- Нуклеозиды, нуклеотиды, нуклеиновые кислоты: функции и обмен

Содержание
- 2. Нуклеотиды Нуклеотиды – это низкомолекулярные вещества, которые выполняют функции биорегуляторов (НАД, НАДФ, ФМН, ФАД, АТФ, АДФ,
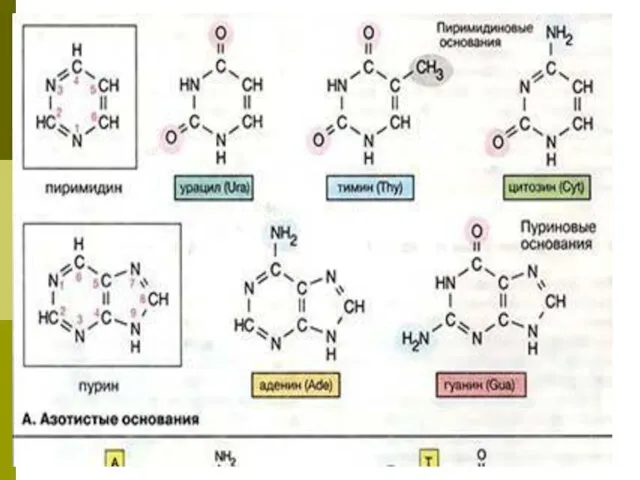
- 3. Азотистые основания При гидролизе нуклеиновых кислот образуются пуриновые (аденин, гуанин) и пиримидиновые (тимин, цитозин, урацил) основания,
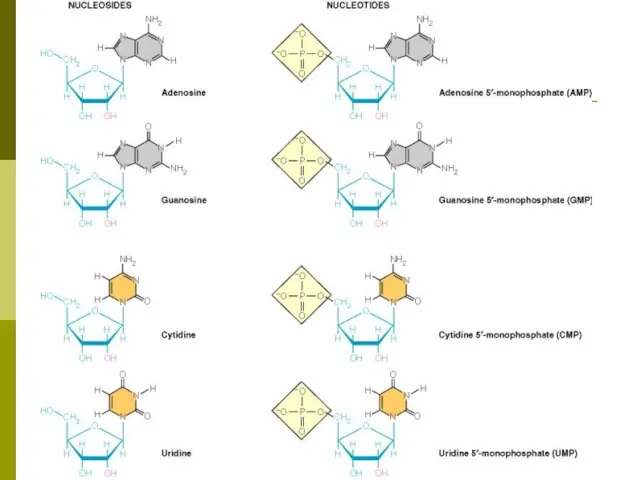
- 5. Нуклеозиды, нуклеотиды Нуклеозид - азотистое основание + углеводный компонент Нуклеотид - азотистое основание + углеводный компонент
- 6. Нуклеозид Нуклеотид
- 8. Пуриновые азотистые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике, в дальнейшем практически не используются.
- 9. Биосинтез пуринов На основе 5-фосфорибозил-1-пирофосфата строится имидазольное кольцо, затем – пуриновое. Общий предшественник пуриновых нуклеотидов –
- 10. Происхождение атомов пуринового кольца
- 11. Синтез АМФ и ГМФ Синтез АМФ и ГМФ осуществляется из ИМФ. Для синтеза ГМФ необходимы НАД+,
- 13. Регуляция синтеза пуриновых нуклеотидов Синтез пуриновых нуклеотидов тормозится конечными продуктами АМФ и ГМФ по принципу обратной
- 14. Особенностью синтеза пиримидиновых нуклеотидов является то, что вначале происходит синтез пиримидинового основания, а затем к нему
- 15. Биосинтез пиримидинов Биосинтез пиримидинов начинается с построения гетероцикла с участием NH3,,СО2,глу, асп. Общий предшественник пиримидинов оротовая
- 16. Происхождение атомов азота пиримидинового кольца
- 17. Катаболизм пуринов АМФ ?аденозин ?инозин ? гипоксантин ? ксантин ? мочевая кислота ГМФ ? гуанозин ?
- 18. Катаболизм пиримидинов ЦМФ ? УМФ ? урацил ТМФ ? тимин Восстановление и гидролиз пиримидинов ? раскрытие
- 19. Синтез дезоксинуклеотидов Все нуклеотиды образуются с участием фосфорибозилпирофосфата. Дезоксирибонуклеотиды образуются при восстановлении рибозы до дезоксирибозы в
- 20. Нуклеиновые кислоты
- 21. НУКЛЕОПРОТЕИНЫ Нуклеопротеины – сложные белки, которые состоят из белковой части и небелковой части – простетической группы,
- 22. ФУНКЦИИ ДНК (ПО А. ЛЕНИНДЖЕPУ) хpанение запаса генетической инфоpмации, необходимой для кодиpования стpуктуpы всех белков и
- 23. ВИДЫ ДНК ядерные (хромосомные) ДНК; ДНК плазмид; ДНК хлоропластов; ДНК митохондрий; ДНК вирусов.
- 24. Виды РНК матричные (информационные) транспортные рибосомальные
- 25. Пространственная структура нуклеиновых кислот Первичная структура – последовательность нуклеотидов Вторичная структура – двойная спираль ДНК (А,В,С,Д
- 26. Правила Чаргаффа Количество пуриновых оснований равно количеству пиримидиновых оснований: А + Г = Ц + Т
- 27. ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ АЗОТИСТЫМИ ОСНОВАНИЯМИ
- 28. ВТОРИЧНАЯ СТРУКТУРА ДНК
- 29. Матричные синтезы
- 30. Виды передачи генетической информации ДНК → ДНК – репликация ДНК → РНК – транскрипция РНК →
- 32. Репликация ДНК Вариант матричного синтеза, представляет собой удвоение цепей ДНК. Матрицей служит каждая из одноцепочечных последовательностей
- 34. Полуконсервативная репликация
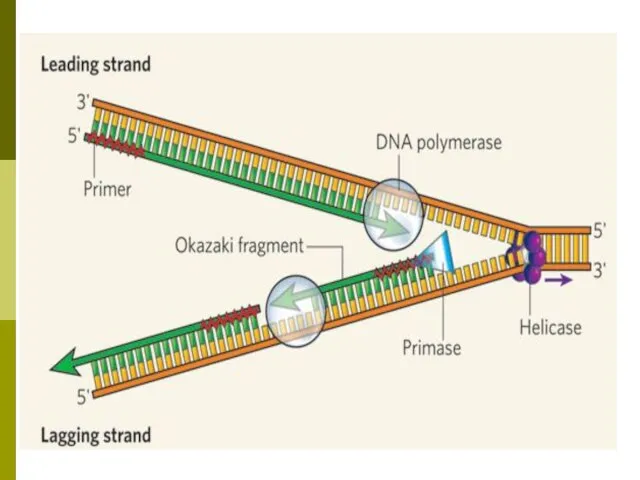
- 35. Репликация ДНК Три этапа: инициация, элонгация, терминация (созревание). Репарация ошибок и повреждений. В репликации участвуют: 1)
- 36. Характеристики репликации матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с
- 38. Ответ ДНК на повреждение Активация контрольной точки клеточного цикла Активация транскрипционной программы Репарация ДНК Клеточный метаболизм
- 39. Репарация ДНК Измененный участок ДНК распознается и удаляется при помощи ферментов ДНК-репарирующих эндонуклеаз. ДНК-полимераза I связывается
- 40. Транскрипция Транскрипция - биосинтез РНК на матрице ДНК Последовательность рибонуклеотидов в молекуле РНК комплементарна последовательности дезоксирибонуклеотидов
- 41. Ферменты У эукариот три РНК-полимеразы: I – синтезирует тРНК, II – мРНК, III – рРНК. У
- 42. Транскрипция 3 стадии транскрипции: инициация, элонгация, терминация ИНИЦИАЦИЯ синтеза начинается с «узнавания» полимеразой промоторного сайта (не
- 43. Оперон и транскриптон Единицей транскрипции у прокариот является оперон Единицей транскрипции у эукариот транскриптон
- 44. Структура оперона Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза; в ДНК E. coli
- 46. Посттрансляционная модификация РНК В результате транскрипции образуются три типа предшественников РНК (первичные транскрипты): предшественник мРНК, или
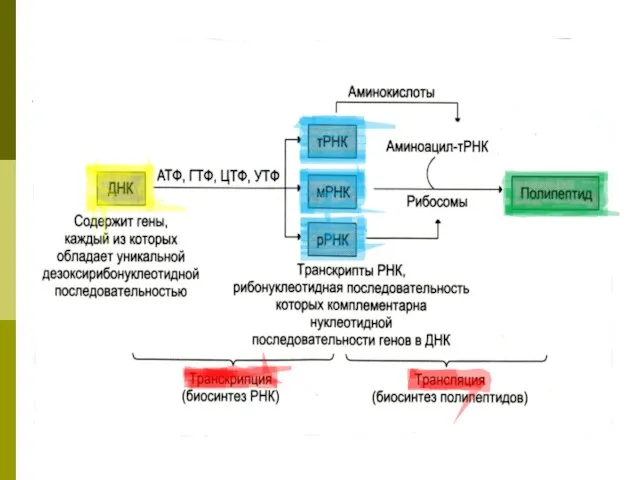
- 47. Трансляция Перевод генетической информации с кодонов м РНК на аминокислотную последовательность белка (экспрессия гена). Генетический код:
- 48. Для перевода нуклеотидного кода в аминокислотную последовательность служат молекулы-адаптеры аминоацил-тРНК: на 3’-конце – аминокислота, а в
- 49. Трансляция для синтеза белка необходимы: 20 аминокислот мРНК Рибосома 50 тРНК (одна тРНК может связывать несколько
- 50. РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на 5’-конце мРНК в функционирующую
- 51. ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА ДНК: информация о последовательности аминокислот в полипептидной цепи записана в
- 52. Генетический код
- 53. Белоксинтезирующая система клетки мРНК – матрица, на которой записана последовательность аминокислот белка в виде последовательности триплетов.
- 57. Фолдинг и процессинг От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин. Сигнальная последовательность на N-конце
- 59. Скачать презентацию

Нуклеотиды
Нуклеотиды – это низкомолекулярные вещества, которые выполняют функции биорегуляторов (НАД, НАДФ,
Нуклеотиды
Нуклеотиды – это низкомолекулярные вещества, которые выполняют функции биорегуляторов (НАД, НАДФ,

Азотистые основания
При гидролизе нуклеиновых кислот образуются пуриновые (аденин, гуанин) и пиримидиновые
Азотистые основания
При гидролизе нуклеиновых кислот образуются пуриновые (аденин, гуанин) и пиримидиновые

Нуклеозиды, нуклеотиды
Нуклеозид - азотистое основание + углеводный компонент
Нуклеотид - азотистое
Нуклеозиды, нуклеотиды
Нуклеозид - азотистое основание + углеводный компонент
Нуклеотид - азотистое
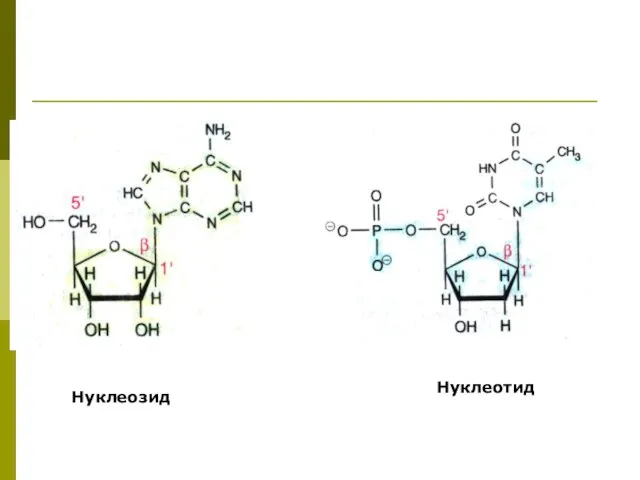
Нуклеозид
Нуклеотид
Нуклеозид
Нуклеотид

Пуриновые азотистые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике,
Пуриновые азотистые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике,

Биосинтез пуринов
На основе 5-фосфорибозил-1-пирофосфата строится имидазольное кольцо,
затем – пуриновое.
Общий предшественник
Биосинтез пуринов
На основе 5-фосфорибозил-1-пирофосфата строится имидазольное кольцо,
затем – пуриновое.
Общий предшественник
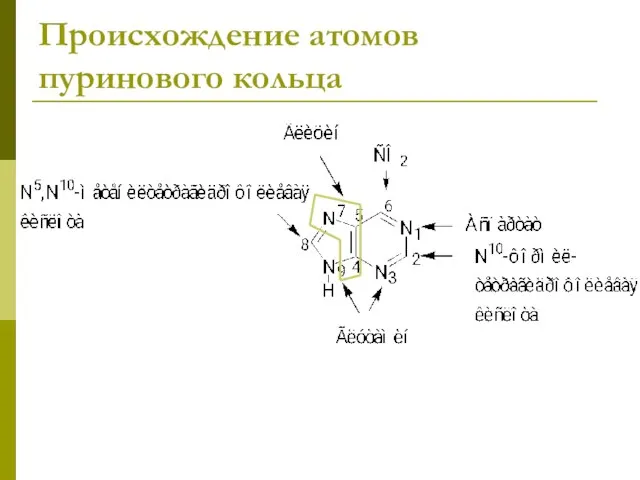
Происхождение атомов пуринового кольца
Происхождение атомов пуринового кольца

Синтез АМФ и ГМФ
Синтез АМФ и ГМФ осуществляется из ИМФ.
Для
Синтез АМФ и ГМФ
Синтез АМФ и ГМФ осуществляется из ИМФ.
Для
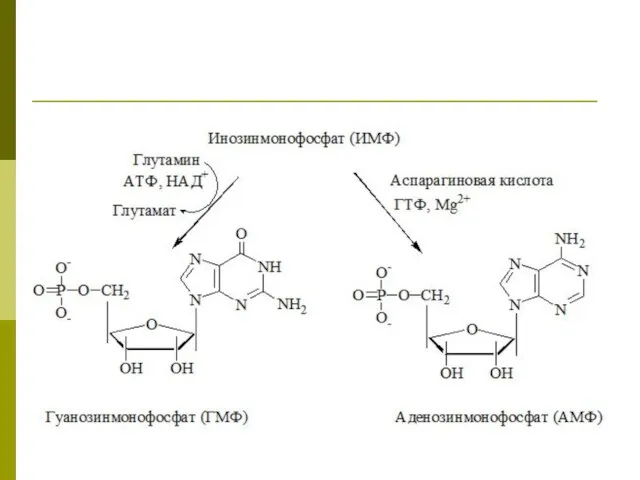

Регуляция синтеза пуриновых нуклеотидов
Синтез пуриновых нуклеотидов тормозится конечными продуктами АМФ и
Регуляция синтеза пуриновых нуклеотидов
Синтез пуриновых нуклеотидов тормозится конечными продуктами АМФ и

Особенностью синтеза пиримидиновых нуклеотидов является то, что вначале происходит синтез пиримидинового
Особенностью синтеза пиримидиновых нуклеотидов является то, что вначале происходит синтез пиримидинового

Биосинтез пиримидинов
Биосинтез пиримидинов начинается с построения гетероцикла с участием NH3,,СО2,глу, асп.
Общий
Биосинтез пиримидинов
Биосинтез пиримидинов начинается с построения гетероцикла с участием NH3,,СО2,глу, асп.
Общий
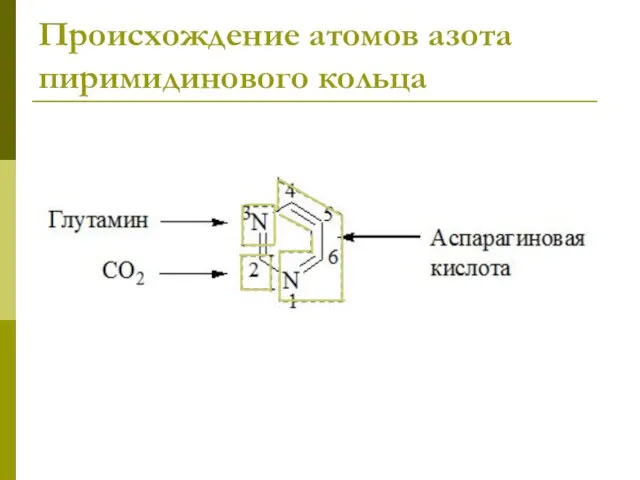
Происхождение атомов азота пиримидинового кольца
Происхождение атомов азота пиримидинового кольца

Катаболизм пуринов
АМФ ?аденозин ?инозин ? гипоксантин ?
ксантин ? мочевая кислота
ГМФ
Катаболизм пуринов
АМФ ?аденозин ?инозин ? гипоксантин ?
ксантин ? мочевая кислота
ГМФ

Катаболизм пиримидинов
ЦМФ ? УМФ ? урацил
ТМФ ? тимин
Восстановление и гидролиз пиримидинов
Катаболизм пиримидинов
ЦМФ ? УМФ ? урацил
ТМФ ? тимин
Восстановление и гидролиз пиримидинов

Синтез дезоксинуклеотидов
Все нуклеотиды образуются с участием фосфорибозилпирофосфата.
Дезоксирибонуклеотиды образуются при восстановлении рибозы
Синтез дезоксинуклеотидов
Все нуклеотиды образуются с участием фосфорибозилпирофосфата.
Дезоксирибонуклеотиды образуются при восстановлении рибозы

Нуклеиновые кислоты
Нуклеиновые кислоты

НУКЛЕОПРОТЕИНЫ
Нуклеопротеины – сложные белки, которые состоят из белковой части и небелковой
НУКЛЕОПРОТЕИНЫ
Нуклеопротеины – сложные белки, которые состоят из белковой части и небелковой

ФУНКЦИИ ДНК (ПО А. ЛЕНИНДЖЕPУ)
хpанение запаса генетической инфоpмации, необходимой для кодиpования
ФУНКЦИИ ДНК (ПО А. ЛЕНИНДЖЕPУ)
хpанение запаса генетической инфоpмации, необходимой для кодиpования

ВИДЫ ДНК
ядерные (хромосомные) ДНК;
ДНК плазмид;
ДНК хлоропластов;
ДНК митохондрий;
ДНК
ВИДЫ ДНК
ядерные (хромосомные) ДНК;
ДНК плазмид;
ДНК хлоропластов;
ДНК митохондрий;
ДНК

Виды РНК
матричные (информационные)
транспортные
рибосомальные
Виды РНК
матричные (информационные)
транспортные
рибосомальные

Пространственная структура нуклеиновых кислот
Первичная структура – последовательность нуклеотидов
Вторичная структура –
двойная
Пространственная структура нуклеиновых кислот
Первичная структура – последовательность нуклеотидов
Вторичная структура –
двойная

Правила Чаргаффа
Количество пуриновых оснований равно количеству пиримидиновых оснований:
А + Г
Количество пуриновых оснований равно количеству пиримидиновых оснований:
А + Г
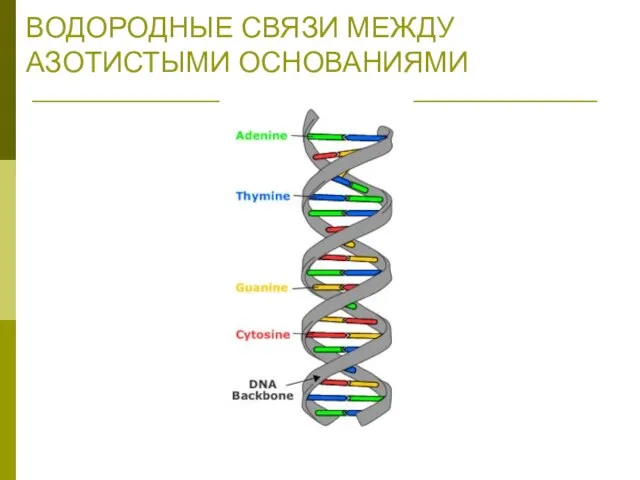
ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ АЗОТИСТЫМИ ОСНОВАНИЯМИ
ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ АЗОТИСТЫМИ ОСНОВАНИЯМИ
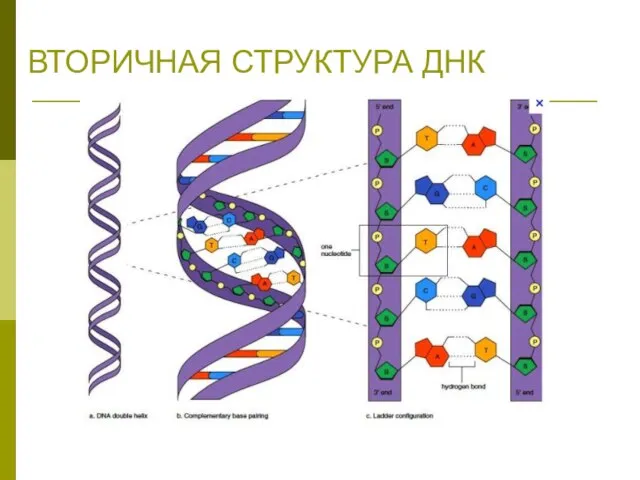
ВТОРИЧНАЯ СТРУКТУРА ДНК
ВТОРИЧНАЯ СТРУКТУРА ДНК

Матричные синтезы
Матричные синтезы

Виды передачи генетической информации
ДНК → ДНК – репликация
ДНК → РНК
Виды передачи генетической информации
ДНК → ДНК – репликация
ДНК → РНК

Репликация ДНК
Вариант матричного синтеза, представляет собой удвоение цепей ДНК.
Матрицей служит
Репликация ДНК
Вариант матричного синтеза, представляет собой удвоение цепей ДНК. Матрицей служит
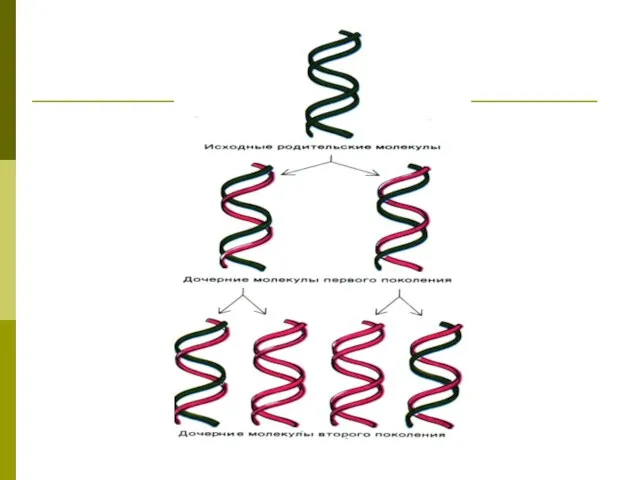
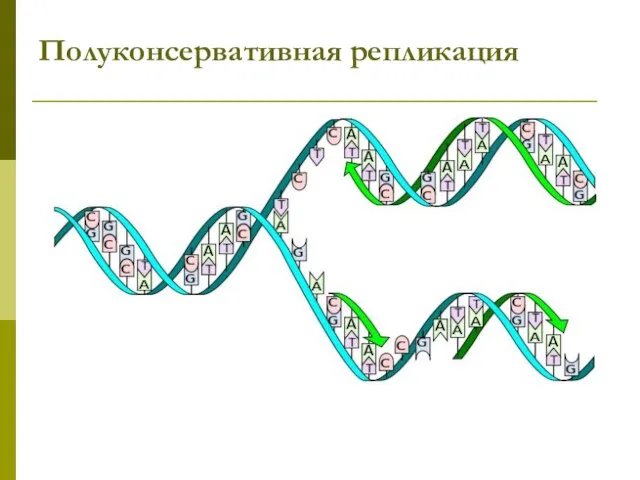
Полуконсервативная репликация
Полуконсервативная репликация

Репликация ДНК
Три этапа: инициация, элонгация,
терминация (созревание).
Репарация ошибок и повреждений.
В
Репликация ДНК
Три этапа: инициация, элонгация,
терминация (созревание).
Репарация ошибок и повреждений.
В

Характеристики репликации
матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской
Характеристики репликации
матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской
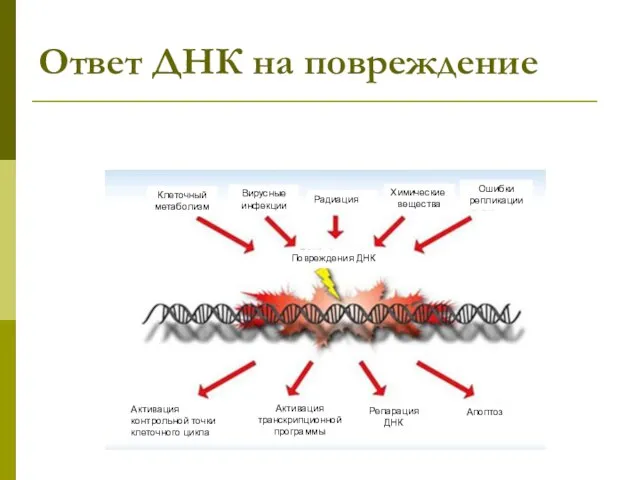
Ответ ДНК на повреждение
Активация контрольной точки клеточного цикла
Активация транскрипционной программы
Репарация ДНК
Клеточный
Ответ ДНК на повреждение
Активация контрольной точки клеточного цикла
Активация транскрипционной программы
Репарация ДНК
Клеточный

Репарация ДНК
Измененный участок ДНК распознается и удаляется при помощи ферментов ДНК-репарирующих
Репарация ДНК
Измененный участок ДНК распознается и удаляется при помощи ферментов ДНК-репарирующих

Транскрипция
Транскрипция - биосинтез РНК на матрице ДНК
Последовательность рибонуклеотидов в молекуле РНК
Транскрипция
Транскрипция - биосинтез РНК на матрице ДНК
Последовательность рибонуклеотидов в молекуле РНК

Ферменты
У эукариот три РНК-полимеразы:
I – синтезирует тРНК, II – мРНК,
Ферменты
У эукариот три РНК-полимеразы: I – синтезирует тРНК, II – мРНК,

Транскрипция
3 стадии транскрипции: инициация,
элонгация,
терминация
ИНИЦИАЦИЯ синтеза
начинается с «узнавания»
Транскрипция
3 стадии транскрипции: инициация,
элонгация,
терминация
ИНИЦИАЦИЯ синтеза
начинается с «узнавания»

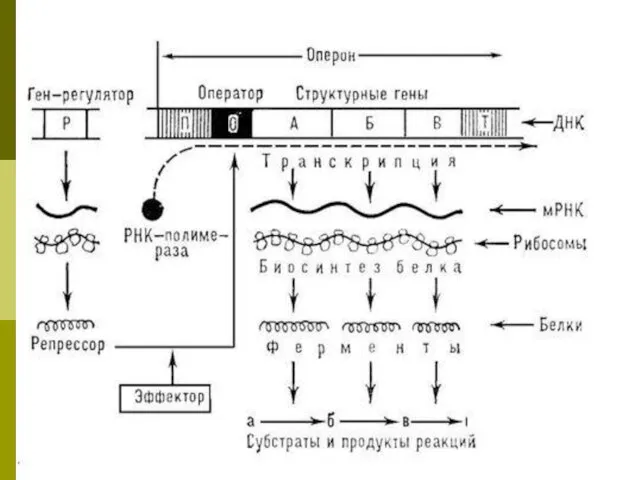
Оперон и транскриптон
Единицей транскрипции у прокариот является оперон
Единицей транскрипции у
Оперон и транскриптон
Единицей транскрипции у прокариот является оперон
Единицей транскрипции у

Структура оперона
Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза;
Структура оперона
Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза;

Посттрансляционная модификация РНК
В результате транскрипции образуются три типа предшественников РНК (первичные
Посттрансляционная модификация РНК
В результате транскрипции образуются три типа предшественников РНК (первичные

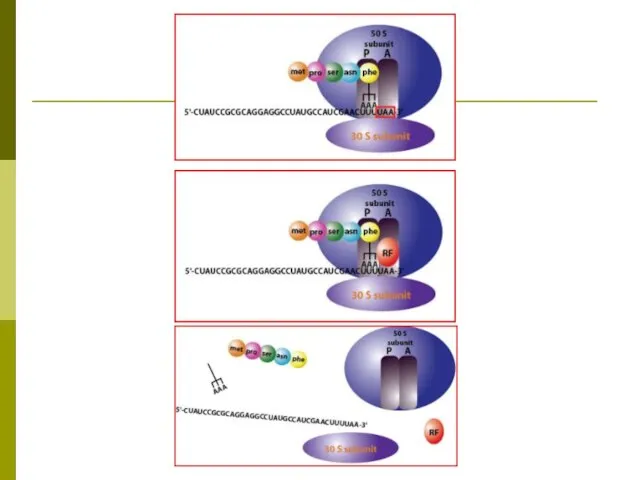
Трансляция
Перевод генетической информации с кодонов м РНК на аминокислотную последовательность белка
Трансляция
Перевод генетической информации с кодонов м РНК на аминокислотную последовательность белка

Для перевода нуклеотидного кода в аминокислотную последовательность служат молекулы-адаптеры аминоацил-тРНК: на
Для перевода нуклеотидного кода в аминокислотную последовательность служат молекулы-адаптеры аминоацил-тРНК: на

Трансляция
для синтеза белка необходимы:
20 аминокислот
мРНК
Рибосома
50 тРНК (одна тРНК может
Трансляция
для синтеза белка необходимы:
20 аминокислот
мРНК
Рибосома
50 тРНК (одна тРНК может

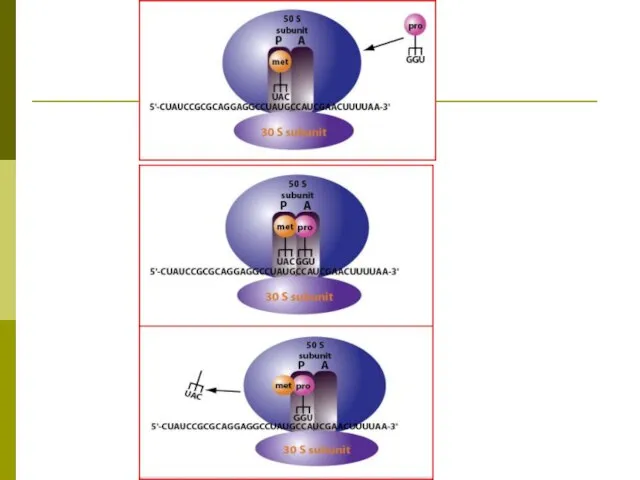
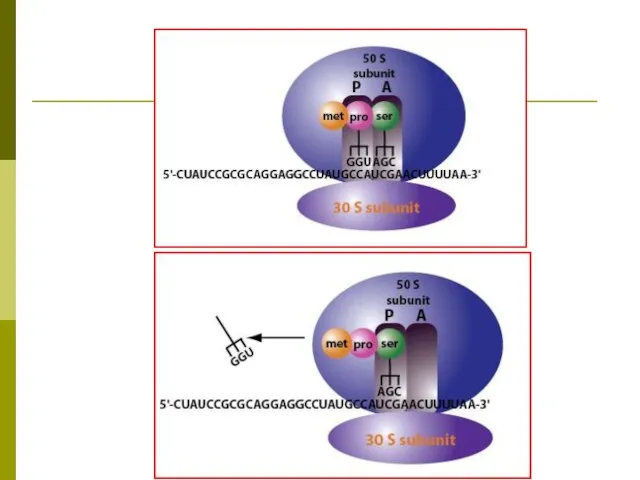
РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА
В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на
РИБОСОМНЫЙ ЦИКЛ ДЖ.УОТСОНА
В начале синтеза полипептидной цепи субъединицы рибосомы объединяются на

ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА
ДНК: информация о последовательности аминокислот в
ПЕРЕДАЧА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ СИНТЕЗЕ БЕЛКА
ДНК: информация о последовательности аминокислот в
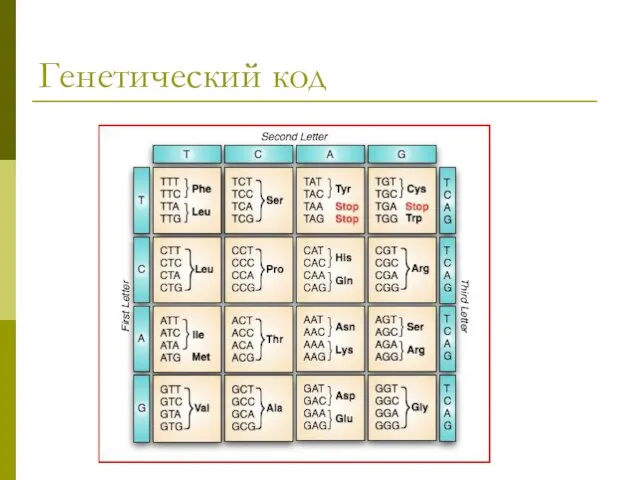
Генетический код
Генетический код

Белоксинтезирующая система клетки
мРНК – матрица, на которой записана последовательность аминокислот белка
Белоксинтезирующая система клетки
мРНК – матрица, на которой записана последовательность аминокислот белка

Фолдинг и процессинг
От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин.
Сигнальная
Фолдинг и процессинг
От синтезированного пептида в цитозоле отщепляется инициирующая аминокислота (формил)метионин.
Сигнальная
 Мышечные ткани
Мышечные ткани Терморегуляция. Изотермия, теплообразование, теплоотдача
Терморегуляция. Изотермия, теплообразование, теплоотдача Природа живая и не живая
Природа живая и не живая 20180117_zhivotnyy_i_rastitelnyy_mir_yuzhnoy_ameriki-szhat_-_kopiya
20180117_zhivotnyy_i_rastitelnyy_mir_yuzhnoy_ameriki-szhat_-_kopiya Рыба-меч
Рыба-меч Из гусеницы в бабочку Презентацию создала ученица 6 класса ДДТ Козенбаева Юлиана, Консультант педагог дополнительного образова
Из гусеницы в бабочку Презентацию создала ученица 6 класса ДДТ Козенбаева Юлиана, Консультант педагог дополнительного образова Друзья природы
Друзья природы Подкорковые ядра
Подкорковые ядра Интеллектуальный турнир Своя игра
Интеллектуальный турнир Своя игра Отдел мохообразные
Отдел мохообразные Презентация на тему МАТЬ-И-МАЧЕХА
Презентация на тему МАТЬ-И-МАЧЕХА  Дыхательная недостаточность. Лечебный и педиатрический факультеты (задачи)
Дыхательная недостаточность. Лечебный и педиатрический факультеты (задачи) Як спілкуються тварини
Як спілкуються тварини Автор: Афанасьева Татьяна Викторовна МКОУ Орешковская основная общеобразовательная школа Луховицкого района Московской области
Автор: Афанасьева Татьяна Викторовна МКОУ Орешковская основная общеобразовательная школа Луховицкого района Московской области GM-foods. Would you like to eat GM tomato?
GM-foods. Would you like to eat GM tomato? Дыхание растений
Дыхание растений Биология наука о жизни
Биология наука о жизни Птицы лесов
Птицы лесов Задания по теме Эмбриогенез 1 и 2 часть
Задания по теме Эмбриогенез 1 и 2 часть Презентация к уроку биологии на тему: «Иммунитет» 8 класс Выполнила: учитель химии и биологии НОУ СОШ «Кристалл» Жданова Надежда
Презентация к уроку биологии на тему: «Иммунитет» 8 класс Выполнила: учитель химии и биологии НОУ СОШ «Кристалл» Жданова Надежда  Хрящевые ткани
Хрящевые ткани Органы цветковых растений. (6 класс)
Органы цветковых растений. (6 класс) Класс Однодольные Monocotyledonae
Класс Однодольные Monocotyledonae  Микроорганизмдерді дақылдаудың негіздері
Микроорганизмдерді дақылдаудың негіздері Презентация на тему "Обмен веществ 8 класс" - скачать презентации по Биологии
Презентация на тему "Обмен веществ 8 класс" - скачать презентации по Биологии Презентация на тему Гипотезы возникновения жизни на Земле
Презентация на тему Гипотезы возникновения жизни на Земле Птицы наши друзья. Зимующие и перелётные птицы
Птицы наши друзья. Зимующие и перелётные птицы Размножение и оплодотворение у растений
Размножение и оплодотворение у растений