- Митохондрии. Хлоропласты

Содержание
- 2. Митохондрии – окраска по Альтману
- 3. Колокализация митохондрий и микротрубочек Мтх – окраска родамином, МТ- окраска на альфа-тубулин
- 4. Митохондрии в мышце - хондриом
- 5. Митохондрия – срез (ЭМ)
- 6. Митохондрии в живой клетке – окраска родамином 123
- 7. Компоненты митохондрий Наружная мембрана: гладкая; содержит мало белков (отношение липид/белок ~1:1); проницаема для веществ с молекулярным
- 8. Структура митохондрий
- 9. Транспорт белков в митохондрии Сигнальная последовательность митохондриальных белков, как правило, расположена на N-конце, и она отщепляется
- 10. Динамика митохондрий Митохондрии в клетке имеют непостоянную форму и функционально связаны в единую сеть (быстро обмениваются
- 11. Фрагментация митохондрий Физиологическая роль фрагментации митохондрий – удаление дефектных компартментов органелл.
- 12. Регуляция слияния митохондрий
- 13. Основные функции митохондрий Окислительное фосфорилирование: генерация разности потенциалов в результате окисления органических субстратов; синтез АТФ в
- 14. Цепь переноса электронов и цикл трикарбоновых кислот
- 15. Перенос электронов в мембране митохондрии
- 16. Цепь переноса электронов
- 17. Работа внутренней митохондриальной мембраны
- 18. АТФ-аза митохондрий и прокариот АТФ-аза – миниатюрный электромотор, вмонтированный в мембрану перпендикулярно ее плоскости. Ротор –
- 19. АТФ-АДФ транслоказа Составляет около 10% от общего белка внутренней мембраны. Весьма консервативна, в особенности, канал для
- 20. Хемиосмотическая теория Цепь переноса электронов переносит протоны из матрикса митохондрии в цитозоль в процессе снижения уровня
- 21. Генерация энергии митохондриями Универсальная форма энергии – электрохимический потенциал на мембране (ΔH+ у эукариот, ΔН+ или
- 22. Регуляция окислительного фосфорилирования через систему обратных связей Дыхательный контроль – в норме потребление кислорода ограничивается концентрацией
- 23. Ингибиторы митохондрий Разобщители – FCCP, 2,4-ДНФ, жирные кислоты. Повышают проницаемость мембраны для протонов. Сильнейшие яды. Увеличивают
- 24. Митохондрия как терморегулятор Жирные кислоты – природные разобщители, которые регулируют соотношение окисления и синтеза АТФ. Митохондрии
- 25. Энергетические затраты животной клетки Работа K/Na–АТФазы – 15-60% энергии (в среднем около 35%). Поддержание протонного градиента
- 26. Растительная клетка с хлоропластами и вакуолью
- 27. Схема организации хлоропласта
- 28. Хлоропласт, вид на срезе Характерный размер – 1х5 мкм. Нередко хлоропласты образуют сеть, имея непрерывную наружную
- 29. Хлоропласт, детали структуры
- 30. Мембраны хлоропласта Наружная мембрана: гладкая; высокопроницаемая. Внутренняя мембрана: гладкая (не образует крист); практически непроницаема для любых
- 31. Компоненты хлоропласта Строма имеет щелочной рН. Она содержит нуклеиновые кислоты (ДНК, РНК) и рибосомы хлоропласта; содержит
- 32. Основы фотосинтеза Возможные преобразования энергии кванта света в молекулярном ансамбле: 1. флуоресценция 2. Ферстеровский резонансный перенос
- 33. Световые реакции фотосинтеза Фотосистема II – комплекс из примерно 20 белков с большим количеством трансмембранных доменов.
- 34. Перенос электронов в мембране хлоропласта Перенос протонов поперек мембраны граны хлоропласта аналогичен переносу поперек внутренней мембраны
- 35. Цепь переноса электронов и фотосистемы
- 36. Световые реакции
- 37. Световые и темновые реакции фотосинтеза Световые реакции – зависят от постоянного потока фотонов и протекают на
- 38. Темновые реакции фотосинтеза
- 39. Развитие хлоропластов Хлоропласты (например, в побегах) развиваются под действием света из пропластид, лишенных гран (т.н. агранулярные
- 40. Дифференцировка хлоропластов Пропластиды – бесцветные двумембранные структуры, диаметром 0,4-1 мкм, содержащие ДНК хлоропластов. Они содержатся в
- 41. Деление хлоропластов Деление хлоропластов происходит медленно. Оно регулируется несколькими группами белков – динамин, FtsZ (аналог прокариотического
- 42. Сравнение митохондрий и хлоропластов Первичные источники энергии Цепь переноса электронов на мембране Протонная помпа – синтез
- 43. Митохондрии и хлоропласты: нуклеиновые кислоты Митохондрия: кольцевые молекулы ДНК – 16 кБ, кодируют 13 белков (часть
- 44. Митохондрии и хлоропласты: полуавтономность Митохондрия: может функционировать без собственной ДНК, однако эффективность дыхания резко снижается. У
- 46. Скачать презентацию
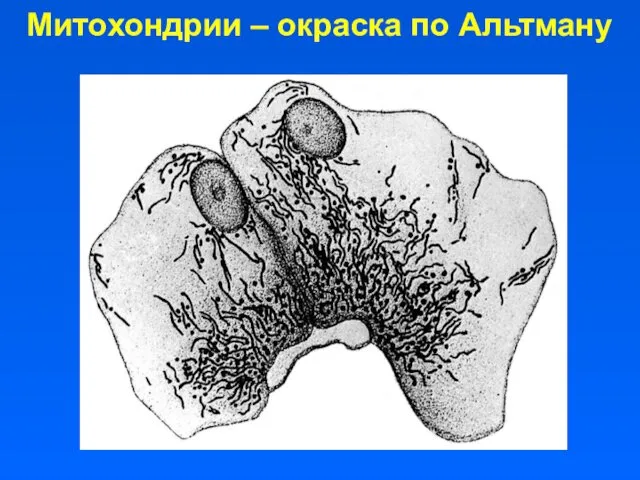
Митохондрии – окраска по Альтману
Митохондрии – окраска по Альтману
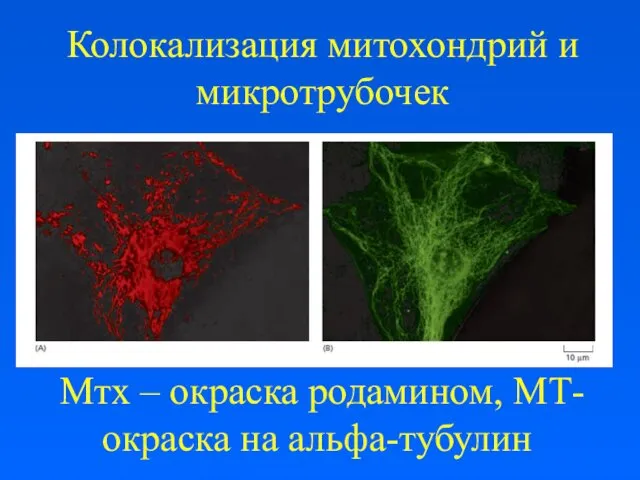
Колокализация митохондрий и микротрубочек
Мтх – окраска родамином, МТ- окраска на альфа-тубулин
Колокализация митохондрий и микротрубочек
Мтх – окраска родамином, МТ- окраска на альфа-тубулин

Митохондрии в мышце - хондриом
Митохондрии в мышце - хондриом
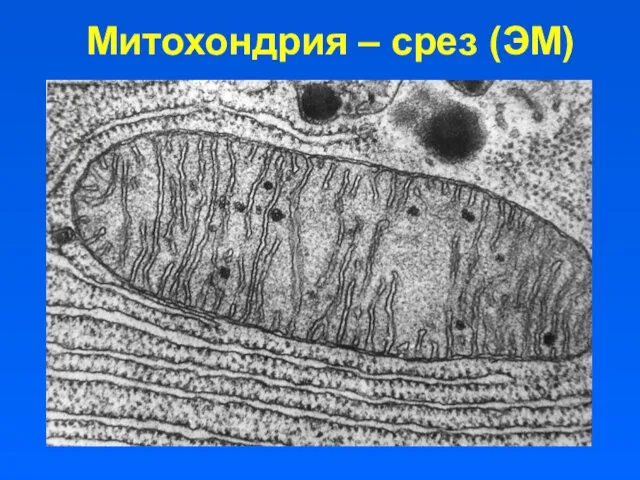
Митохондрия – срез (ЭМ)
Митохондрия – срез (ЭМ)
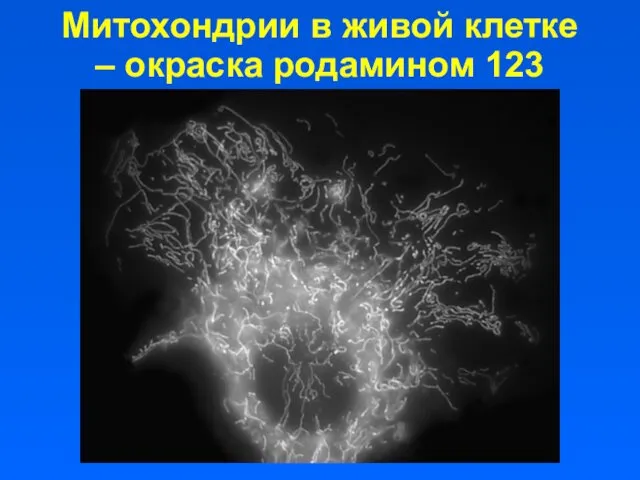
Митохондрии в живой клетке – окраска родамином 123
Митохондрии в живой клетке – окраска родамином 123

Компоненты митохондрий
Наружная мембрана: гладкая; содержит мало белков (отношение липид/белок ~1:1); проницаема
Компоненты митохондрий
Наружная мембрана: гладкая; содержит мало белков (отношение липид/белок ~1:1); проницаема
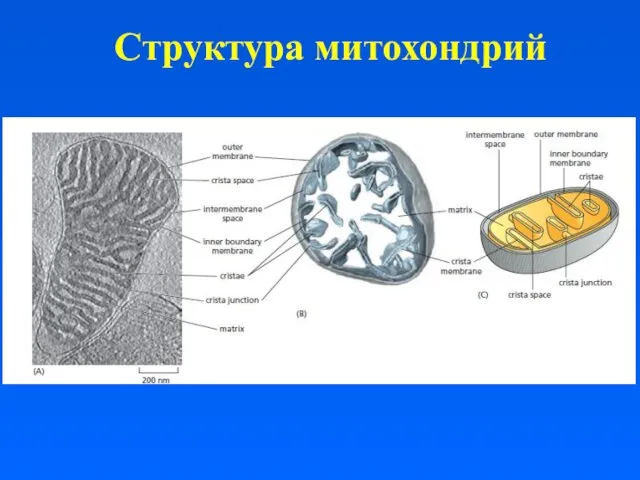
Структура митохондрий
Структура митохондрий

Транспорт белков в митохондрии
Сигнальная последовательность митохондриальных белков, как правило, расположена на
Транспорт белков в митохондрии
Сигнальная последовательность митохондриальных белков, как правило, расположена на

Динамика митохондрий
Митохондрии в клетке имеют непостоянную форму и функционально связаны в
Динамика митохондрий
Митохондрии в клетке имеют непостоянную форму и функционально связаны в
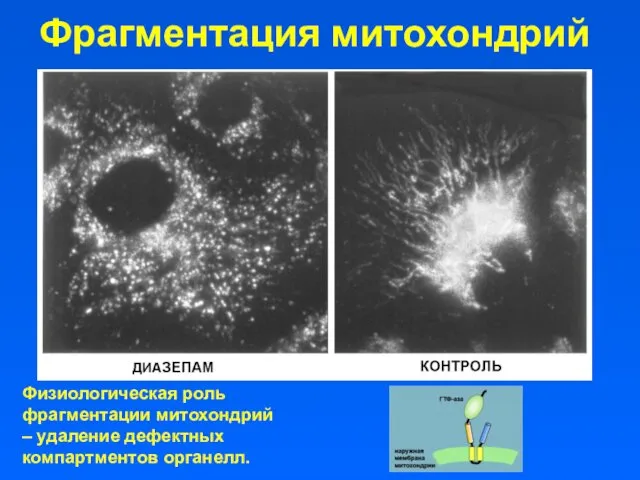
Фрагментация митохондрий
Физиологическая роль фрагментации митохондрий – удаление дефектных компартментов органелл.
Фрагментация митохондрий
Физиологическая роль фрагментации митохондрий – удаление дефектных компартментов органелл.
Регуляция слияния митохондрий
Регуляция слияния митохондрий

Основные функции митохондрий
Окислительное фосфорилирование: генерация разности потенциалов в результате окисления органических
Основные функции митохондрий
Окислительное фосфорилирование: генерация разности потенциалов в результате окисления органических
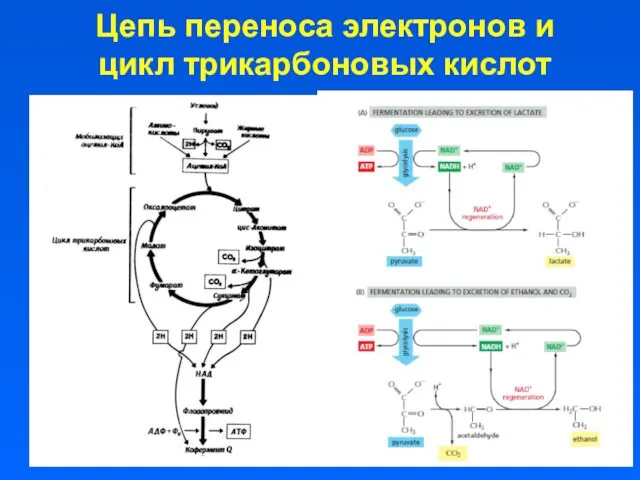
Цепь переноса электронов и цикл трикарбоновых кислот
Цепь переноса электронов и цикл трикарбоновых кислот
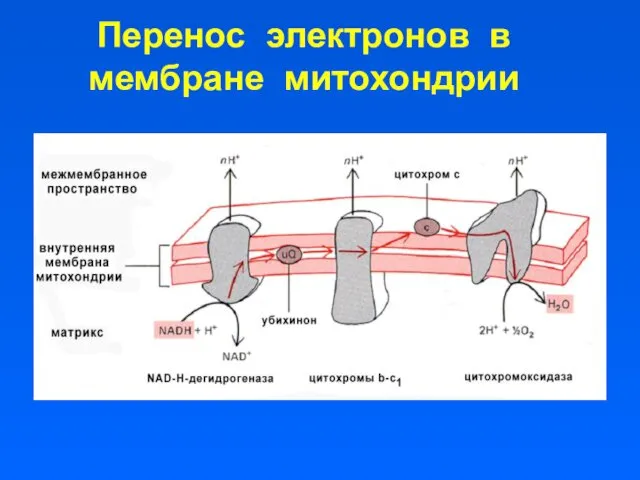
Перенос электронов в мембране митохондрии
Перенос электронов в мембране митохондрии
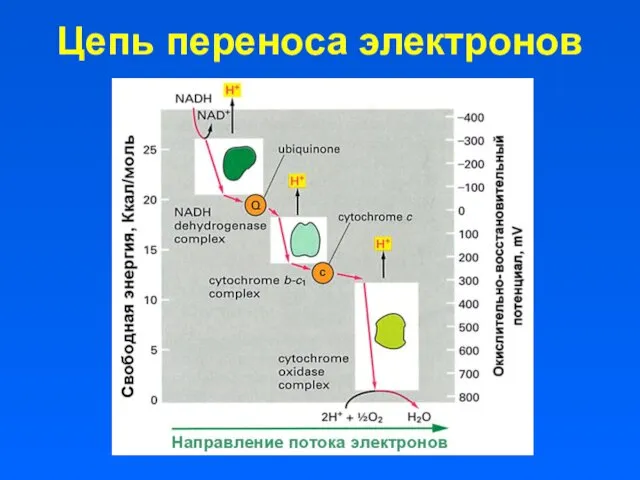
Цепь переноса электронов
Цепь переноса электронов
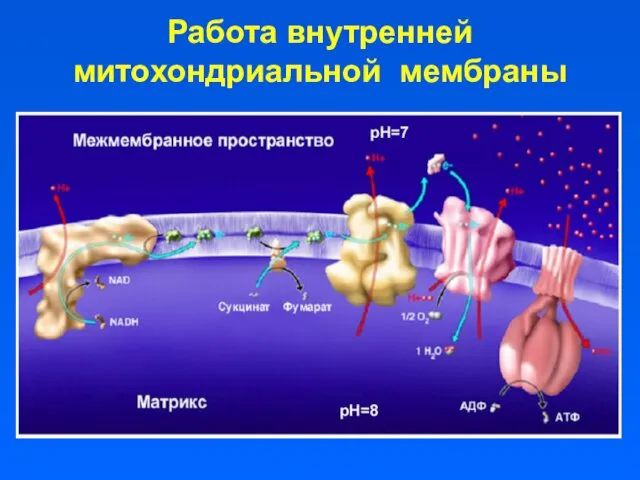
Работа внутренней митохондриальной мембраны
Работа внутренней митохондриальной мембраны
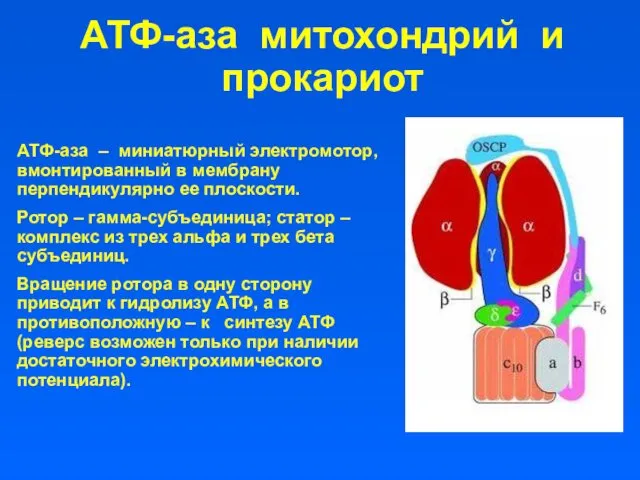
АТФ-аза митохондрий и прокариот
АТФ-аза – миниатюрный электромотор, вмонтированный в мембрану перпендикулярно
АТФ-аза митохондрий и прокариот
АТФ-аза – миниатюрный электромотор, вмонтированный в мембрану перпендикулярно
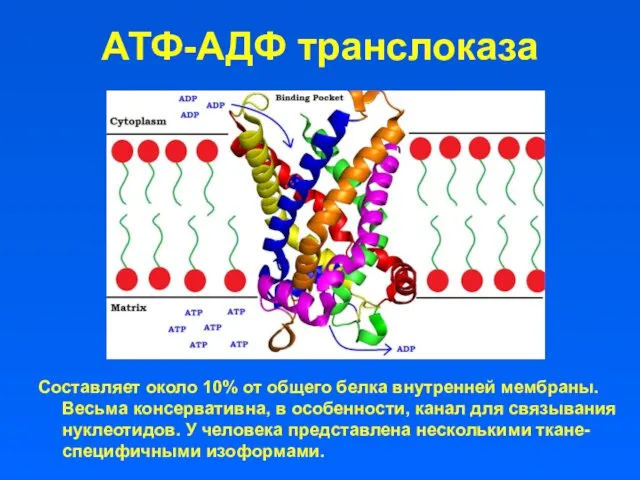
АТФ-АДФ транслоказа
Составляет около 10% от общего белка внутренней мембраны. Весьма консервативна,
АТФ-АДФ транслоказа
Составляет около 10% от общего белка внутренней мембраны. Весьма консервативна,

Хемиосмотическая теория
Цепь переноса электронов переносит протоны из матрикса митохондрии в цитозоль
Хемиосмотическая теория
Цепь переноса электронов переносит протоны из матрикса митохондрии в цитозоль

Генерация энергии митохондриями
Универсальная форма энергии – электрохимический потенциал на мембране (ΔH+
Генерация энергии митохондриями
Универсальная форма энергии – электрохимический потенциал на мембране (ΔH+

Регуляция окислительного фосфорилирования через систему обратных связей
Дыхательный контроль – в норме
Регуляция окислительного фосфорилирования через систему обратных связей
Дыхательный контроль – в норме

Ингибиторы митохондрий
Разобщители – FCCP, 2,4-ДНФ, жирные кислоты. Повышают проницаемость мембраны для
Ингибиторы митохондрий
Разобщители – FCCP, 2,4-ДНФ, жирные кислоты. Повышают проницаемость мембраны для

Митохондрия как терморегулятор
Жирные кислоты – природные разобщители, которые регулируют соотношение окисления
Митохондрия как терморегулятор
Жирные кислоты – природные разобщители, которые регулируют соотношение окисления

Энергетические затраты животной клетки
Работа K/Na–АТФазы – 15-60% энергии (в среднем около
Энергетические затраты животной клетки
Работа K/Na–АТФазы – 15-60% энергии (в среднем около
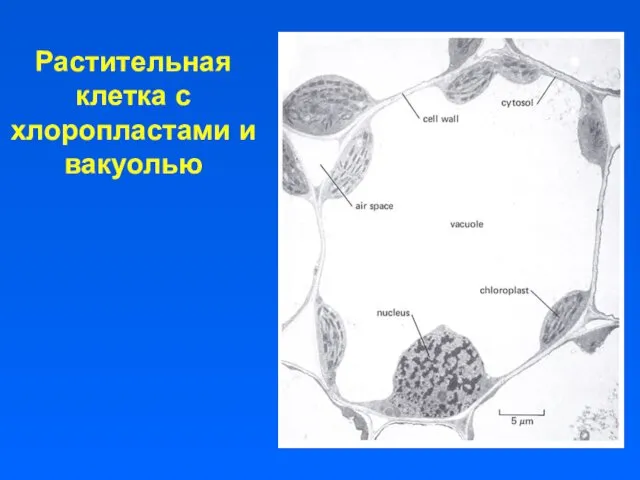
Растительная клетка с хлоропластами и вакуолью
Растительная клетка с хлоропластами и вакуолью
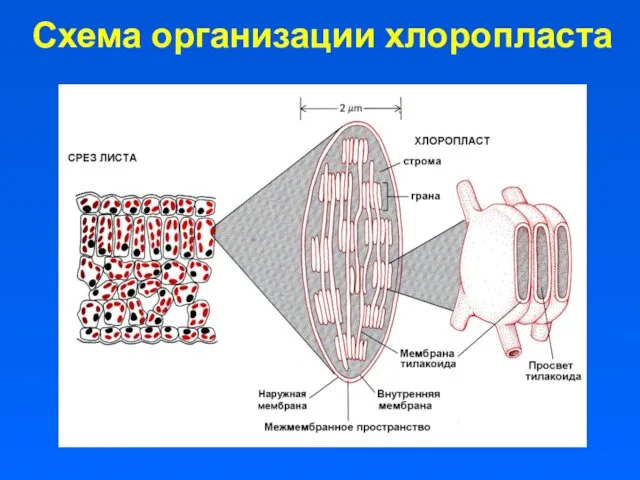
Схема организации хлоропласта
Схема организации хлоропласта
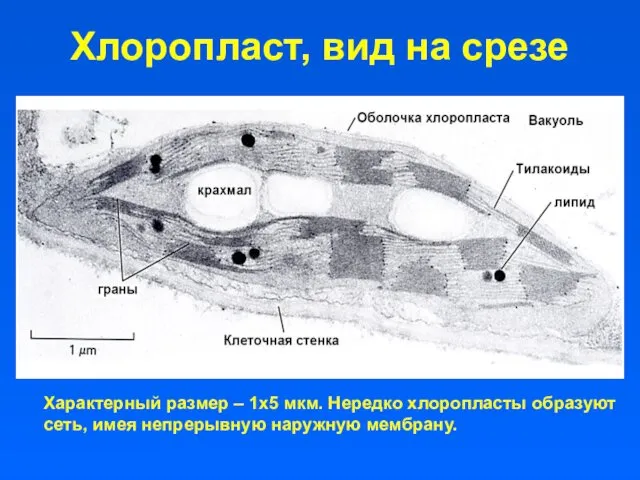
Хлоропласт, вид на срезе
Характерный размер – 1х5 мкм. Нередко хлоропласты образуют
Хлоропласт, вид на срезе
Характерный размер – 1х5 мкм. Нередко хлоропласты образуют
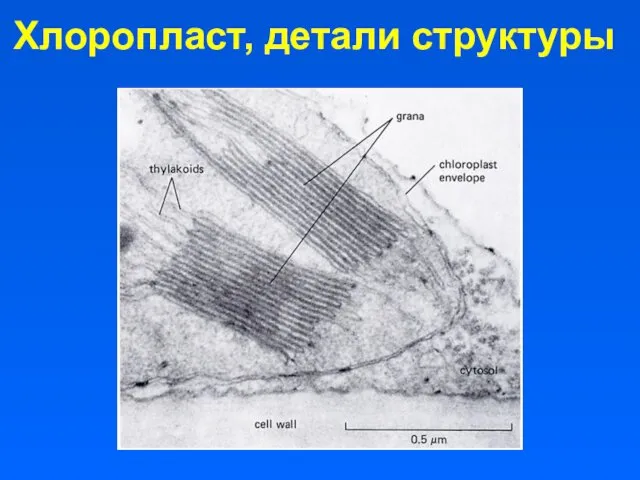
Хлоропласт, детали структуры
Хлоропласт, детали структуры

Мембраны хлоропласта
Наружная мембрана: гладкая; высокопроницаемая.
Внутренняя мембрана: гладкая (не образует крист);
Мембраны хлоропласта
Наружная мембрана: гладкая; высокопроницаемая.
Внутренняя мембрана: гладкая (не образует крист);

Компоненты хлоропласта
Строма имеет щелочной рН. Она содержит нуклеиновые кислоты (ДНК, РНК)
Компоненты хлоропласта
Строма имеет щелочной рН. Она содержит нуклеиновые кислоты (ДНК, РНК)
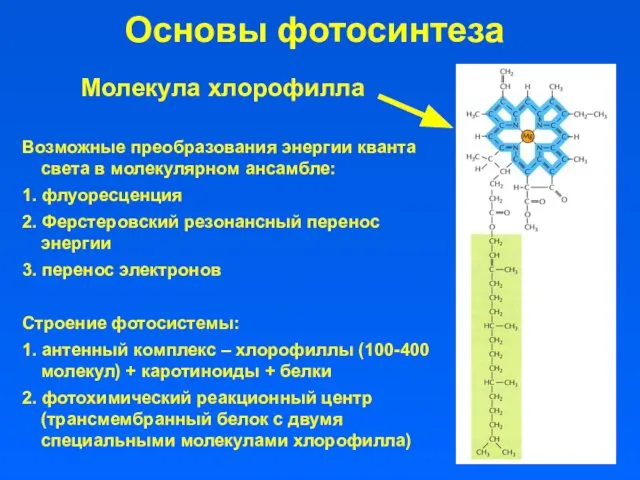
Основы фотосинтеза
Возможные преобразования энергии кванта света в молекулярном ансамбле:
1. флуоресценция
2. Ферстеровский
Основы фотосинтеза
Возможные преобразования энергии кванта света в молекулярном ансамбле:
1. флуоресценция
2. Ферстеровский

Световые реакции фотосинтеза
Фотосистема II – комплекс из примерно 20 белков с
Световые реакции фотосинтеза
Фотосистема II – комплекс из примерно 20 белков с
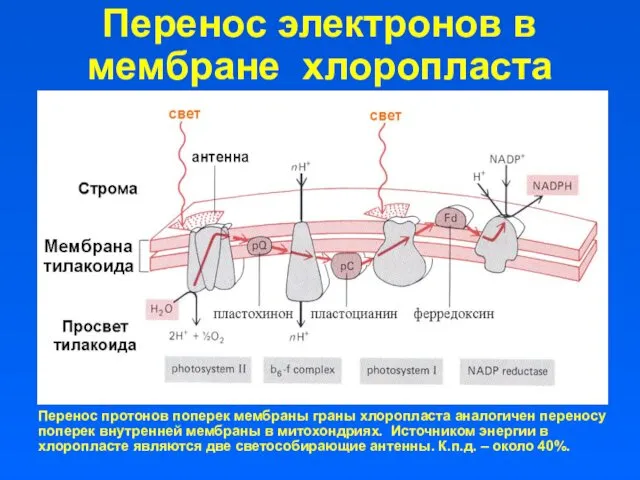
Перенос электронов в мембране хлоропласта
Перенос протонов поперек мембраны граны хлоропласта аналогичен
Перенос электронов в мембране хлоропласта
Перенос протонов поперек мембраны граны хлоропласта аналогичен
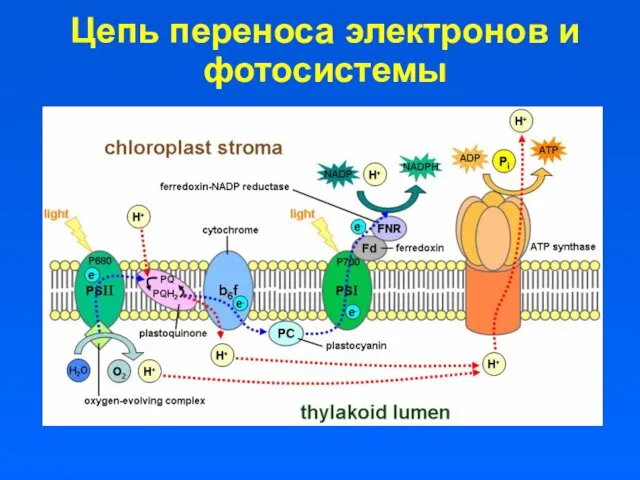
Цепь переноса электронов и фотосистемы
Цепь переноса электронов и фотосистемы
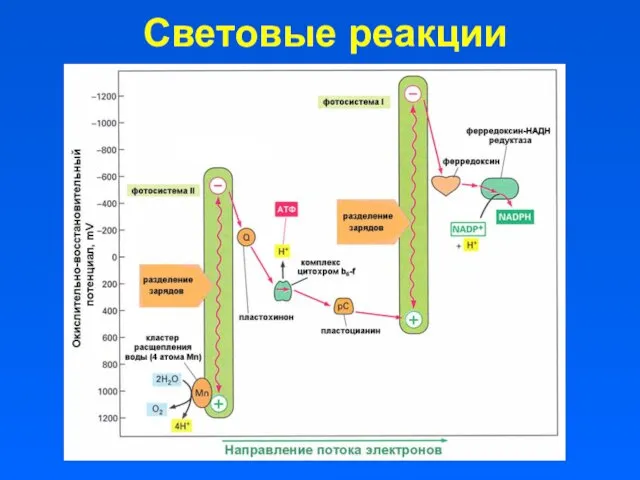
Световые реакции
Световые реакции

Световые и темновые реакции фотосинтеза
Световые реакции – зависят от постоянного потока
Световые и темновые реакции фотосинтеза
Световые реакции – зависят от постоянного потока
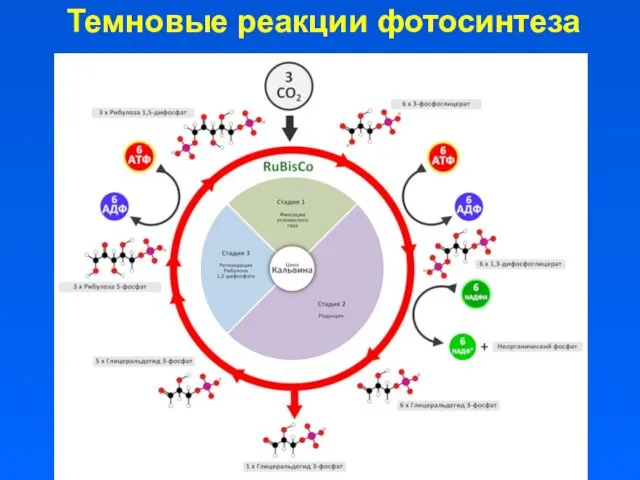
Темновые реакции фотосинтеза
Темновые реакции фотосинтеза

Развитие хлоропластов
Хлоропласты (например, в побегах) развиваются под действием света из пропластид,
Развитие хлоропластов
Хлоропласты (например, в побегах) развиваются под действием света из пропластид,

Дифференцировка хлоропластов
Пропластиды – бесцветные двумембранные структуры, диаметром 0,4-1 мкм, содержащие ДНК
Дифференцировка хлоропластов
Пропластиды – бесцветные двумембранные структуры, диаметром 0,4-1 мкм, содержащие ДНК
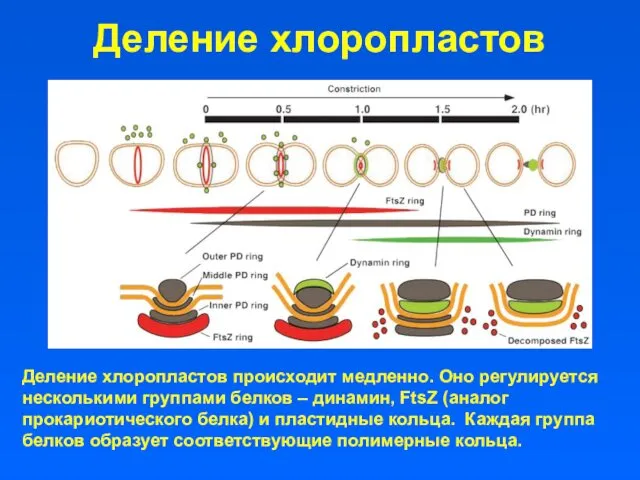
Деление хлоропластов
Деление хлоропластов происходит медленно. Оно регулируется несколькими группами белков –
Деление хлоропластов
Деление хлоропластов происходит медленно. Оно регулируется несколькими группами белков –
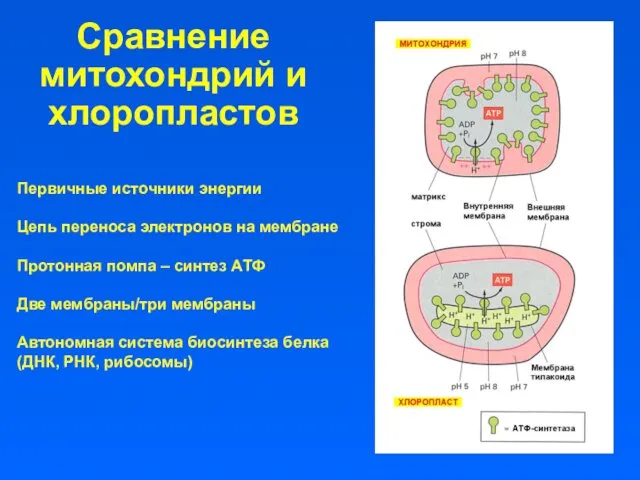
Сравнение митохондрий и хлоропластов
Первичные источники энергии
Цепь переноса электронов на мембране
Протонная
Сравнение митохондрий и хлоропластов
Первичные источники энергии
Цепь переноса электронов на мембране
Протонная

Митохондрии и хлоропласты: нуклеиновые кислоты
Митохондрия: кольцевые молекулы ДНК – 16
Митохондрии и хлоропласты: нуклеиновые кислоты
Митохондрия: кольцевые молекулы ДНК – 16

Митохондрии и хлоропласты: полуавтономность
Митохондрия: может функционировать без собственной ДНК, однако эффективность
Митохондрии и хлоропласты: полуавтономность
Митохондрия: может функционировать без собственной ДНК, однако эффективность
 Презентация на тему Лимфатическая система
Презентация на тему Лимфатическая система  Энеолит. Ботай мәдениеті
Энеолит. Ботай мәдениеті Царство грибы. Разнообразие грибов
Царство грибы. Разнообразие грибов Кто такие рыбы
Кто такие рыбы Полезные и вредные мутации
Полезные и вредные мутации Строение дерева
Строение дерева Морфофункциональная функция характеристики маточной трубы
Морфофункциональная функция характеристики маточной трубы Презентация на тему Железы нашего организма
Презентация на тему Железы нашего организма  Животные домашние и дикие
Животные домашние и дикие Программирование внутриклеточных реакций Белецкий Б. А.
Программирование внутриклеточных реакций Белецкий Б. А. Phylum: Nematoda
Phylum: Nematoda Обитатели водоёмов
Обитатели водоёмов Размножение и развитие насекомых Разработал учитель химии и биологии Петреня Игорь Михайлович
Размножение и развитие насекомых Разработал учитель химии и биологии Петреня Игорь Михайлович Клеточная инженерия эмбрионов
Клеточная инженерия эмбрионов Витамин С (Аскорбиновая кислота)
Витамин С (Аскорбиновая кислота) Абиотические факторы среды
Абиотические факторы среды Презентация на тему "Информационные технологии в биологических исследованиях" - скачать бесплатно презентации по Биологии
Презентация на тему "Информационные технологии в биологических исследованиях" - скачать бесплатно презентации по Биологии Ядовитые животные
Ядовитые животные Презентация на тему "КАК ПИТАЮТСЯ РАСТЕНИЯ? ФОТОСИНТЕЗ" - скачать презентации по Биологии
Презентация на тему "КАК ПИТАЮТСЯ РАСТЕНИЯ? ФОТОСИНТЕЗ" - скачать презентации по Биологии Гельминтология. Размножение и развитие
Гельминтология. Размножение и развитие Изучаем обитателей царства насекомых
Изучаем обитателей царства насекомых Листостебельные мхи
Листостебельные мхи Царство Грибы
Царство Грибы Дигибридное скрещивание Добровольская Юлия,10 а.
Дигибридное скрещивание Добровольская Юлия,10 а. Понятия вид и критерии вида
Понятия вид и критерии вида Lepechin Ivan Ivanovich
Lepechin Ivan Ivanovich Разнообразие экосистем. Сравнительная характеристик а природных и искусственных экосистем
Разнообразие экосистем. Сравнительная характеристик а природных и искусственных экосистем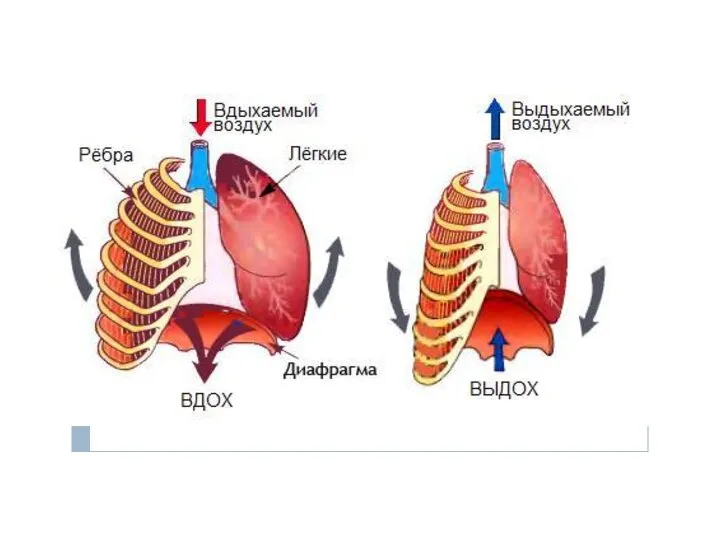 Речевое дыхание
Речевое дыхание