Е.Е., Плетнёва П.В.
Влияние минерального питания на рост и развитие растений Авторы: Дюрягина Е.Е., Плетнёва П.В. Насекомые рекордсмены
Насекомые рекордсмены Презентация на тему Экосистема озера
Презентация на тему Экосистема озера  Презентация на тему "Влияние внешних условий на рост и развитие плесневых грибов" - скачать презентации по Биологии
Презентация на тему "Влияние внешних условий на рост и развитие плесневых грибов" - скачать презентации по Биологии Пингвины
Пингвины Регуляция пищеварения. Гигиена органов пищеварения
Регуляция пищеварения. Гигиена органов пищеварения Гербарий моего двора
Гербарий моего двора Семейство медвежьи
Семейство медвежьи Земноводные - презентации по Биологии
Земноводные - презентации по Биологии Блохи (Siphonapterа)
Блохи (Siphonapterа) Исследование различных по качественному составу соков с целью более эффективного использования их для укрепления здоровья
Исследование различных по качественному составу соков с целью более эффективного использования их для укрепления здоровья Презентация на тему Живой организм и его свойства Какими свойствами обладают живые организмы?
Презентация на тему Живой организм и его свойства Какими свойствами обладают живые организмы? Лишайники
Лишайники Способи розмноження рослин Презентацію підготувала учениця 11 класу Циганок Наталія
Способи розмноження рослин Презентацію підготувала учениця 11 класу Циганок Наталія  Возникновение жизни на земле
Возникновение жизни на земле Подтип Хелицеровые (Chelicerata). Класс Паукообразные (Arachnida). Особенности организации
Подтип Хелицеровые (Chelicerata). Класс Паукообразные (Arachnida). Особенности организации Осеннее явление в лесной экосистеме Кодры
Осеннее явление в лесной экосистеме Кодры Строение ногтя
Строение ногтя Насекомые
Насекомые Фотосинтез. Понятие фотосинтеза
Фотосинтез. Понятие фотосинтеза Презентация на тему "Головной мозг, его строение и функции" - скачать презентации по Биологии
Презентация на тему "Головной мозг, его строение и функции" - скачать презентации по Биологии Вплив алкоголю на організм людини Робота учениці 6-го класу Михайлівської ЗОШ І-ІІст Дзензюр Ангеліни
Вплив алкоголю на організм людини Робота учениці 6-го класу Михайлівської ЗОШ І-ІІст Дзензюр Ангеліни  Половой отбор? Павлины против!
Половой отбор? Павлины против! ДИНОЗАВР
ДИНОЗАВР Русская выхухоль имеет право на жизнь
Русская выхухоль имеет право на жизнь Хвойные
Хвойные Царство Растения. Кружок Горизонты биологии: система живых организмов
Царство Растения. Кружок Горизонты биологии: система живых организмов Межклеточные контакты
Межклеточные контакты