- Нервная ткань. Молекулярные механизмы синаптической передачи

Содержание
- 2. Передача сигнала в нейронах идет в два этапа. На первом этапе передается электрический импульс вдоль отростка
- 4. В процессе передачи нервного импульса в любом синапсе можно выделить следующие этапы: -потенциал действия, дошедший до
- 6. Ацетилхолиновый синапс. Нервно-мышечный синапс относится к группе так называемых холинэргических синапсов, поскольку в качестве медиатора используется
- 7. Ацетилхолиновый синапс. постганглионарных нейронов парасимпатической нервной системы (сердца, гладких мышц, желез) называют м-холинорецепторы (мускарин чувствительные), Ацетилхолиновые
- 8. Воздействие ацетилхолина через н-холинорецептры приводит к передаче нервного импульса с преганглионарного аксона на постганглионарные нейроны в
- 9. Адренергический синапс Медиаторами в адренергическом синапсе служат норадреналин Норадреналин содержатся в синапсах разных отделах головного мозга
- 10. Постсинаптические мембраны имеют два основных класса адренорецепторов: α - и β- типы. α1 - рецепторы возбуждения
- 11. - Адренорецептроное взаимодействие в адренергическом синапсе не приводит к открытию лиганд - зависимых ионных каналов. -
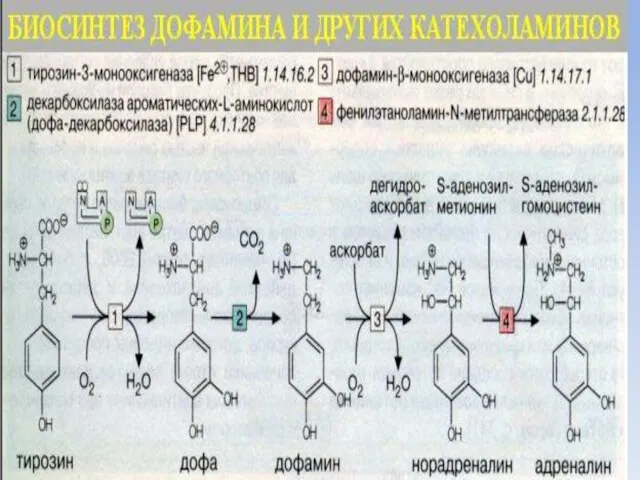
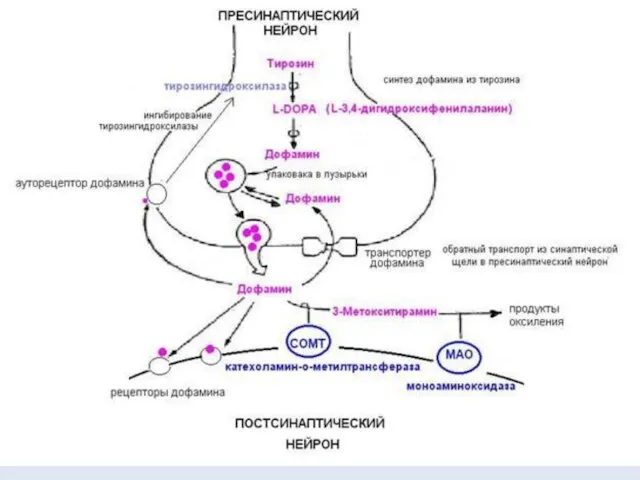
- 12. Дофаминовый синапс Разновидностью адренергического синапса является дофаминовый синапс, медиаторную функцию, в котором выполняет дофамин. Дофамин –
- 13. Поступивший в синаптическую щель дофамин связывается с рецепторами на постсинаптической мембране и с ауторецепторами пресинаптического нейрона.
- 14. Дофаминовые синапсы с D1- рецепторами на постсинаптической мембране образованы нейронами черной субстанции ствола мозга, аксоны которых
- 15. Серотониновый синапс Серотонин (5-окситриптамин) является медиатором нейронов, обеспечивающих межнейронные связи срединных ядер ствола мозга, участвующие в
- 16. Серотонин синтезируется в цитоплазме нервных окончаний из триптофана и накапливается в синаптических пузырьках. Различают три типа
- 17. Блокада S3-рецепторов антисеротониновыми препаратами может снять тошноту и рвоту, Блокада S2-рецепторов – мигренозные боли. Блокада серотониновых
- 18. Серотонин (5-окситриптамин) является медиатором сенсорных и облегчающих нейронов. Количество серотонина выделяемого в сенсорных нейронах изменяется в
- 19. Тормозные синапсы Медиаторами синапсов передающих тормозящий сигнал являются аминокислота глицин и γ-аминомасляная кислота (ГАМК). γ-аминомасляная кислота
- 20. γ-аминомасляная кислота образуется из глутаминовой кислоты под воздействием глутаматдекарбоксилазы. Коферментом этого фермента является фосфопиридоксаль – производное
- 21. Глицин – медиатор межнейронных тормозных синапсов продолговатого и спинного мозга. Специфическим антагонистом рецепторов глицина является стрихнин.
- 22. Нейро-медиаторы пептидной природы Вещество Р – полипептид был выделен в 1931 из стволовой части мозга, состоит
- 23. Ангиотензин II – октапептид участвует в регуляции водно-солевого обмена. Как медиатор функционирует в области третьего желудочка
- 24. Эндорфины Эндорфины оказывают выраженные морфиноподобные эффекты в мозге (вызывают обезболивание, снимают чувство страха и др). Эндорфины
- 25. Эндорфины и энкефалины синтезируются в гипоталамусе и других отделах мозга - в основном там, где есть
- 26. Эндогенные опиаты – пептиды по-разному могут влиять на сексуальное поведение, так β-эндорфин и метионин-энкефалин оказывают тормозящий
- 27. Механизмы памяти Кратковременная память представлена ревербирующими контурами, возникающими в коре больших полушарий и быстро исчезающими. По
- 28. Допустим, улитке подали электрический ток на хвост. Ей нужно хвост отдернуть. Каким образом это происходит. От
- 29. Если раздражение продолжает поступать регулярно, то эта реакция – долгосрочная память (работает 12-24 часа). В этом
- 31. Долговременная память сохраняется в течение длительного времени и связана с появлением устойчивых изменений в химизме нейронов
- 32. Полиамины спермидин и спермин содержатся во всех клетках и локализованы в ядре. Они входят с состав
- 33. Таким образом, прохождение импульса через синапс приводит к удлинению свойств этого синапса, т.е. имеет место пролонгация
- 34. Есть концепция, о конкурентном характере формирования связей в разных участках коры, принимающих сигналы от многочисленных рецепторов.
- 36. Таким образом, в результате приобретения жизненного опыта (памяти) изначально одинаковые нейроны делятся на группы, которые взаимодействуют
- 38. Скачать презентацию

Передача сигнала в нейронах идет в два этапа.
На первом этапе
Передача сигнала в нейронах идет в два этапа.
На первом этапе

В процессе передачи нервного импульса в любом синапсе можно выделить следующие
В процессе передачи нервного импульса в любом синапсе можно выделить следующие

Ацетилхолиновый синапс.
Нервно-мышечный синапс относится к группе так называемых холинэргических синапсов, поскольку
Ацетилхолиновый синапс.
Нервно-мышечный синапс относится к группе так называемых холинэргических синапсов, поскольку

Ацетилхолиновый синапс.
постганглионарных нейронов парасимпатической нервной системы (сердца, гладких мышц, желез) называют
Ацетилхолиновый синапс.
постганглионарных нейронов парасимпатической нервной системы (сердца, гладких мышц, желез) называют

Воздействие ацетилхолина через н-холинорецептры приводит к передаче нервного импульса с преганглионарного
Воздействие ацетилхолина через н-холинорецептры приводит к передаче нервного импульса с преганглионарного

Адренергический синапс
Медиаторами в адренергическом синапсе служат норадреналин
Норадреналин содержатся в
Адренергический синапс
Медиаторами в адренергическом синапсе служат норадреналин
Норадреналин содержатся в

Постсинаптические мембраны имеют два основных класса адренорецепторов: α - и β-
Постсинаптические мембраны имеют два основных класса адренорецепторов: α - и β-

- Адренорецептроное взаимодействие в адренергическом синапсе не приводит к открытию лиганд
- Адренорецептроное взаимодействие в адренергическом синапсе не приводит к открытию лиганд

Дофаминовый синапс
Разновидностью адренергического синапса является дофаминовый синапс, медиаторную функцию, в
Дофаминовый синапс
Разновидностью адренергического синапса является дофаминовый синапс, медиаторную функцию, в

Поступивший в синаптическую щель дофамин связывается с рецепторами на постсинаптической мембране
Поступивший в синаптическую щель дофамин связывается с рецепторами на постсинаптической мембране

Дофаминовые синапсы с D1- рецепторами на постсинаптической мембране образованы нейронами черной
Дофаминовые синапсы с D1- рецепторами на постсинаптической мембране образованы нейронами черной

Серотониновый синапс
Серотонин (5-окситриптамин) является медиатором
нейронов, обеспечивающих межнейронные связи срединных ядер
Серотониновый синапс
Серотонин (5-окситриптамин) является медиатором
нейронов, обеспечивающих межнейронные связи срединных ядер

Серотонин синтезируется в цитоплазме нервных окончаний из триптофана и накапливается в
Серотонин синтезируется в цитоплазме нервных окончаний из триптофана и накапливается в

Блокада S3-рецепторов антисеротониновыми препаратами может снять тошноту и рвоту,
Блокада S2-рецепторов –
Блокада S3-рецепторов антисеротониновыми препаратами может снять тошноту и рвоту,
Блокада S2-рецепторов –

Серотонин (5-окситриптамин) является медиатором
сенсорных и облегчающих нейронов.
Количество серотонина выделяемого
Серотонин (5-окситриптамин) является медиатором
сенсорных и облегчающих нейронов.
Количество серотонина выделяемого

Тормозные синапсы
Медиаторами синапсов передающих тормозящий сигнал являются аминокислота глицин и
Тормозные синапсы
Медиаторами синапсов передающих тормозящий сигнал являются аминокислота глицин и

γ-аминомасляная кислота образуется из глутаминовой кислоты под воздействием глутаматдекарбоксилазы. Коферментом этого
γ-аминомасляная кислота образуется из глутаминовой кислоты под воздействием глутаматдекарбоксилазы. Коферментом этого

Глицин – медиатор межнейронных тормозных синапсов продолговатого и спинного мозга.
Специфическим
Глицин – медиатор межнейронных тормозных синапсов продолговатого и спинного мозга.
Специфическим

Нейро-медиаторы пептидной природы
Вещество Р – полипептид был выделен в 1931
Нейро-медиаторы пептидной природы
Вещество Р – полипептид был выделен в 1931

Ангиотензин II – октапептид участвует в регуляции водно-солевого обмена. Как медиатор
Ангиотензин II – октапептид участвует в регуляции водно-солевого обмена. Как медиатор

Эндорфины
Эндорфины оказывают выраженные морфиноподобные эффекты в мозге (вызывают обезболивание, снимают чувство
Эндорфины
Эндорфины оказывают выраженные морфиноподобные эффекты в мозге (вызывают обезболивание, снимают чувство

Эндорфины и энкефалины синтезируются в гипоталамусе и других отделах мозга -
Эндорфины и энкефалины синтезируются в гипоталамусе и других отделах мозга -

Эндогенные опиаты – пептиды по-разному могут влиять на сексуальное поведение,
так
Эндогенные опиаты – пептиды по-разному могут влиять на сексуальное поведение,
так

Механизмы памяти
Кратковременная память представлена ревербирующими контурами, возникающими в коре больших полушарий
Механизмы памяти
Кратковременная память представлена ревербирующими контурами, возникающими в коре больших полушарий

Допустим, улитке подали электрический ток на хвост. Ей нужно хвост отдернуть.
Допустим, улитке подали электрический ток на хвост. Ей нужно хвост отдернуть.

Если раздражение продолжает поступать регулярно, то эта реакция – долгосрочная память
Если раздражение продолжает поступать регулярно, то эта реакция – долгосрочная память
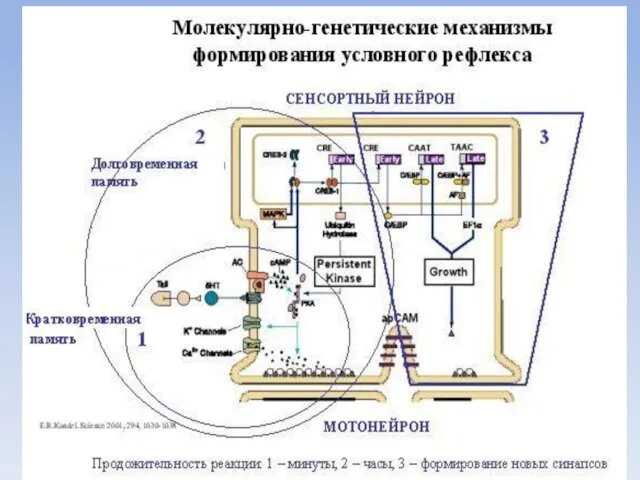

Долговременная память
сохраняется в течение длительного времени и связана с появлением устойчивых
Долговременная память
сохраняется в течение длительного времени и связана с появлением устойчивых

Полиамины спермидин и спермин содержатся во всех клетках и локализованы в
Полиамины спермидин и спермин содержатся во всех клетках и локализованы в

Таким образом, прохождение импульса через синапс приводит к удлинению свойств этого
Таким образом, прохождение импульса через синапс приводит к удлинению свойств этого

Есть концепция, о конкурентном характере формирования связей в разных участках коры,
Есть концепция, о конкурентном характере формирования связей в разных участках коры,
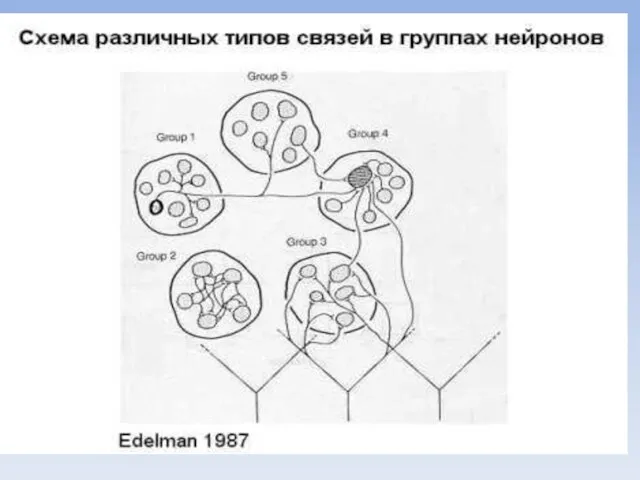

Таким образом, в результате приобретения жизненного опыта (памяти) изначально одинаковые нейроны
Таким образом, в результате приобретения жизненного опыта (памяти) изначально одинаковые нейроны
 Активные методы обучения на уроках биологии и природоведения. Русская гимназия г. Валга Авраменко О.В. 2002 г.
Активные методы обучения на уроках биологии и природоведения. Русская гимназия г. Валга Авраменко О.В. 2002 г.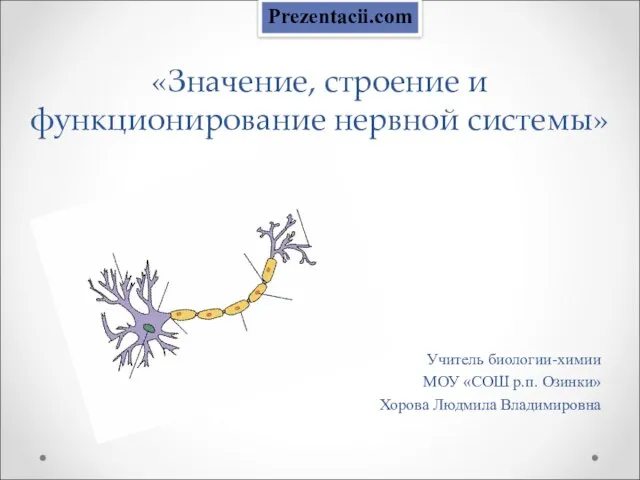 «Значение, строение и функционирование нервной системы» Учитель биологии-химии МОУ «СОШ р.п. Озинки» Хорова Людмила Владимировн
«Значение, строение и функционирование нервной системы» Учитель биологии-химии МОУ «СОШ р.п. Озинки» Хорова Людмила Владимировн Многообразие птиц
Многообразие птиц Приматы Биология 7 класс
Приматы Биология 7 класс  Презентация на тему "Взаимодействие разных организмов" - скачать презентации по Биологии
Презентация на тему "Взаимодействие разных организмов" - скачать презентации по Биологии Урок биологии. 9 класс. Генетические опыты Менделя
Урок биологии. 9 класс. Генетические опыты Менделя Клеточная биология: просто о сложном
Клеточная биология: просто о сложном Генетика - анализирующее скрещивание
Генетика - анализирующее скрещивание Презентация на тему Отходы большого города: как их собирают, удаляют и перерабатывают
Презентация на тему Отходы большого города: как их собирают, удаляют и перерабатывают  С-реактивный белок Строение, экспрессия, биологическая роль, связь с воспалением и сердечно-сосудистыми заболеваниями.
С-реактивный белок Строение, экспрессия, биологическая роль, связь с воспалением и сердечно-сосудистыми заболеваниями.  Презентация на тему Экология и здоровье человека
Презентация на тему Экология и здоровье человека  Динозавры Быков Дмитрий, 5 «Б» класс, школа №1
Динозавры Быков Дмитрий, 5 «Б» класс, школа №1 Кровь: состав и выполняемые функции
Кровь: состав и выполняемые функции Слоны. Викторина
Слоны. Викторина Мое домашнее животное
Мое домашнее животное Обмен веществ и превращение энергии
Обмен веществ и превращение энергии Размножение организмов. Оплодотворение. Урок биологии в 7 классе. Учитель: Бобылева Н.П.
Размножение организмов. Оплодотворение. Урок биологии в 7 классе. Учитель: Бобылева Н.П.  Презентация на тему Лоси
Презентация на тему Лоси Урок 1. Наука биология Разработал: Лебедев С.Н., учитель биологии высшей категории
Урок 1. Наука биология Разработал: Лебедев С.Н., учитель биологии высшей категории Уровни регуляции движений
Уровни регуляции движений Семейство Рутовые
Семейство Рутовые Пути заражения паразитарными заболеваниями
Пути заражения паразитарными заболеваниями Презентация на тему "Нематоды" - скачать бесплатно презентации по Биологии
Презентация на тему "Нематоды" - скачать бесплатно презентации по Биологии Витамины
Витамины  Эволюционное учение Чарльза Дарвина
Эволюционное учение Чарльза Дарвина Презентация на тему "Химический состав клетки" - скачать презентации по Биологии
Презентация на тему "Химический состав клетки" - скачать презентации по Биологии 10 самых удивительных растений
10 самых удивительных растений Класс брюхоногие
Класс брюхоногие