- Непрерывное культивирование микроорганизмов

Содержание
- 2. непрерывное культивирование более перспективно Сущность: - в ферментаторе поддерживаются постоянные условия среды, - микроорганизмы остаются в
- 3. Если для культивирования продуцента используется один ферментатор - гомогенно-непрерывный процесс. Если используется батарея ферментаторов - это
- 4. При непрерывном культивировании микроорганизмов отсутствует смена фаз развития культуры. Скорость потока питательной среды и отвода культуральной
- 5. Две разновидности непрерывных процессов - тубулярные; питательная среда и посевной материал непрерывно поступают в аппарат, в
- 6. Тубулярный процесс Жидкость на входе в аппарат смешивается с посевным материалом. По мере их продвижения в
- 7. Тубулярный процесс Схема тубулярного непрерывного процесса ферментации
- 8. Тубулярный процесс Время движения жидкости t1 от входа в аппарат до любого сечения по длине потока
- 9. Тубулярный процесс Тубулярный процесс с учетом закономерностей протекания процесса ферментации полностью подобен периодическому. Сложность: в непрерывной
- 10. Тубулярный процесс Схема тубулярного непрерывного процесса с рециркуляцией посевного материала
- 11. Тубулярный процесс Преимуществом тубулярного процесса является возможность более полного исчерпания субстрата (как и в периодическом процессе).
- 12. Хемостатный процесс непрерывного культивирования Ферментация протекает в аппарате с мешалкой; В аппарат с постоянной скоростью подается
- 13. схема хемостатного непрерывного процесса ферментации
- 14. Посевной материал на вход в аппарат не подают непрерывно. Засев производится единовременно при запуске культуры от
- 15. Допущения для математической модели: μ = μm(S/KS + S); (по Моно) μ = 0 (отсутствует диссимиляция
- 16. Выражение для материального баланса в ферментере V dX/dt = μmS/(KS+S)· XV – FX; V dS/dt =
- 17. систему уравнений: V dS/dt = FS0 – 1/YXS · μmS/(KS+S) · XV – FS; V dX/dt
- 18. В непрерывном хемостатном процессе довольно быстро устанавливается стационарное состояние, при котором - скорость роста биомассы будет
- 19. Решая при этих условиях уравнение баланса биомассы, получаем: μ X – DX = 0 и μ
- 20. 2 X переходные процессы, возникающие в непрерывном хемостатном процессе при изменении скорости разбавления или удельной скорости
- 21. Из уравнения dX/dt = μX – DX следует, что если μ>D, то скорость роста биомассы в
- 22. в хемостате осуществляется саморегулирование Если величина μ При этом новое установившееся состояние будет при более низкой
- 23. Определение концентрации субстрата и биомассы в установившемся процессе в хемостате С учетом μ X – DX
- 24. Графические зависимости между установившимися значениями X и S в хемостатном процессе и скоростью разбавления D называют
- 25. Хемостатная кривая X(D) имеет особеннность: концентрация биомассы снижается до нуля при повышении скорости разбавления до некоторого
- 26. Отсюда следует, что вид хемостатной кривой зависит от S0. При D = 0, X = YXSS0
- 27. Интересную особенность имеет зависимость стационарной (остаточной) концентрации субстрата от начальной (входной) концентрации. В уравнении S =
- 28. парадоксальный вывод: при любом изменении концентрации субстрата во входящем потоке S0 в стационарном состоянии при заданной
- 29. Что регулируется оператором? - скорость разбавления; (поэтому процесс иногда называют «спидостат»). Возникает вопрос: к чему стремиться?
- 30. Практический интерес представляет определение условий (скорости разбавления) при которых величина будет оптимальной.
- 31. производительность по биомассе в хемостатном процессе сначала возрастает до Donт, а затем резко падает Зависимость производитель-
- 32. СРАВНЕНИЕ ПРОИЗВОДИТЕЛЬНОСТИ ПЕРИОДИЧЕСКОГО И НЕПРЕРЫВНОГО ПРОЦЕССОВ Предположим, что после загрузки и до самого конца процесса биомасса
- 33. Рост биомассы описывается уравнением:
- 34. Предположим, что рост идет до некой максимальной для данного аппарата концентрации биомассы Хт (концентрация биомассы не
- 35. картина роста биомассы при принятых допущениях
- 36. Подставляя Х= Хт и t = tК в уравнение получаем:
- 37. Для непрерывного процесса в том же аппарате производительность по биомассе определяется по другому. Чтобы найти максимально
- 38. Но это возрастание также не может быть бесконечным, пределом выступают массообменные возможности аппарата. Работу можно проводить
- 39. подставляя Определим соотношение Qнепр/Qпериод: в уравнение: Dопт = μm [1 - √KS/(S0 + KS)], получаем Dопт
- 40. В начале процесса инокулят обычно составляет по объему около 5 % от объема среды, а концентрация
- 41. примем t0 = 10 часов (это время подготовки к следующей ферментации). Для быстро растущих культур (бактерии,
- 42. ОТКЛОНЕНИЯ ОТ ТЕОРИИ ХЕМОСТАТА ПРИ ЛИМИТИРОВАНИИ ПРОЦЕССА РАЗЛИЧНЫМИ СУБСТРАТАМИ Хемостатные кривые X(D) чувствительны к изменению кинетических
- 43. Если решить систему уравнений: dX/dt = μmS/(KS+S) - DX dS/dt = D (S0-S) – 1/YXS μX;
- 44. вид хемостатной кривой при лимитировании азотом или серой
- 45. Азот или сера. Наблюдаемые отклонения отражают превышение концентрации биомассы при малых скоростях разбавления по сравнению с
- 46. Магний, фосфор, калий. По виду хемостатная кривая похожа на кривую для азота, но причина подобного отклонения
- 47. Сложная среда. На неопределенных по составу питательных средах происходит при различных скоростях разбавления изменение лимитирования (переход
- 48. Пристеночный рост биомассы. Хемостатная кривая в этом случае имеет аномально длинный «хвост», рост биомассы возможен даже
- 49. ХЕМОСТАТНЫЙ ПРОЦЕСС КУЛЬТИВИРОВАНИЯ, ЛИМИТИРОВАННЫЙ КИСЛОРОДОМ Кинетика роста аэробных культур микроорганизмов зависит от двух субстратов. Вторым является
- 50. Если ферментационное оборудование недостаточно интенсивное, эта зависимость вырождается в однофакторную зависимость от концентрации растворенного кислорода С:
- 51. Из условия стационарности dX/dt = О можно найти значение установившейся концентрации растворенного кислорода:
- 52. Последним членом уравнения в связи с низкой растворимостью кислорода в жидкости можно пренебречь. Для стационарного состояния
- 53. отсюда концентрация негазообразного субстрата S, хотя и не лимитирует процесс, также зависит от D или производительность
- 54. производительность существенно зависит от величин KLa и С*. Критическая скорость разбавления определяется по выражению:
- 55. При некотором достаточно малом значении скорости разбавления концентрация негазообразного субстрата снижается настолько, что начинает лимитировать рост
- 56. в этом случае оптимальной скоростью разбавления является точка переключения лимитирования с растворенного в жидкости субстрата S
- 57. АВТОСЕЛЕКЦИЯ В НЕПРЕРЫВНОМ ПРОЦЕССЕ В ходе процесса культивирования под воздействием космических лучей и различных мутагенных факторов
- 58. изменение концентрации остаточного субстрата в непрерывном процессе при замещении штамма 1 штаммом 2
- 59. Если процесс протекает при скорости разбавления D1 то исходный штамм обеспечивает саморегулирование процесса при концентрации S1.
- 60. Рост происходит до тех пор, пока концентрация субстрата не снизится до S2. При этом исходный штамм
- 61. внешнее проявление автоселекции в непрерывном процессе
- 63. Скачать презентацию

непрерывное культивирование более перспективно
Сущность:
- в ферментаторе поддерживаются постоянные условия
непрерывное культивирование более перспективно
Сущность:
- в ферментаторе поддерживаются постоянные условия

Если для культивирования продуцента используется один ферментатор - гомогенно-непрерывный процесс.
Если
Если для культивирования продуцента используется один ферментатор - гомогенно-непрерывный процесс.
Если

При непрерывном культивировании микроорганизмов отсутствует смена фаз развития культуры.
Скорость потока питательной среды
При непрерывном культивировании микроорганизмов отсутствует смена фаз развития культуры.
Скорость потока питательной среды

Две разновидности непрерывных процессов
- тубулярные;
питательная среда и посевной материал непрерывно поступают
Две разновидности непрерывных процессов
- тубулярные;
питательная среда и посевной материал непрерывно поступают

Тубулярный процесс
Жидкость на входе в аппарат смешивается с посевным материалом.
По мере
Тубулярный процесс
Жидкость на входе в аппарат смешивается с посевным материалом.
По мере
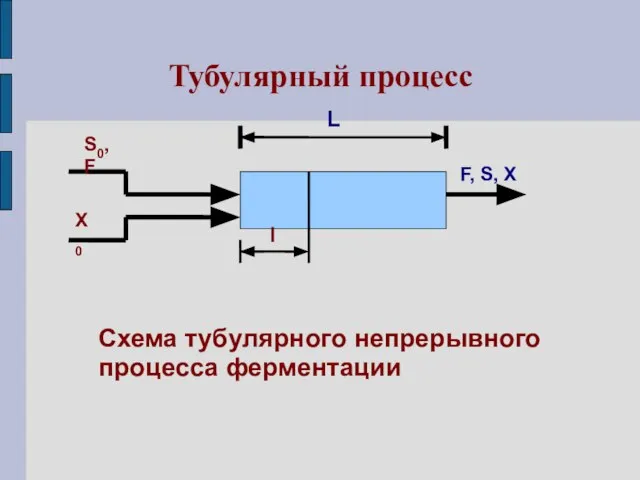
Тубулярный процесс
Схема тубулярного непрерывного
процесса ферментации
Тубулярный процесс
Схема тубулярного непрерывного
процесса ферментации

Тубулярный процесс
Время движения жидкости t1 от входа в аппарат до любого
Тубулярный процесс
Время движения жидкости t1 от входа в аппарат до любого

Тубулярный процесс
Тубулярный процесс с учетом закономерностей протекания процесса ферментации полностью подобен
Тубулярный процесс
Тубулярный процесс с учетом закономерностей протекания процесса ферментации полностью подобен
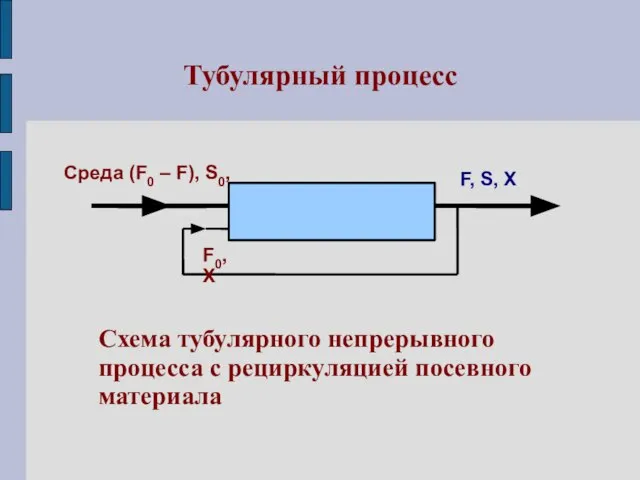
Тубулярный процесс
Схема тубулярного непрерывного
процесса с рециркуляцией посевного
материала
Тубулярный процесс
Схема тубулярного непрерывного
процесса с рециркуляцией посевного
материала

Тубулярный процесс
Преимуществом тубулярного процесса является возможность более полного исчерпания субстрата (как
Тубулярный процесс
Преимуществом тубулярного процесса является возможность более полного исчерпания субстрата (как

Хемостатный процесс непрерывного культивирования
Ферментация протекает в аппарате с мешалкой;
В аппарат с
Хемостатный процесс непрерывного культивирования
Ферментация протекает в аппарате с мешалкой;
В аппарат с
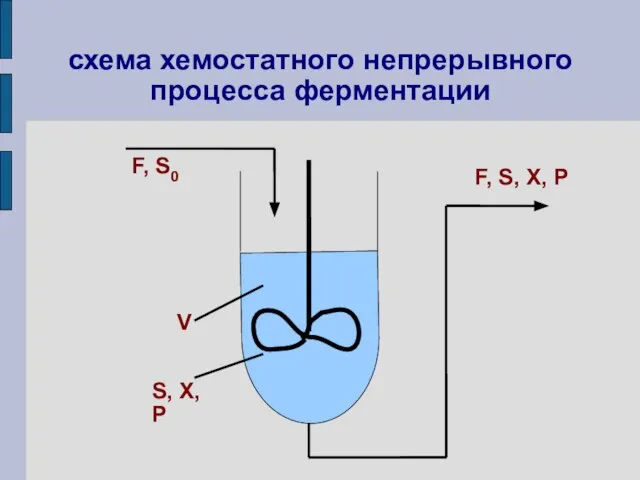
схема хемостатного непрерывного процесса ферментации
схема хемостатного непрерывного процесса ферментации

Посевной материал на вход в аппарат не подают непрерывно.
Засев
Посевной материал на вход в аппарат не подают непрерывно.
Засев

Допущения для математической модели:
μ = μm(S/KS + S); (по Моно)
μ =
Допущения для математической модели:
μ = μm(S/KS + S); (по Моно)
μ =

Выражение для материального баланса в ферментере
V dX/dt = μmS/(KS+S)· XV
Выражение для материального баланса в ферментере
V dX/dt = μmS/(KS+S)· XV

систему уравнений:
V dS/dt = FS0 – 1/YXS · μmS/(KS+S) ·
систему уравнений:
V dS/dt = FS0 – 1/YXS · μmS/(KS+S) ·

В непрерывном хемостатном процессе довольно быстро устанавливается стационарное состояние,
при котором
В непрерывном хемостатном процессе довольно быстро устанавливается стационарное состояние,
при котором

Решая при этих условиях уравнение баланса биомассы, получаем:
μ X – DX
Решая при этих условиях уравнение баланса биомассы, получаем:
μ X – DX
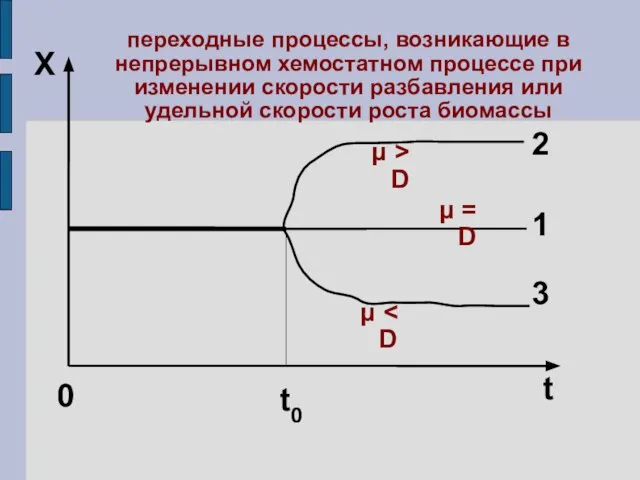
2
X
переходные процессы, возникающие в непрерывном хемостатном процессе при изменении скорости разбавления
2
X
переходные процессы, возникающие в непрерывном хемостатном процессе при изменении скорости разбавления

Из уравнения dX/dt = μX – DX следует, что если μ>D,
Из уравнения dX/dt = μX – DX следует, что если μ>D,

в хемостате осуществляется саморегулирование
Если величина μ < D, то dX/dt <
в хемостате осуществляется саморегулирование
Если величина μ < D, то dX/dt <

Определение концентрации субстрата и биомассы в установившемся процессе в хемостате
С учетом
Определение концентрации субстрата и биомассы в установившемся процессе в хемостате
С учетом
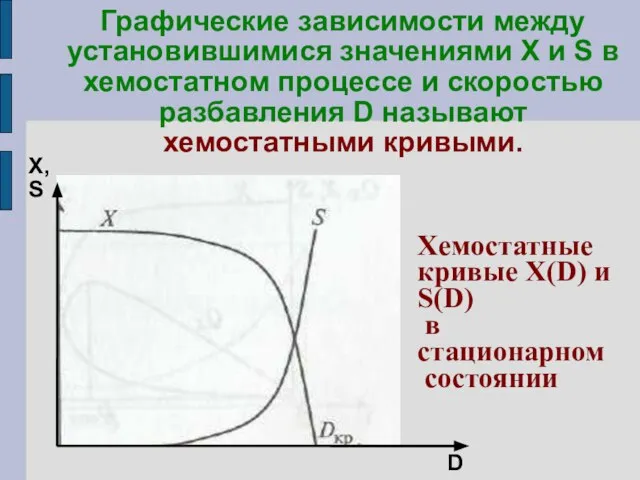
Графические зависимости между установившимися значениями X и S в хемостатном процессе
Графические зависимости между установившимися значениями X и S в хемостатном процессе

Хемостатная кривая X(D) имеет особеннность: концентрация биомассы снижается до нуля при
Хемостатная кривая X(D) имеет особеннность: концентрация биомассы снижается до нуля при

Отсюда следует, что вид хемостатной кривой зависит от S0.
При D
Отсюда следует, что вид хемостатной кривой зависит от S0.
При D

Интересную особенность имеет зависимость стационарной (остаточной) концентрации субстрата от начальной (входной)
Интересную особенность имеет зависимость стационарной (остаточной) концентрации субстрата от начальной (входной)

парадоксальный вывод: при любом изменении концентрации субстрата во входящем потоке S0
парадоксальный вывод: при любом изменении концентрации субстрата во входящем потоке S0

Что регулируется оператором?
- скорость разбавления; (поэтому процесс иногда называют «спидостат»).
Возникает вопрос:
Что регулируется оператором?
- скорость разбавления; (поэтому процесс иногда называют «спидостат»).
Возникает вопрос:

Практический интерес представляет определение условий (скорости разбавления) при которых величина будет
Практический интерес представляет определение условий (скорости разбавления) при которых величина будет
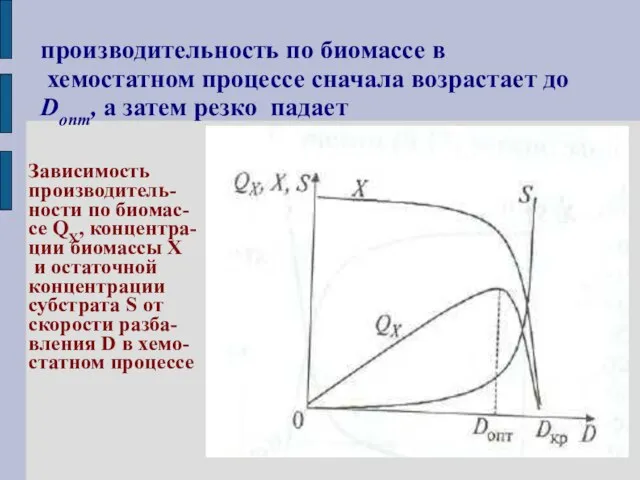
производительность по биомассе в
хемостатном процессе сначала возрастает до Donт, а
производительность по биомассе в хемостатном процессе сначала возрастает до Donт, а

СРАВНЕНИЕ ПРОИЗВОДИТЕЛЬНОСТИ ПЕРИОДИЧЕСКОГО И НЕПРЕРЫВНОГО ПРОЦЕССОВ
Предположим, что после загрузки
СРАВНЕНИЕ ПРОИЗВОДИТЕЛЬНОСТИ ПЕРИОДИЧЕСКОГО И НЕПРЕРЫВНОГО ПРОЦЕССОВ
Предположим, что после загрузки

Рост биомассы описывается уравнением:
Рост биомассы описывается уравнением:

Предположим, что рост идет до некой максимальной для данного аппарата
Предположим, что рост идет до некой максимальной для данного аппарата
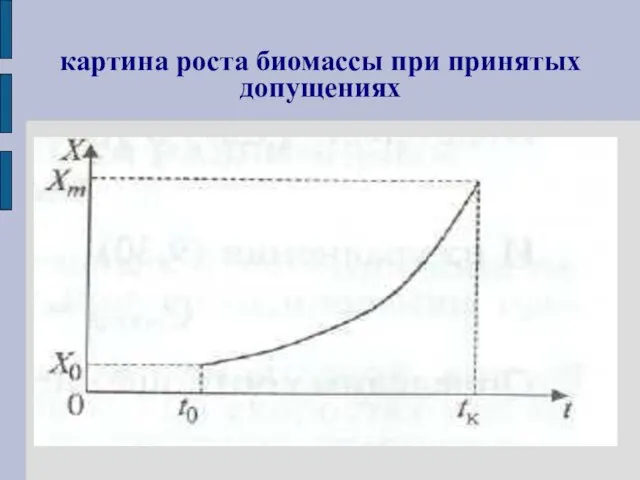
картина роста биомассы при принятых допущениях
картина роста биомассы при принятых допущениях

Подставляя Х= Хт и t = tК в уравнение
получаем:
Подставляя Х= Хт и t = tК в уравнение
получаем:

Для непрерывного процесса в том же аппарате производительность по биомассе определяется
Для непрерывного процесса в том же аппарате производительность по биомассе определяется

Но это возрастание также не может быть бесконечным, пределом выступают
Но это возрастание также не может быть бесконечным, пределом выступают

подставляя
Определим соотношение Qнепр/Qпериод:
в уравнение: Dопт = μm [1 - √KS/(S0 +
подставляя
Определим соотношение Qнепр/Qпериод:
в уравнение: Dопт = μm [1 - √KS/(S0 +

В начале процесса инокулят обычно составляет по объему около 5
В начале процесса инокулят обычно составляет по объему около 5

примем t0 = 10 часов (это время подготовки к следующей ферментации).
Для
примем t0 = 10 часов (это время подготовки к следующей ферментации).
Для

ОТКЛОНЕНИЯ ОТ ТЕОРИИ ХЕМОСТАТА
ПРИ ЛИМИТИРОВАНИИ ПРОЦЕССА РАЗЛИЧНЫМИ СУБСТРАТАМИ
Хемостатные кривые X(D)
ОТКЛОНЕНИЯ ОТ ТЕОРИИ ХЕМОСТАТА
ПРИ ЛИМИТИРОВАНИИ ПРОЦЕССА РАЗЛИЧНЫМИ СУБСТРАТАМИ
Хемостатные кривые X(D)
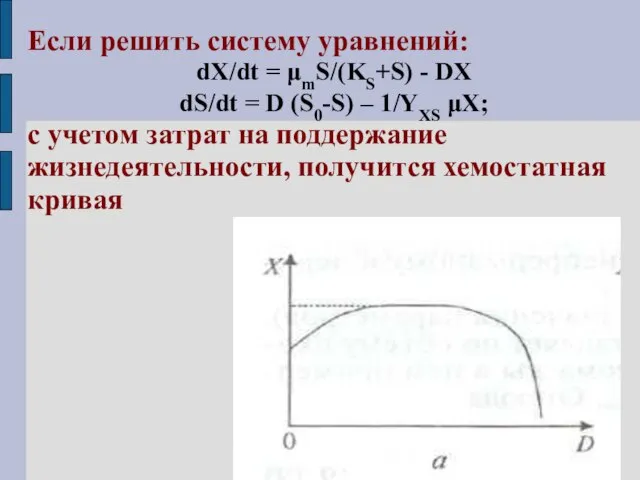
Если решить систему уравнений:
dX/dt = μmS/(KS+S) - DX
dS/dt = D (S0-S)
Если решить систему уравнений:
dX/dt = μmS/(KS+S) - DX
dS/dt = D (S0-S)

вид хемостатной кривой при лимитировании азотом или серой
вид хемостатной кривой при лимитировании азотом или серой

Азот или сера. Наблюдаемые отклонения отражают превышение концентрации биомассы при малых
Азот или сера. Наблюдаемые отклонения отражают превышение концентрации биомассы при малых
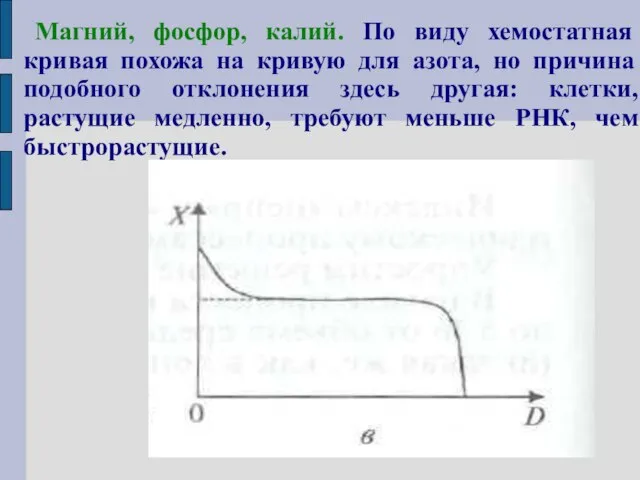
Магний, фосфор, калий. По виду хемостатная кривая похожа на кривую для
Магний, фосфор, калий. По виду хемостатная кривая похожа на кривую для
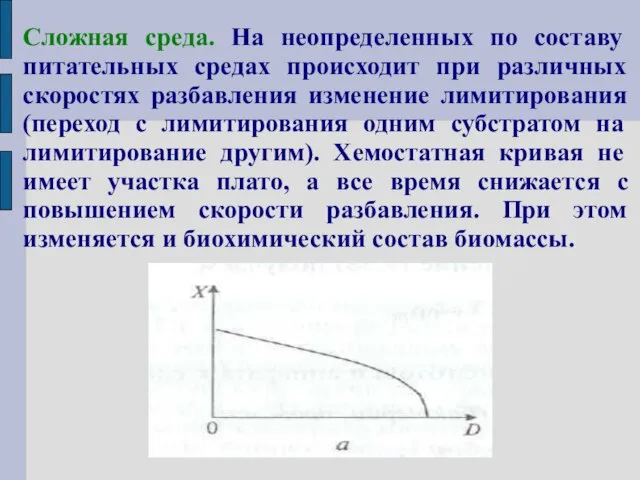
Сложная среда. На неопределенных по составу питательных средах происходит при различных
Сложная среда. На неопределенных по составу питательных средах происходит при различных
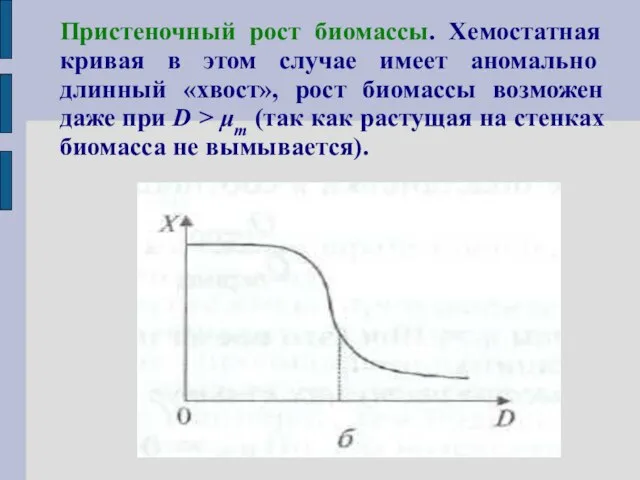
Пристеночный рост биомассы. Хемостатная кривая в этом случае имеет аномально длинный
Пристеночный рост биомассы. Хемостатная кривая в этом случае имеет аномально длинный

ХЕМОСТАТНЫЙ ПРОЦЕСС КУЛЬТИВИРОВАНИЯ, ЛИМИТИРОВАННЫЙ КИСЛОРОДОМ
Кинетика роста аэробных культур микроорганизмов
ХЕМОСТАТНЫЙ ПРОЦЕСС КУЛЬТИВИРОВАНИЯ, ЛИМИТИРОВАННЫЙ КИСЛОРОДОМ
Кинетика роста аэробных культур микроорганизмов

Если ферментационное оборудование недостаточно интенсивное, эта зависимость вырождается в однофакторную зависимость
от
Если ферментационное оборудование недостаточно интенсивное, эта зависимость вырождается в однофакторную зависимость от

Из условия стационарности dX/dt = О можно найти значение установившейся
Из условия стационарности dX/dt = О можно найти значение установившейся

Последним членом уравнения в связи с низкой растворимостью кислорода в
Последним членом уравнения в связи с низкой растворимостью кислорода в

отсюда
концентрация негазообразного субстрата S, хотя и не лимитирует процесс, также зависит
отсюда
концентрация негазообразного субстрата S, хотя и не лимитирует процесс, также зависит
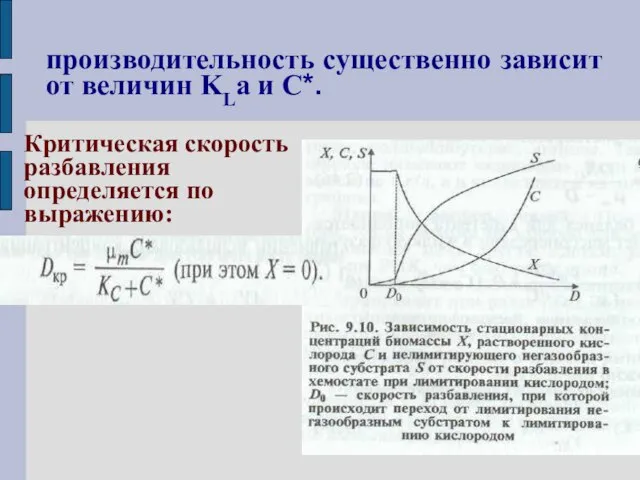
производительность существенно зависит от величин KLa и С*.
Критическая скорость
разбавления
определяется
производительность существенно зависит от величин KLa и С*.
Критическая скорость
разбавления
определяется

При некотором достаточно малом значении скорости разбавления концентрация негазообразного субстрата снижается
При некотором достаточно малом значении скорости разбавления концентрация негазообразного субстрата снижается

в этом случае оптимальной скоростью разбавления является точка переключения лимитирования
в этом случае оптимальной скоростью разбавления является точка переключения лимитирования

АВТОСЕЛЕКЦИЯ В НЕПРЕРЫВНОМ ПРОЦЕССЕ
В ходе процесса культивирования под воздействием космических
АВТОСЕЛЕКЦИЯ В НЕПРЕРЫВНОМ ПРОЦЕССЕ
В ходе процесса культивирования под воздействием космических
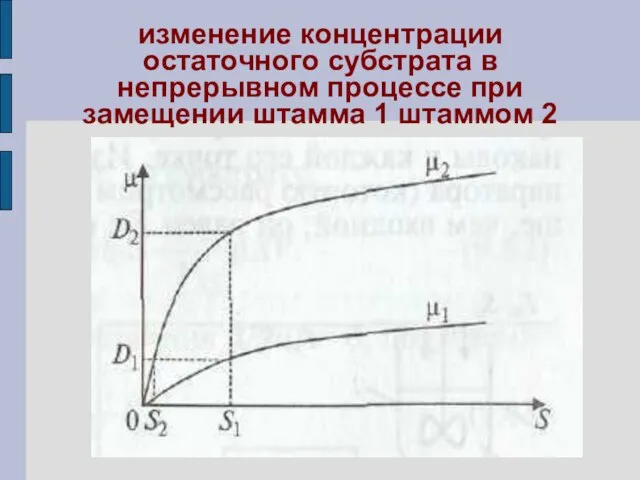
изменение концентрации остаточного субстрата в непрерывном процессе при замещении штамма 1
изменение концентрации остаточного субстрата в непрерывном процессе при замещении штамма 1

Если процесс протекает при скорости разбавления D1 то исходный штамм
Если процесс протекает при скорости разбавления D1 то исходный штамм

Рост происходит до тех пор, пока концентрация субстрата не снизится до
Рост происходит до тех пор, пока концентрация субстрата не снизится до

внешнее проявление автоселекции в непрерывном процессе
внешнее проявление автоселекции в непрерывном процессе
 Скелет человека Осевой скелет и скелет конечностей. Автор: Орлова Наталья Алексеевна Учитель биологии и химии М
Скелет человека Осевой скелет и скелет конечностей. Автор: Орлова Наталья Алексеевна Учитель биологии и химии М Круги кровообращения
Круги кровообращения Деление клетки. Митоз. (9 класс)
Деление клетки. Митоз. (9 класс) Плоды - яблоко, костянка, сборная семянка, сборная костянка. Плоды - яблоко, костянка, сборная семянка, сборная костянка. В это семей
Плоды - яблоко, костянка, сборная семянка, сборная костянка. Плоды - яблоко, костянка, сборная семянка, сборная костянка. В это семей Здоровье человека
Здоровье человека Семейство злаковые
Семейство злаковые ВПР по биологии в 5 классе 2018 год. №7 Работа с текстом
ВПР по биологии в 5 классе 2018 год. №7 Работа с текстом Учебный проект по окружающему миру «Разнообразие животных в природе» 4 класс VIII вида
Учебный проект по окружающему миру «Разнообразие животных в природе» 4 класс VIII вида Тип Хордовые: класс Рыбы, класс Земноводные или Амфибии, класс Пресмыкающиеся или Рептилии
Тип Хордовые: класс Рыбы, класс Земноводные или Амфибии, класс Пресмыкающиеся или Рептилии Движение крови в организме
Движение крови в организме Отряды птиц: Дневные хищные. Совы. Куриные. Презентация по биологии для учащихся 7 класса Автор Невмержицкая Е.Н. учитель биологии
Отряды птиц: Дневные хищные. Совы. Куриные. Презентация по биологии для учащихся 7 класса Автор Невмержицкая Е.Н. учитель биологии Физиологические свойства сердечной мышцы. Нагнетательная функция сердца. Фазовый анализ сердечного цикла
Физиологические свойства сердечной мышцы. Нагнетательная функция сердца. Фазовый анализ сердечного цикла Презентация на тему "Вода – растворитель" - скачать презентации по Биологии
Презентация на тему "Вода – растворитель" - скачать презентации по Биологии Теории возникновения жизни Теории возникновения жизни на Земле.
Теории возникновения жизни Теории возникновения жизни на Земле. Каждый день – всегда, везде, На занятиях, в игре Смело, чётко говорим И тихонечко сидим.
Каждый день – всегда, везде, На занятиях, в игре Смело, чётко говорим И тихонечко сидим.  Вода-жизнь Выполнили ученики 9 класса «А» Романенко Виталий, Жигалова Евгения, Павленко Ольга Научный руководитель: учитель био
Вода-жизнь Выполнили ученики 9 класса «А» Романенко Виталий, Жигалова Евгения, Павленко Ольга Научный руководитель: учитель био Самые необычные насекомые В природе огромное множество насекомых, многие из которых имеют очень интересные и необычные формы.
Самые необычные насекомые В природе огромное множество насекомых, многие из которых имеют очень интересные и необычные формы.  Семейство аистовые
Семейство аистовые Подготовила ученица 9 класса «гимназии № 8» Гафурова Динара.
Подготовила ученица 9 класса «гимназии № 8» Гафурова Динара. Химический состав клетки. Опорные таблицы по курсу 10 класса
Химический состав клетки. Опорные таблицы по курсу 10 класса Каракал
Каракал Холодные полярные пустыни
Холодные полярные пустыни Основы генетики
Основы генетики Общие вопросы анатомии и физиологии костной и мышечной системы
Общие вопросы анатомии и физиологии костной и мышечной системы Презентация на тему Биогеоценоз как особый уровень организации жизни Урок биологии 10 класс
Презентация на тему Биогеоценоз как особый уровень организации жизни Урок биологии 10 класс Особенности высшей нервной деятельности человека. Речь. Мышление.труд. Познавательные процессы
Особенности высшей нервной деятельности человека. Речь. Мышление.труд. Познавательные процессы Міні-проект «Живі фільтрі»
Міні-проект «Живі фільтрі» Тема: Молекулярная биология гена
Тема: Молекулярная биология гена