Моделирование регуляции развития меристемы побега в эмбриогенезе Arabidopsis thaliana Научный руководитель: к.б.н., доц. Лихошвай В. А. До
- Моделирование регуляции развития меристемы побега в эмбриогенезе Arabidopsis thaliana Научный руководитель: к.б.н., доц. Лихошвай В. А. До

Содержание
- 2. Объект исследования: Arabidopsis thaliana Arabidopsis был первым растением, геном которого был полностью секвенирован. Существует программа, по
- 3. Меристема побега Arabidopsis thaliana: Новосибирск, 2008 Апикальная меристема побега (АМП) с формирующимися листьями
- 4. Основные гормоны, регулирующие рост и развитие растений: Новосибирск, 2008 Ауксин – основной гормон растений, который регулирует
- 5. Новосибирск, 2008 Цель: Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при развитии меристемы побега
- 6. Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина и ауксинового ответа. Ауксин накапливается
- 7. Химические структуры веществ, обладающих активностью ауксина: Новосибирск, 2008 Ben L. Pederson, 2007
- 8. Метаболизм ауксина: Новосибирск, 2008 Woodward and Bartel, 2005 Потенциальные пути биосинтеза ауксина в клетке Арабидопсиса. Пути
- 9. Конъюгация ауксина: Новосибирск, 2008 Потенциальные пути метаболизма ауксина в клетке Арабидопсиса. Woodward and Bartel, 2005
- 10. Структурная модель генной сети метаболизма ауксина*: Новосибирск, 2008 *-Ananko et al., 2005
- 11. *цитоплазма *ядро *клеточная мембрана *пероксисома *ЭПР *хлоропласт *митохондрия Новосибирск, 2008 Структурная модель генной сети метаболизма ауксина:
- 12. Новосибирск, 2008 Структурная модель генной сети метаболизма ауксина:
- 13. Новосибирск, 2008 Редуцированная структурная модель генной сети метаболизма ауксина:
- 14. MGSgenerator: блок автоматической конвертации формата ГС в формат моделей: Новосибирск, 2008
- 15. MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем: Новосибирск, 2008
- 16. Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега: В клетках побега Арабидопсиса, свободная
- 17. Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке в зависимости от начальной концентрации:
- 18. Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях (trp2,trp3): Расчёты модели качественно совпадают
- 19. Эмбриональное развитие меристемы побега Arabidopsis Thaliana Область моделирования Новосибирск, 2008
- 20. Разработка пространственно – распределённой модели: Основные принципы: Клетки автомата могут обмениваться химическими сигналами. Было выбрана 3
- 21. Типы клеток автомата: Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS и находятся в верхней
- 22. Type – тип клетки BS0, SS0, SD0 – значения сигналов продуцируемых данной клеткой. BS, SS, SD
- 23. Новосибирск, 2008 Темпы делений клеток меристемы побега: Промеристем Л2Меристем Л3Меристем Транзитные Латеральные
- 24. Взаимодействие «клеток» модели: Глобальное взаимодействие Новосибирск, 2008 Суммарное влияние на клетку с координатой (i, j) есть
- 25. Stem signal Differentiation signal Basal signal Визуализация модели клеточного автомата: Новосибирск, 2008
- 26. Результаты моделирования. Нормальное развитие зародыша: Новосибирск, 2008
- 27. Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and Floral Meristem Cell Proliferation and Fate.
- 28. Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки): В развивающейся АМП Нормальное развитие
- 29. Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки): Нормальное развитие Мутация II типа
- 30. Распределение паттернов ауксина в развивающейся апикальной меристеме побега: Иммунная локализация белков PIN1 в эмбрионе: окрашена полярное
- 31. Распределение паттернов ауксина в развивающейся апикальной меристеме побега: Полярная локализация белков PIN1 на более поздней стадии
- 32. Новосибирск, 2008 Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие этапы: 1) автоматическая генерация
- 33. Новосибирск, 2008 Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма ауксина дополнена моделями, описывающими
- 34. Новосибирск, 2008 Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana. Для пространственно-распределённой модели подобран оптимальный
- 36. Скачать презентацию

Объект исследования:
Arabidopsis thaliana
Arabidopsis был первым растением, геном которого был полностью
Объект исследования:
Arabidopsis thaliana
Arabidopsis был первым растением, геном которого был полностью
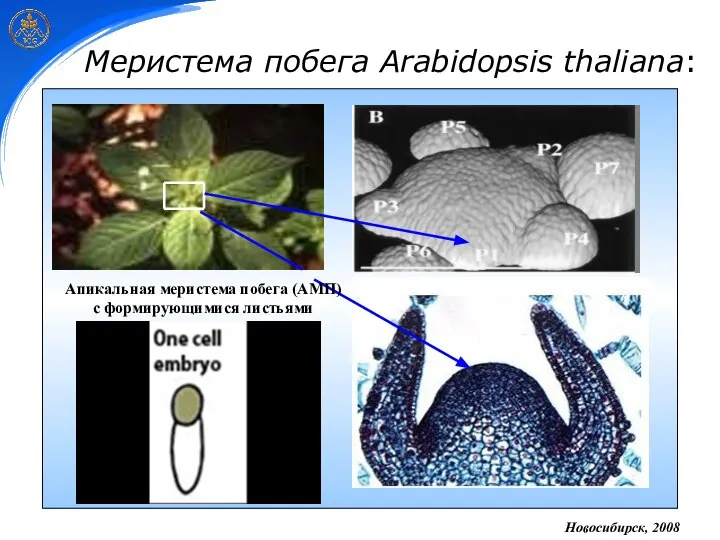
Меристема побега Arabidopsis thaliana:
Новосибирск, 2008
Апикальная меристема побега (АМП)
с формирующимися листьями
Меристема побега Arabidopsis thaliana:
Новосибирск, 2008
Апикальная меристема побега (АМП)
с формирующимися листьями
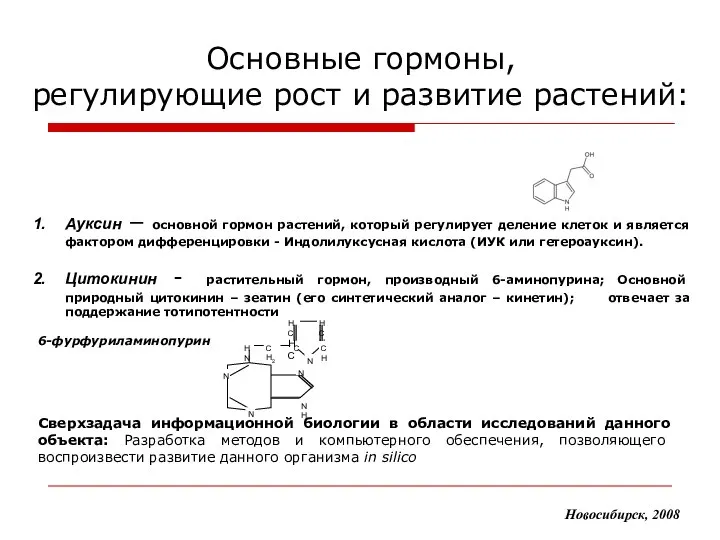
Основные гормоны,
регулирующие рост и развитие растений:
Новосибирск, 2008
Ауксин – основной гормон
Основные гормоны,
регулирующие рост и развитие растений:
Новосибирск, 2008
Ауксин – основной гормон

Новосибирск, 2008
Цель:
Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при
Новосибирск, 2008
Цель:
Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при
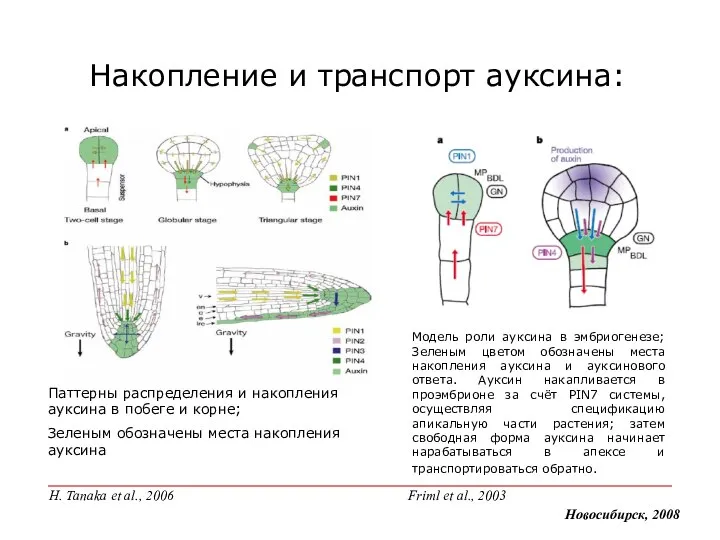
Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина
Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина
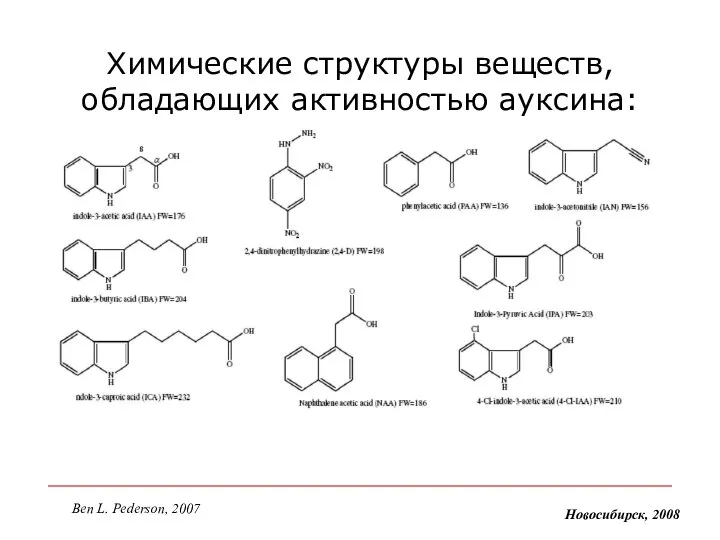
Химические структуры веществ, обладающих активностью ауксина:
Новосибирск, 2008
Ben L. Pederson, 2007
Химические структуры веществ, обладающих активностью ауксина:
Новосибирск, 2008
Ben L. Pederson, 2007
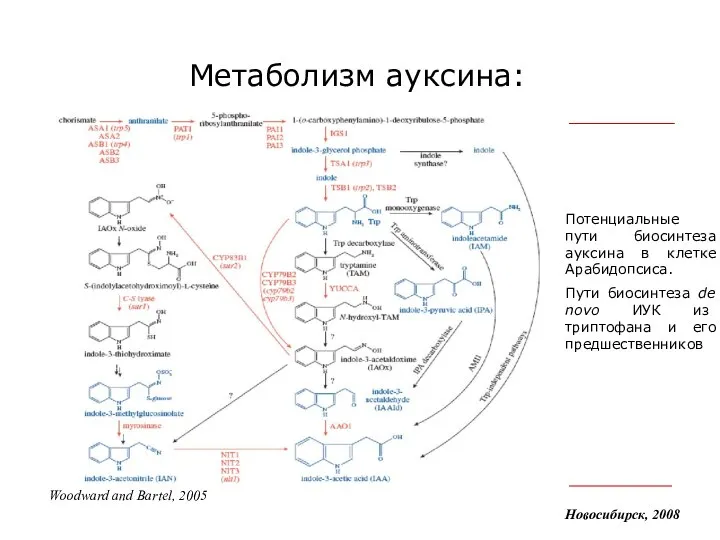
Метаболизм ауксина:
Новосибирск, 2008
Woodward and Bartel, 2005
Потенциальные пути биосинтеза ауксина в клетке
Метаболизм ауксина:
Новосибирск, 2008
Woodward and Bartel, 2005
Потенциальные пути биосинтеза ауксина в клетке
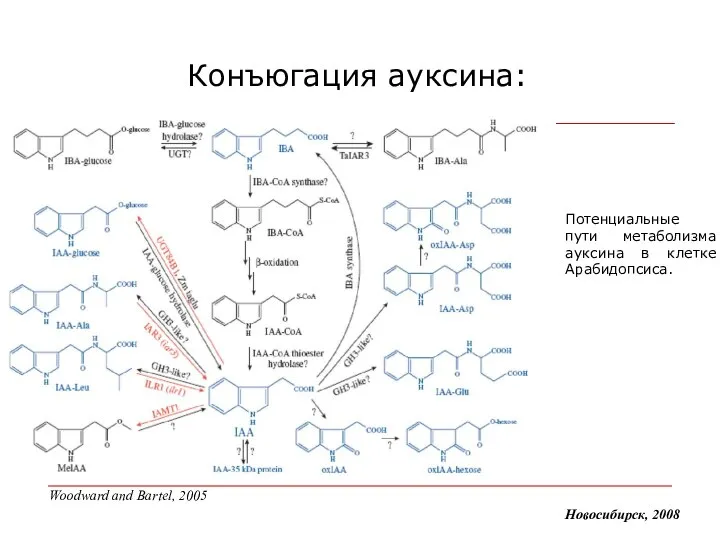
Конъюгация ауксина:
Новосибирск, 2008
Потенциальные пути метаболизма ауксина в клетке Арабидопсиса.
Woodward and Bartel,
Конъюгация ауксина:
Новосибирск, 2008
Потенциальные пути метаболизма ауксина в клетке Арабидопсиса.
Woodward and Bartel,
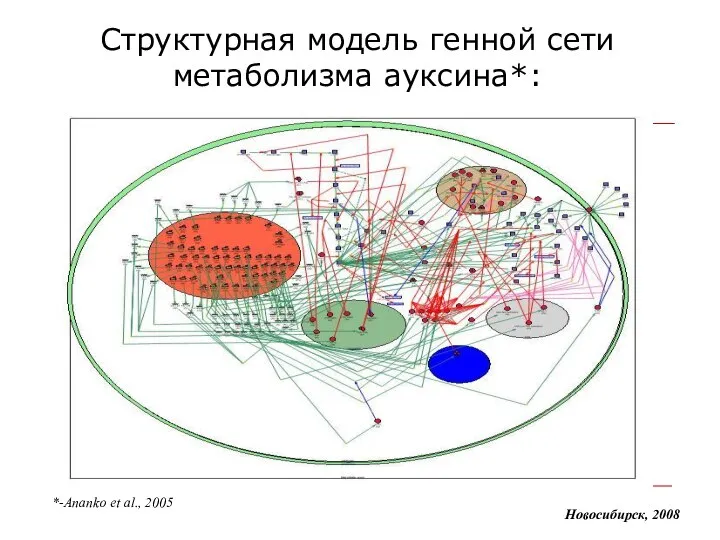
Структурная модель генной сети метаболизма ауксина*:
Новосибирск, 2008
*-Ananko et al., 2005
Структурная модель генной сети метаболизма ауксина*:
Новосибирск, 2008
*-Ananko et al., 2005
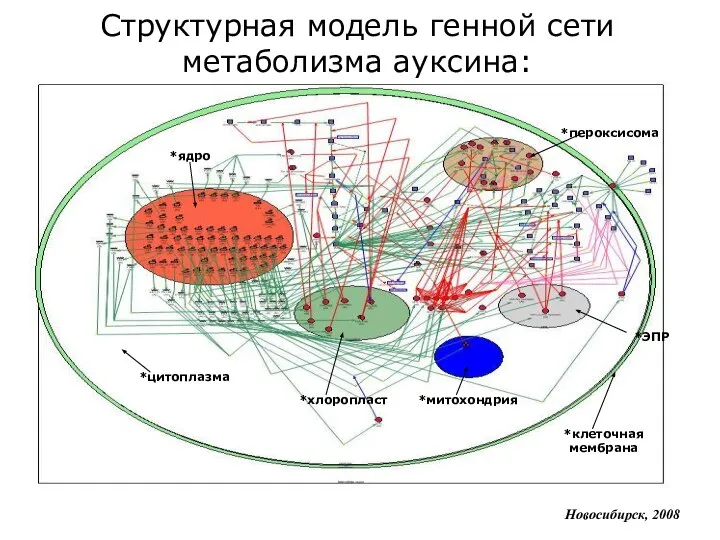
*цитоплазма
*ядро
*клеточная мембрана
*пероксисома
*ЭПР
*хлоропласт
*митохондрия
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
*цитоплазма
*ядро
*клеточная мембрана
*пероксисома
*ЭПР
*хлоропласт
*митохондрия
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
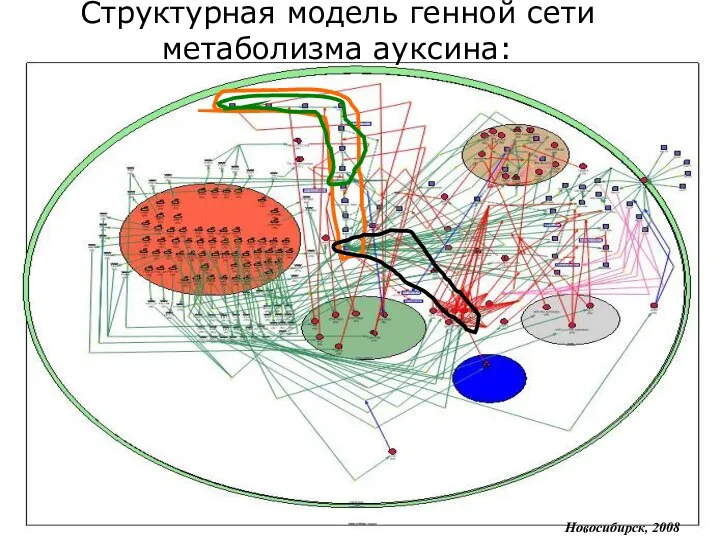
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:

Новосибирск, 2008
Редуцированная структурная модель генной сети метаболизма ауксина:
Новосибирск, 2008
Редуцированная структурная модель генной сети метаболизма ауксина:
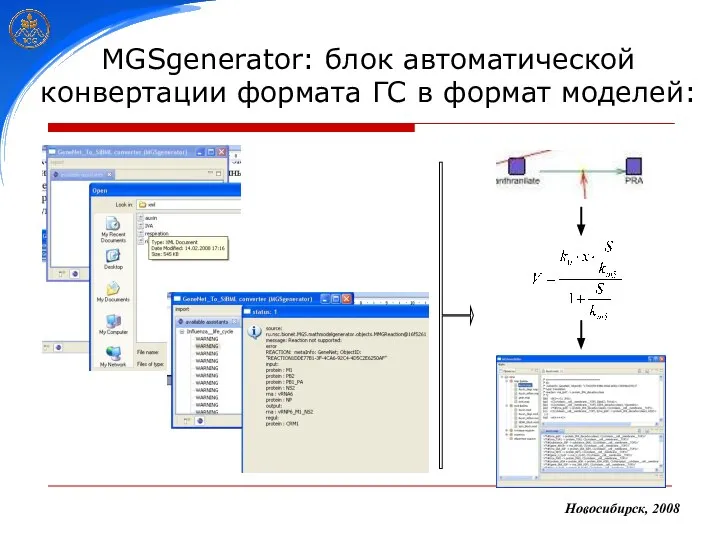
MGSgenerator: блок автоматической конвертации формата ГС в формат моделей:
Новосибирск, 2008
MGSgenerator: блок автоматической конвертации формата ГС в формат моделей:
Новосибирск, 2008
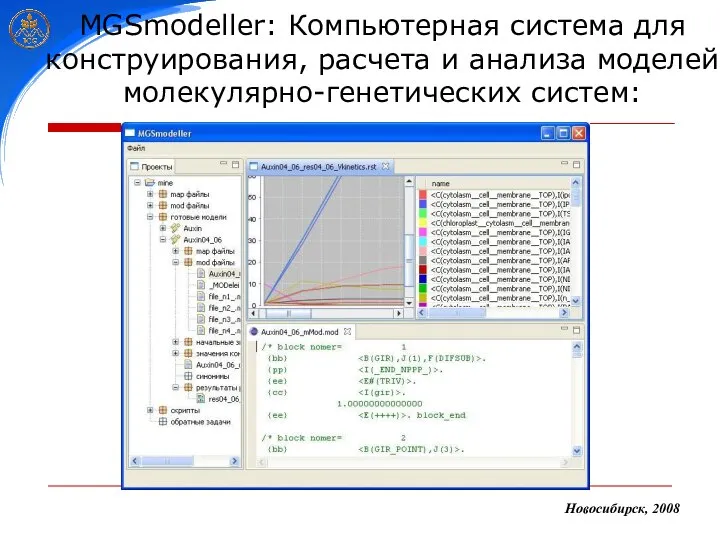
MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем:
Новосибирск,
MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем:
Новосибирск,
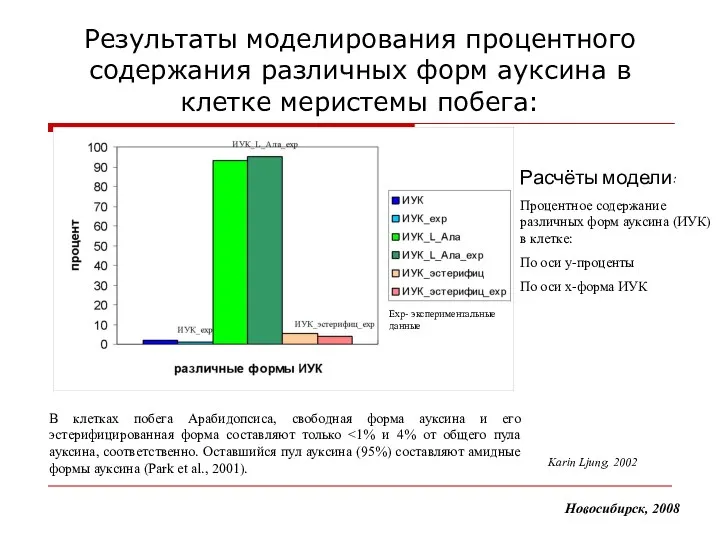
Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега:
В
Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега:
В
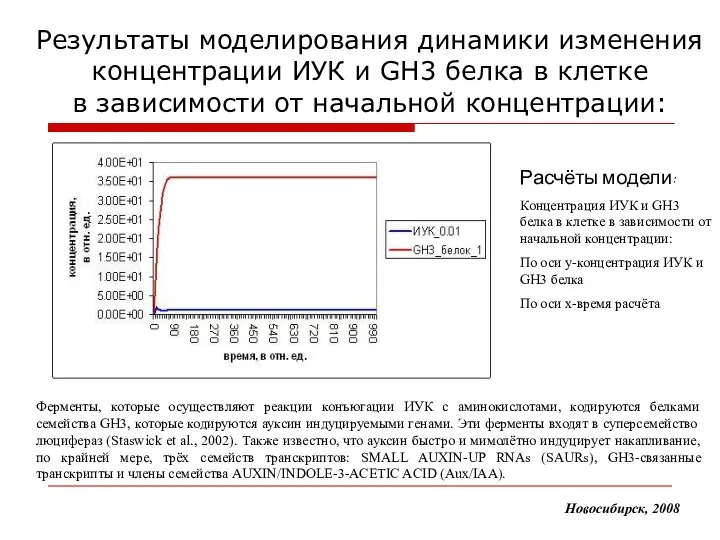
Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке
Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке
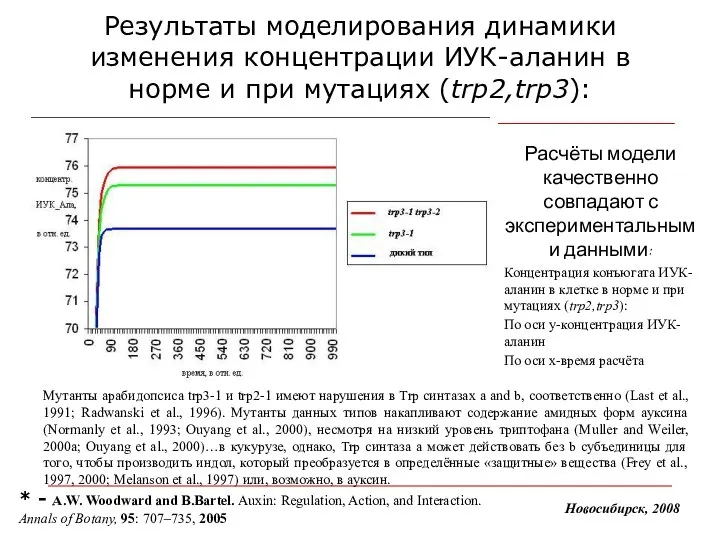
Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях
Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях
Эмбриональное развитие
меристемы побега Arabidopsis Thaliana
Область моделирования
Новосибирск, 2008
Эмбриональное развитие
меристемы побега Arabidopsis Thaliana
Область моделирования
Новосибирск, 2008

Разработка пространственно – распределённой модели:
Основные принципы:
Клетки автомата могут обмениваться химическими сигналами.
Разработка пространственно – распределённой модели:
Основные принципы:
Клетки автомата могут обмениваться химическими сигналами.

Типы клеток автомата:
Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS
Типы клеток автомата:
Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS
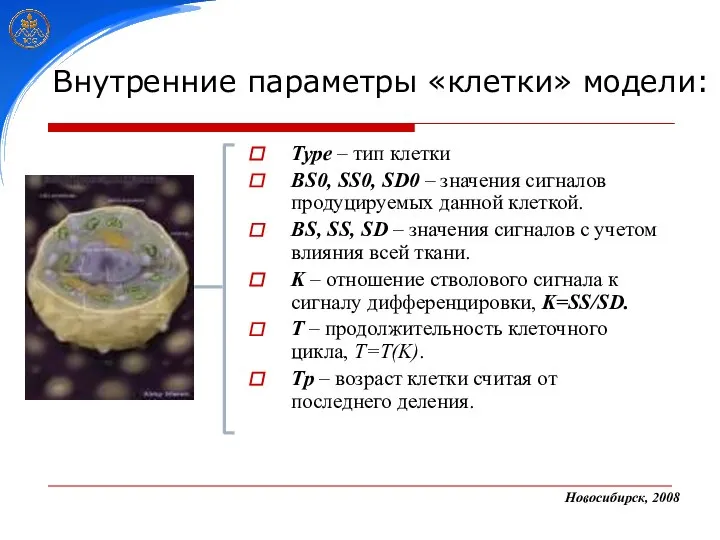
Type – тип клетки
BS0, SS0, SD0 – значения сигналов продуцируемых данной
Type – тип клетки
BS0, SS0, SD0 – значения сигналов продуцируемых данной
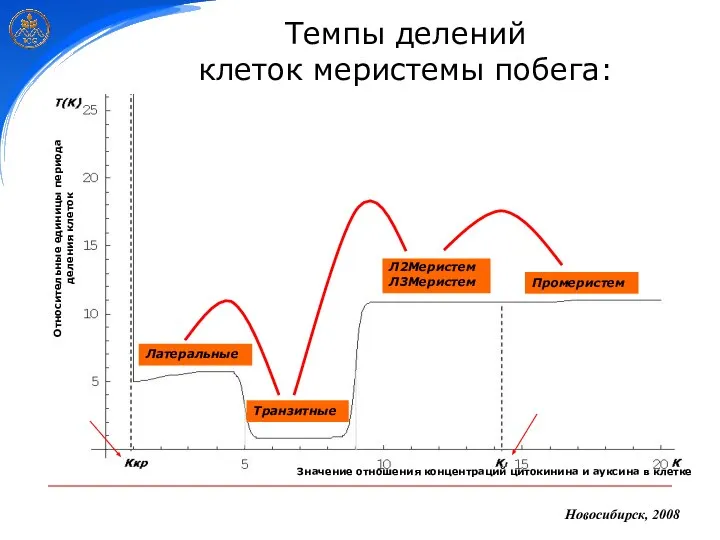
Новосибирск, 2008
Темпы делений
клеток меристемы побега:
Промеристем
Л2Меристем
Л3Меристем
Транзитные
Латеральные
Новосибирск, 2008
Темпы делений
клеток меристемы побега:
Промеристем
Л2Меристем
Л3Меристем
Транзитные
Латеральные
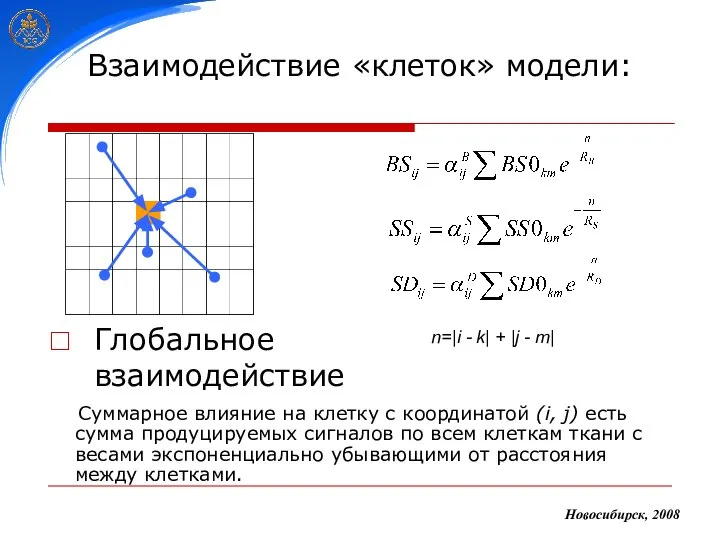
Взаимодействие «клеток» модели:
Глобальное взаимодействие
Новосибирск, 2008
Суммарное влияние на клетку с координатой
Взаимодействие «клеток» модели:
Глобальное взаимодействие
Новосибирск, 2008
Суммарное влияние на клетку с координатой
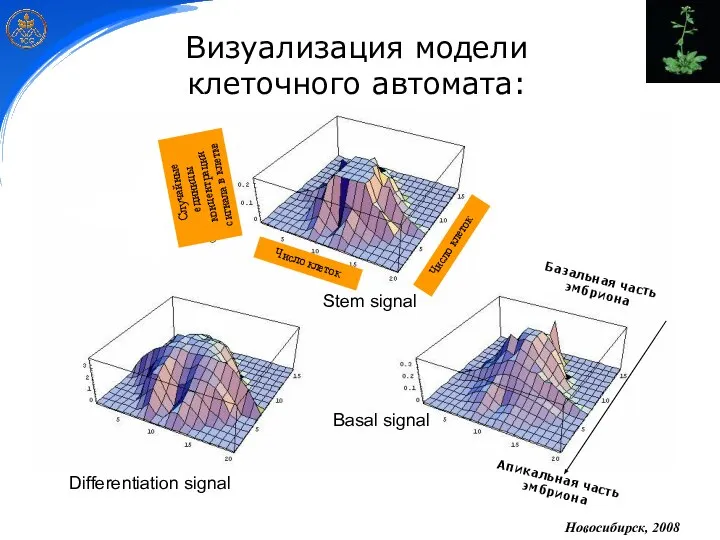
Stem signal
Differentiation signal
Basal signal
Визуализация модели
клеточного автомата:
Новосибирск, 2008
Stem signal
Differentiation signal
Basal signal
Визуализация модели
клеточного автомата:
Новосибирск, 2008
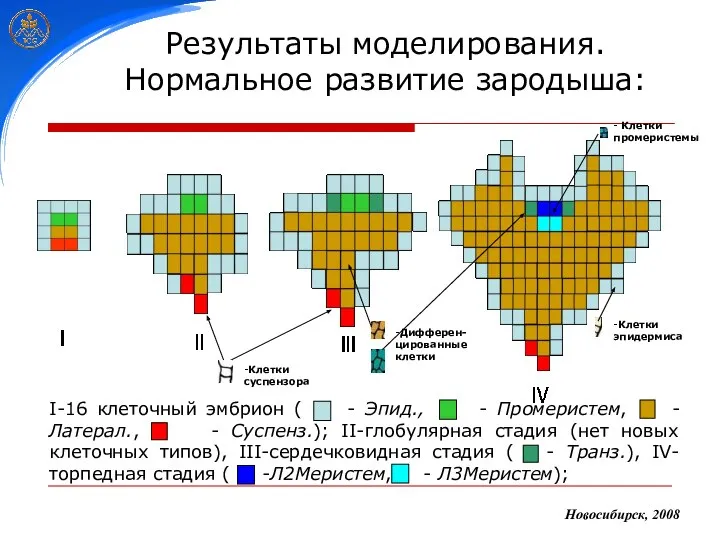
Результаты моделирования.
Нормальное развитие зародыша:
Новосибирск, 2008
Результаты моделирования.
Нормальное развитие зародыша:
Новосибирск, 2008
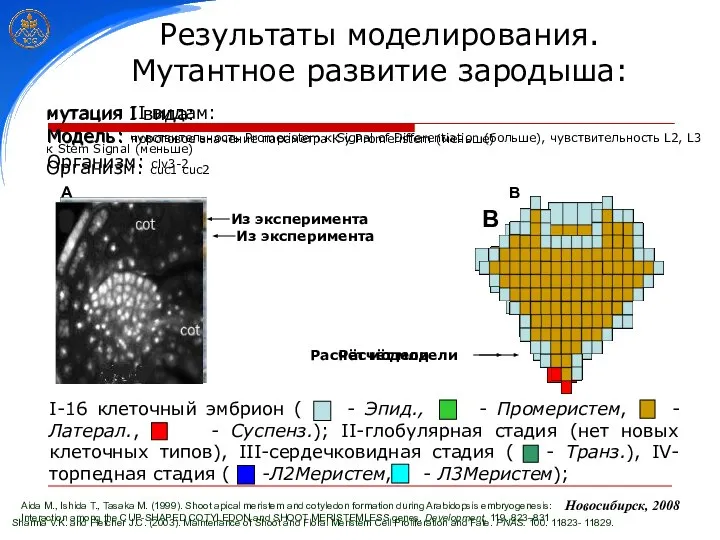
Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and
Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and
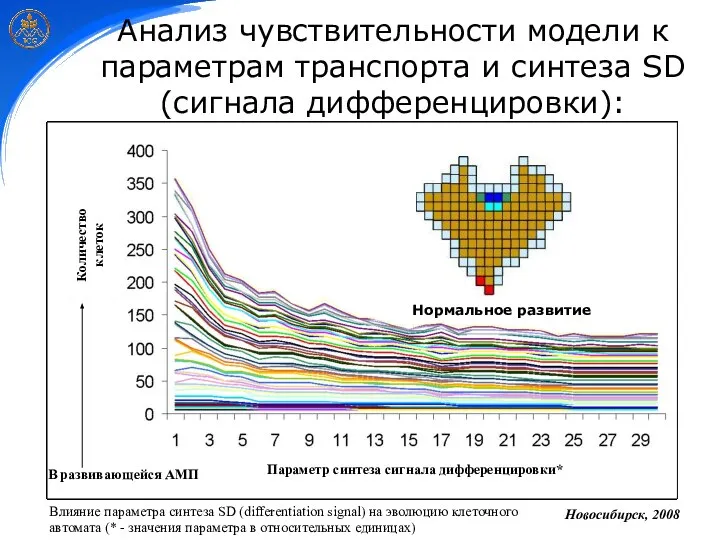
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
В
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
В
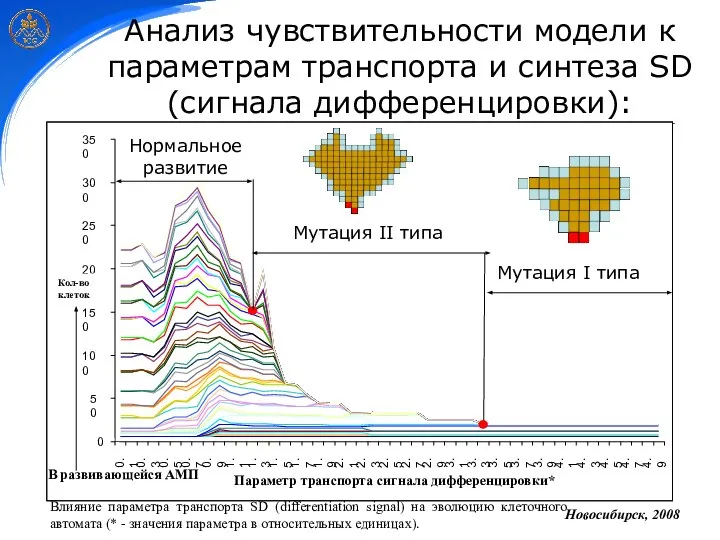
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
Нормальное
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
Нормальное
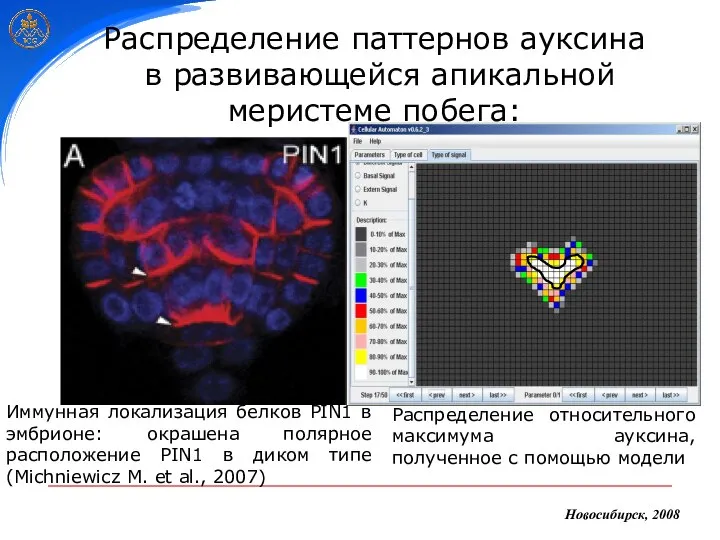
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Иммунная локализация белков PIN1
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Иммунная локализация белков PIN1
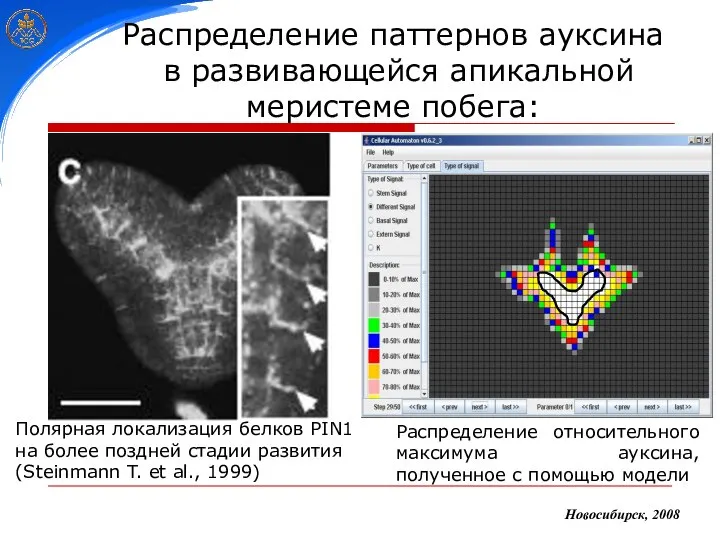
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Полярная локализация белков PIN1
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Полярная локализация белков PIN1

Новосибирск, 2008
Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие
Новосибирск, 2008
Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие

Новосибирск, 2008
Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма
Новосибирск, 2008
Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма

Новосибирск, 2008
Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana.
Новосибирск, 2008
Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana.
 Зачем нужны хвосты?
Зачем нужны хвосты? Опора тела и движение
Опора тела и движение ,Биохимический процесс при занятиях по большому теннису
,Биохимический процесс при занятиях по большому теннису Фотосинтез растений
Фотосинтез растений Семя. Строение семени. Типы семян
Семя. Строение семени. Типы семян Çiçəkli bitkilərdə tozlanma
Çiçəkli bitkilərdə tozlanma Мозг и способы его изучения
Мозг и способы его изучения Презентация на тему Дети Земли
Презентация на тему Дети Земли Тема: Перенос веществ в организме позвоночных животных.
Тема: Перенос веществ в организме позвоночных животных.  Биообъекты-продуценты лечебных, профилактических и диагностических средств
Биообъекты-продуценты лечебных, профилактических и диагностических средств Ферменты: строение, свойства, регуляция активности
Ферменты: строение, свойства, регуляция активности Взаимодействие аллельных и неаллельных генов
Взаимодействие аллельных и неаллельных генов Презентация на тему "ёжик" - скачать бесплатно презентации по Биологии
Презентация на тему "ёжик" - скачать бесплатно презентации по Биологии Конодонты
Конодонты Генетически-модифицированные организмы
Генетически-модифицированные организмы Симпатический отдел вегетативной нервной системы
Симпатический отдел вегетативной нервной системы Роль насекомых в природе
Роль насекомых в природе Проект освоения участка леса для осуществления рекреационного лесопользования
Проект освоения участка леса для осуществления рекреационного лесопользования Презентация на тему "Почва - среда жизни организмов. Состав почвы" - скачать презентации по Биологии
Презентация на тему "Почва - среда жизни организмов. Состав почвы" - скачать презентации по Биологии Сенсорные функции организма. Виды анализаторов
Сенсорные функции организма. Виды анализаторов Опыление и оплодотворение у растений
Опыление и оплодотворение у растений Клетка - структурная единица живого
Клетка - структурная единица живого Единственный способ описать жизнь- это перечислить основные свойства живых организмов: 1)обмен веществ 2) самовоспроизведение 3)н
Единственный способ описать жизнь- это перечислить основные свойства живых организмов: 1)обмен веществ 2) самовоспроизведение 3)н Презентация на тему "Дрожжи" - скачать презентации по Биологии
Презентация на тему "Дрожжи" - скачать презентации по Биологии Обитатели Чёрного моря
Обитатели Чёрного моря Бесполое и половое размножение
Бесполое и половое размножение Многообразие и значение голосеменных
Многообразие и значение голосеменных Органы дыхания и газообмен
Органы дыхания и газообмен